Alien Plant Invasions of the Natural Habitat in the Western Region of Saudi Arabia: Floristic Diversity and Vegetation Structure


Abstract
:1. Introduction
2. Materials and Methods
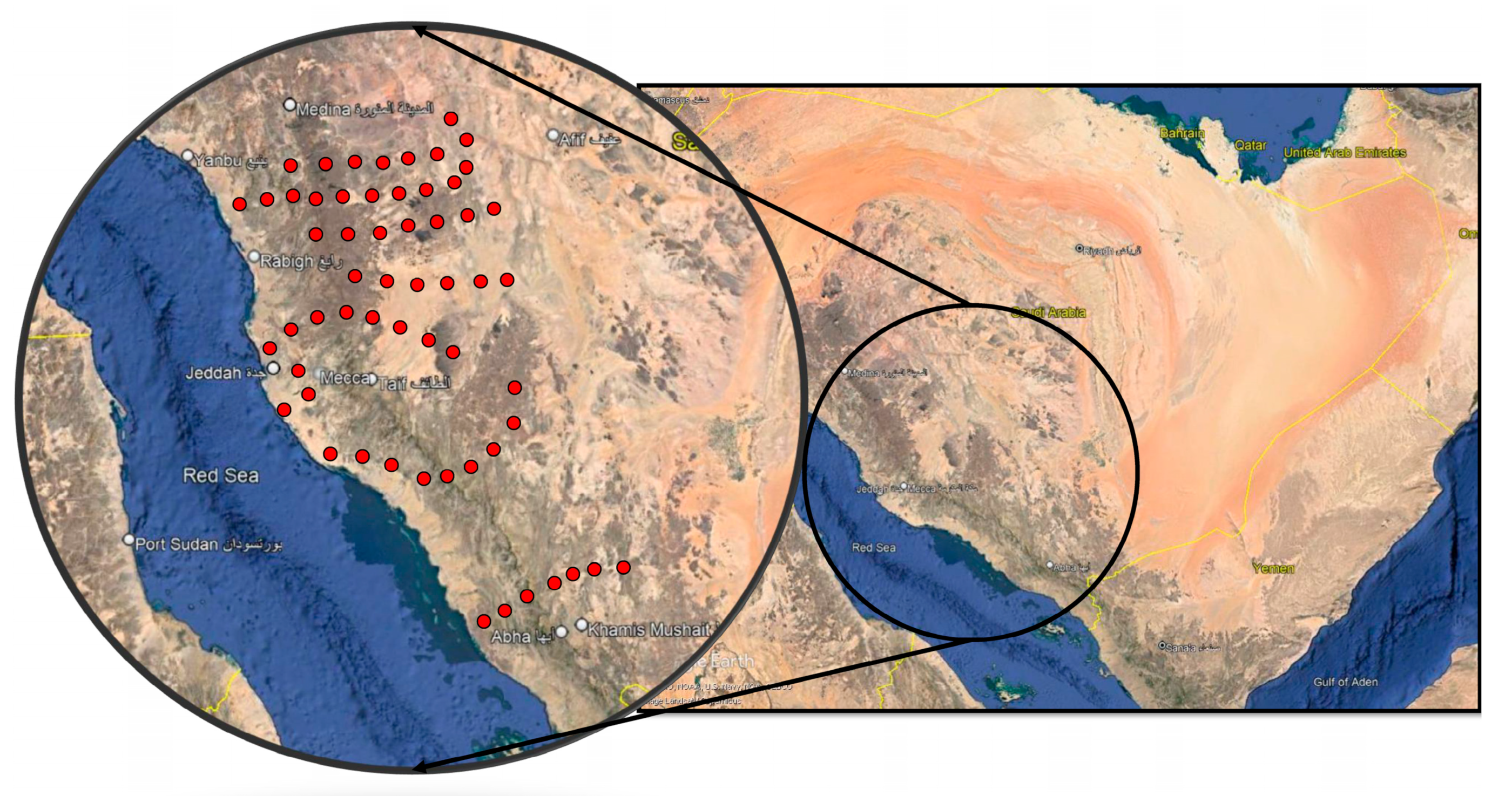
2.1. Study Area
2.2. Climatic Characteristics
2.3. Sampling Stands
2.4. Vegetation Cover and Density Estimation
2.5. Diversity Indices
2.6. Soil Analysis
2.7. Statistical Analysis
3. Results
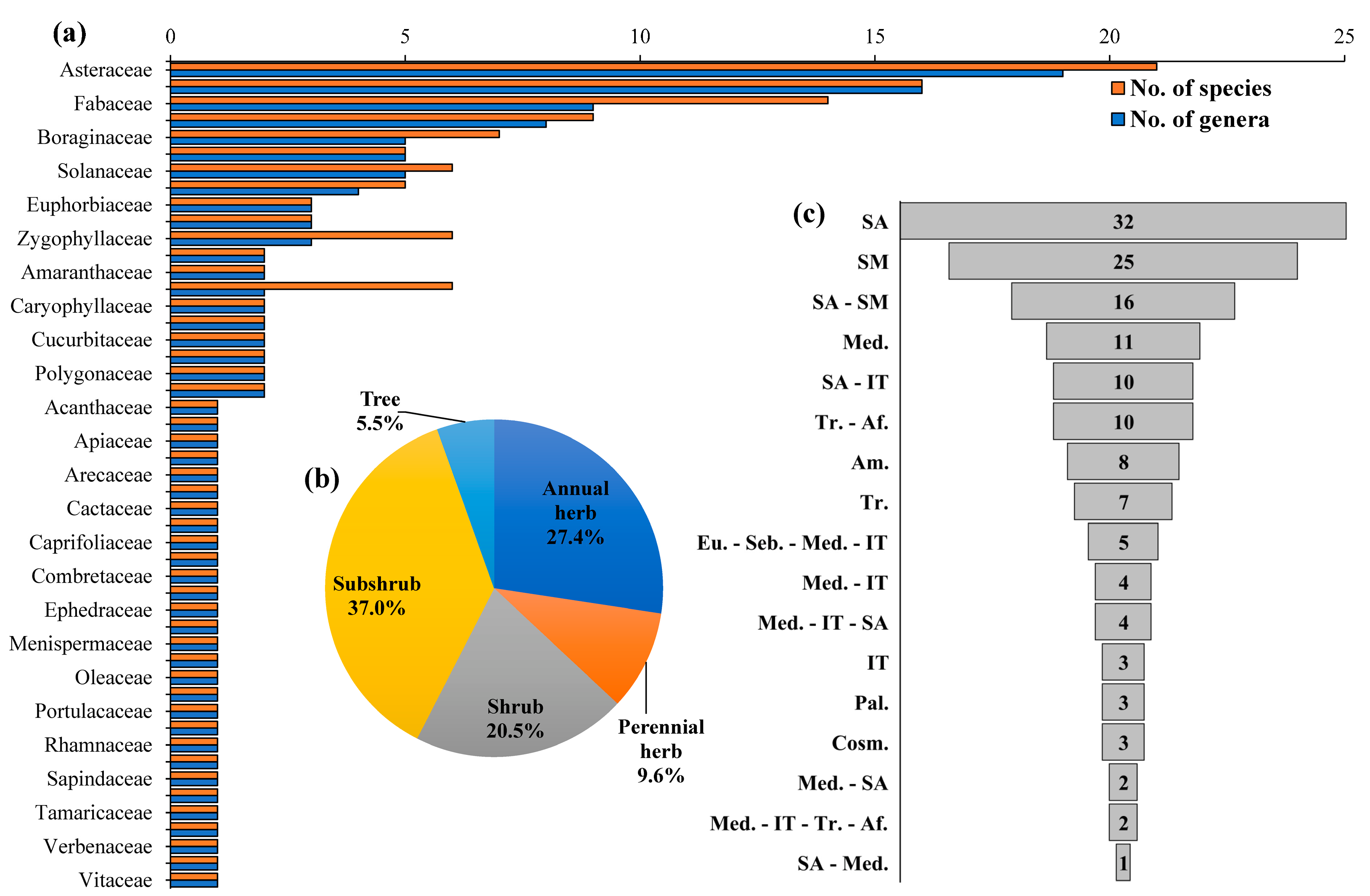
3.1. Floristic Diversity and Phytogeography
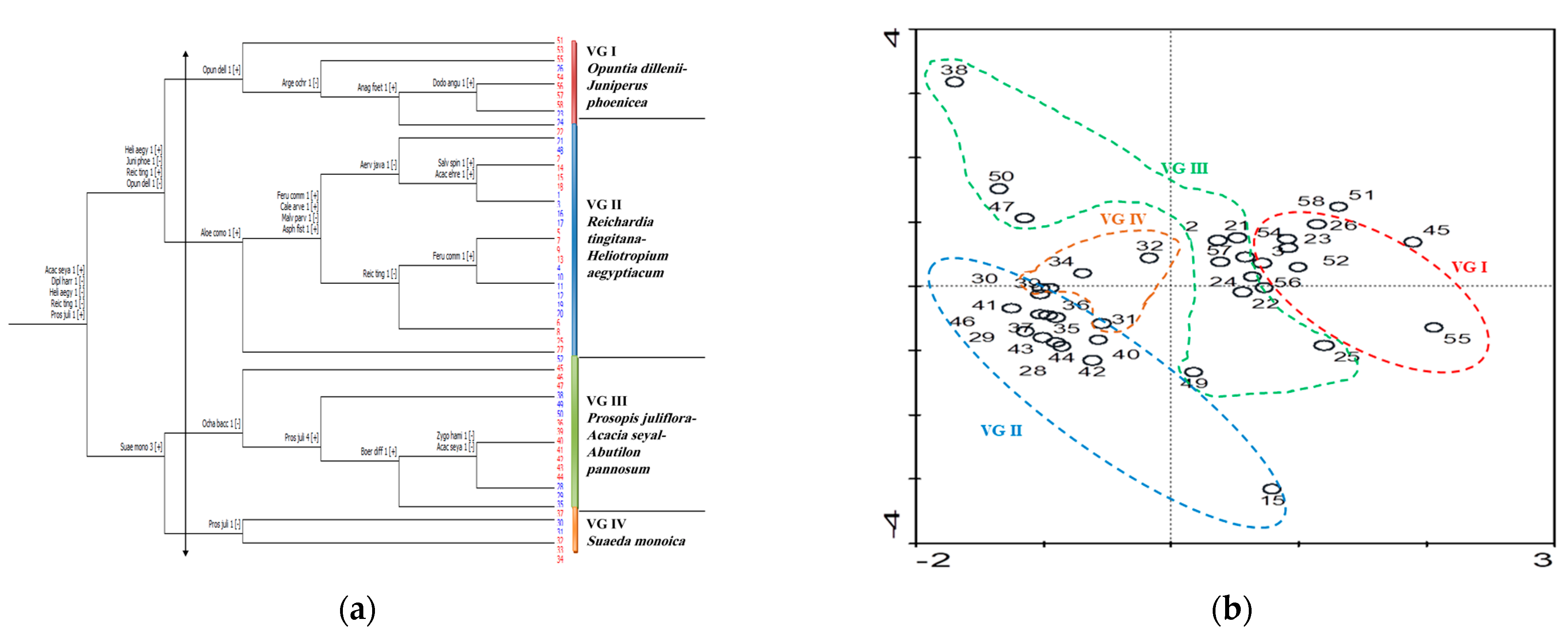
3.2. Vegetation Structure
3.3. Correaltion Analyses
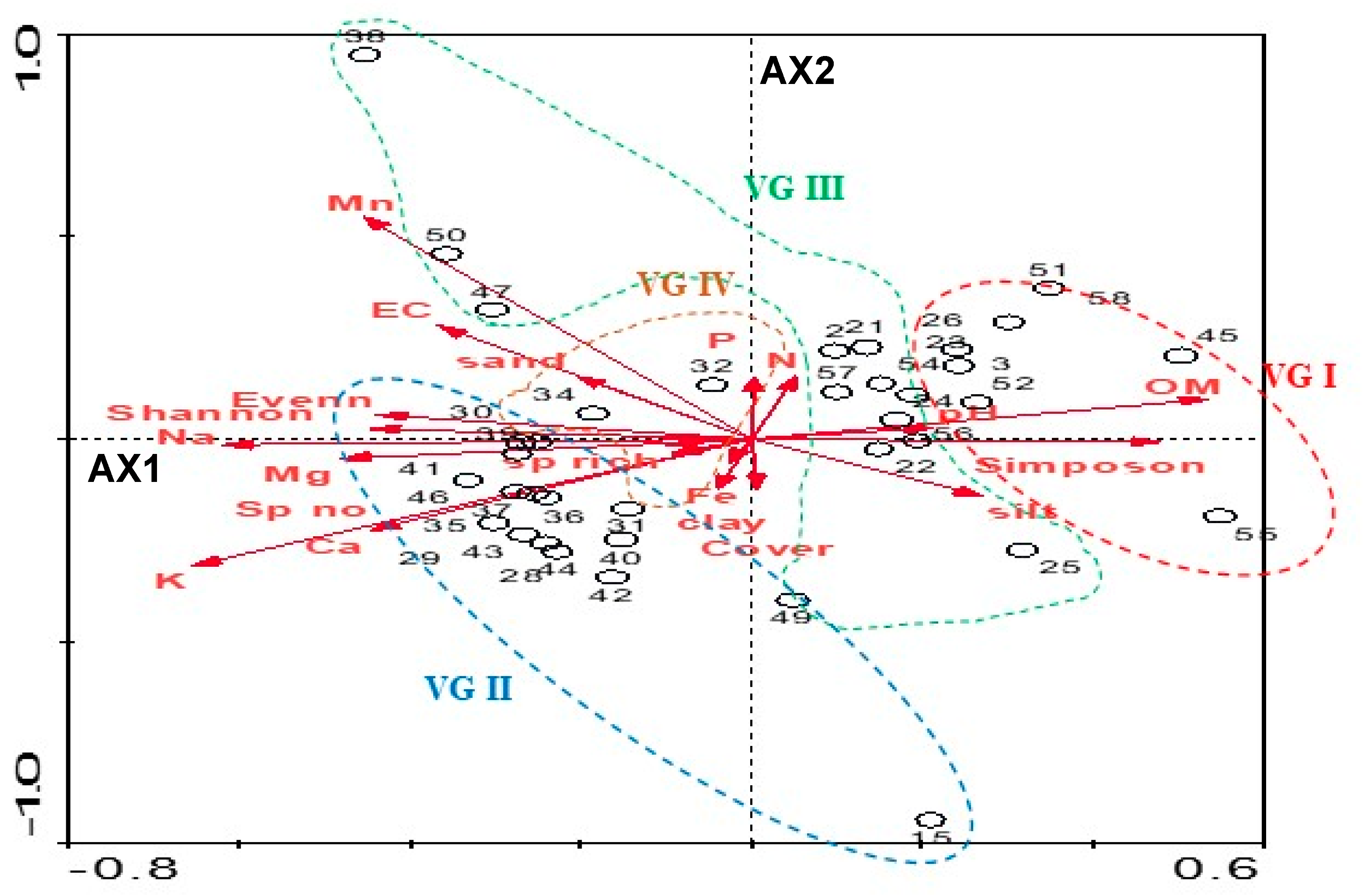
3.3.1. Relationships between Soil Variables and Distribution of Sampling Stands
3.3.2. Relationships between Diversity Indices and Soil Variables
3.4. Soil-Plant Association Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- AlNafie, A.H. Phytogeography of Saudi Arabia. Saudi J. Biol. Sci. 2008, 15, 159–176. [Google Scholar]
- Chaudhary, S.A. Flora of the Kingdom of Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 2001; Volume 2, p. 542. [Google Scholar]
- Collenette, S. Wildflowers of Saudi Arabia; National Commission for Wildlife Conservation and Development (NCWCD): Riyadh, Saudi Arabia, 1999; p. 799. [Google Scholar]
- Collenette, S. A Checklist of Botanical Species in Saudi Arabia; International Asclepiad Society: West Sussex, UK, 1998. [Google Scholar]
- Collenette, S. An Illustrated Guide to the Flowers of Saudi Arabia; Scorpion Publishing Ltd: Riyadh, Saudi Arabia, 1985. [Google Scholar]
- Al-Aklabi, A.; Al-Khulaidi, A.W.; Hussain, A.; Al-Sagheer, N. Main vegetation types and plant species diversity along an altitudinal gradient of Al Baha region, Saudi Arabia. Saudi J. Biol. Sci. 2016, 23, 687–697. [Google Scholar] [CrossRef] [PubMed]
- El-Sheikh, M.A.; Thomas, J.; Alatar, A.A.; Hegazy, A.K.; Abbady, G.A.; Alfarhan, A.H.; Okla, M.I. Vegetation of Thumamah Nature Park: A managed arid land site in Saudi Arabia. Rendiconti Lincei 2013, 24, 349–367. [Google Scholar] [CrossRef]
- Zohary, M. Geobotanical Foundations of the Middle East; Swets & Zeitlinger: Lisse, The Netherlands, 1973. [Google Scholar]
- Al-Namazi, A.A.; Al-Khulaidi, A.W.A.; Algarni, S.; Al-Sagheer, N.A. Natural plant species inventory of hotspot areas in Arabian Peninsula: Southwest Al-Baha region, Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 3309–3324. [Google Scholar] [CrossRef]
- Thomas, J.; El-Sheikh, M.A.; Alatar, A.A. Endemics and endangered species in the biodiversity hotspot of the Shada Mountains, Saudi Arabia. J. Arid. Land 2017, 9, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Al-Abbasi, T.; Al-Farhan, A.; Al-Khulaidi, A.; Hall, M.; Llewellyn, O.; Miller, A.; Patzelt, A. Important plant areas in the Arabian Peninsula. Edinb. J. Bot. 2010, 67, 25–35. [Google Scholar] [CrossRef]
- Hobohm, C.; Janišová, M.; Jansen, J.; Bruchmann, I.; Deppe, U. Biogeography of endemic vascular plants–overview. In Endemism in Vascular Plants; Springer Nature: Cham, Switzerland, 2014; pp. 85–163. [Google Scholar]
- Elaidarous, A.A.; Osman, H.E.; Galal, T.M.; El-Morsy, M.H. Vegetation–environment relationship and floristic diversity of Wadi Al-Sharaea, Makkah Province, Saudi Arabia. Rendiconti Lincei. Scienze Fisiche e Naturali 2022, 33, 169–184. [Google Scholar] [CrossRef]
- Abdel Khalik, K.; Al-Gohary, I.; Al-Sodany, Y. Floristic composition and vegetation: Environmental relationships of Wadi Fatimah, Mecca, Saudi Arabia. Arid. Land Res. Manag. 2017, 31, 316–334. [Google Scholar] [CrossRef]
- Abdel Khalik, K.; El-Sheikh, M.; El-Aidarous, A. Floristic diversity and vegetation analysis of wadi Al-Noman, Mecca, Saudi Arabia. Turk. J. Bot. 2013, 37, 894–907. [Google Scholar] [CrossRef]
- Ghazal, A.M.F. Vegetation patterns and plant communities distribution along an altitudinal gradient at Asir Mountain, southwest Saudi Arabia. Pak. J. Bot. 2015, 47, 1377–1389. [Google Scholar]
- Al-Yemeni, M.; Sher, H. Biological spectrum with some other ecological attributes of the flora and vegetation of the Asir Mountain of South West, Saudi Arabia. Afr. J. Biotechnol. 2010, 9, 5550–5559. [Google Scholar]
- Abd El-Ghani, M.M. Vegetation along a transect in the Hijaz mountains (Saudi Arabia). J. Arid Environ. 1996, 32, 289–304. [Google Scholar] [CrossRef]
- Alharthi, A.; El-Sheikh, M.A.; Elhag, M.; Alatar, A.A.; Abbadi, G.A.; Abdel-Salam, E.M.; Arif, I.A.; Baeshen, A.A.; Eid, E.M. Remote sensing of 10 years changes in the vegetation cover of the northwestern coastal land of Red Sea, Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 3169–3179. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, H. Flora of Kingdom of Saudi Arabia vol. I, II and III; Ministry of Agriculture and Water, National Herbarium, National and Water Research Center: Riyadh, Saudi Arabia, 1999–2001. [Google Scholar]
- Kent, M. Vegetation Description and Data Analysis: A Practical Approach, 2nd ed.; John Wiley & Sons: Chichester, UK, 2012; p. 428. [Google Scholar]
- Ellenberg, D.; Mueller-Dombois, D. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974; p. 547. [Google Scholar]
- Pielou, E.C. Ecological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 1975. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific: Oxford, UK, 1989; p. 368. [Google Scholar]
- Hill, M.O. TWINSPAN: A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-way Table by Classification of the Individuals and Attributes; Section of Ecology and Systematics, Cornell University: New York, NY, USA, 1979; p. 90. [Google Scholar]
- Hill, M.O. DECORANA: A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging; Section of Ecology and Systematics, Cornell University: New York, NY, USA, 1979; p. 52. [Google Scholar]
- Ter Braak, C.J.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Wageningen University: Wageningen, The Netherlands, 2002; Available online: http://www.canoco5.com/ (accessed on 3 December 2022).
- Migahid, A.M. Flora of Saudi Arabia; Riyadh University Publications: Riyadh, Saudi Arabia, 1996; Volume 1. [Google Scholar]
- Migahid, A.M.; El-Sheikh, A.M. A contribution to the study of vegetation in western and north western Saudi Arabia. Proc. Saudi Biol. Soc. 1983, 6, 115–150. [Google Scholar]
- Migahid, M.A.; El-Sheikh, A.M. The Natural Vegetation of Al-Rabadha Area of Western Saudi Arabia. Proc. Saudi Biol. Soc. 1983, 6, 151–174. [Google Scholar]
- Al-Sherif, E.A.; Ayesh, A.M.; Rawi, S.M. Floristic composition, life form and chorology of plant life at Khulais region, Western Saudi Arabia. Pak. J. Bot. 2013, 45, 29–38. [Google Scholar]
- Al Wadie, H. Floristic composition and vegetation of Wadi Talha, Aseer mountains, south west Saudi Arabia. J. Biol. Sci. 2002, 2, 285–288. [Google Scholar]
- Heneidy, S.Z.; Bidak, L.M. Biodiversity of the Plant Species in Bisha, Asir Region, Southwestern Saudi Arabia. Pak. J. Biol. Sci. 2001, 4, 1323–1330. [Google Scholar]
- Hegazy, A.; El-Demerdash, M.; Hosni, H. Vegetation, species diversity and floristic relations along an altitudinal gradient in south-west Saudi Arabia. J. Arid Environ. 1998, 38, 3–13. [Google Scholar] [CrossRef]
- El-Sheikh, M.A.; Al-Shehri, M.A.; Alfarhan, A.H.; Alatar, A.A.; Rajakrishnan, R.; Al-Rowaily, S.L. Threatened Prunus arabica in an ancient volcanic protected area of Saudi Arabia: Floristic diversity and plant associations. Saudi J. Biol. Sci. 2019, 26, 325–333. [Google Scholar] [CrossRef]
- Alatar, A.A.; El-Sheikh, M.A.R.; Thomas, J.; Hegazy, A.K.; El Adawy, H.A. Vegetation, Floristic Diversity, and Size-Classes of Acacia gerrardii in an Arid Wadi Ecosystem. Arid. Land Res. Manag. 2015, 29, 335–359. [Google Scholar] [CrossRef]
- El-Sheikh, M.A. Population structure of woody plants in the arid cloud forests of Dhofar, southern Oman. Acta Bot. Croat. 2013, 72, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Alatar, A.A.; El-Sheikh, M.A.; Thomas, J. Vegetation analysis of Wadi Al-Jufair, a hyper-arid region in Najd, Saudi Arabia. Saudi J. Biol. Sci. 2012, 19, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sheikh, M.A.; Abbadi, G.A.; Bianco, P.M. Vegetation ecology of phytogenic hillocks (nabkhas) in coastal habitats of Jal Az-Zor National Park, Kuwait: Role of patches and edaphic factors. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 832–840. [Google Scholar] [CrossRef]
- Al-Turki, T.; Al-Olayan, H. Contribution to the flora of Saudi Arabia: Hail region. Saudi J. Biol. Sci. 2003, 10, 190–222. [Google Scholar]
- El-Ghanim, W.M.; Hassan, L.M.; Galal, T.M.; Badr, A. Floristic composition and vegetation analysis in Hail region north of central Saudi Arabia. Saudi J. Biol. Sci. 2010, 17, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Takhtadzhian, A.L. The Floristic Regions of the World; University of California Press: Berkeley, CA, USA, 1988; p. 522. [Google Scholar]
- Alharthi, S.T.; El-Sheikh, M.A.; Alfarhan, A.A. Biological change of western Saudi Arabia: Alien plants diversity and their relationship with edaphic variables. J. King Saud Univ. Sci. 2023, 35, 102496. [Google Scholar] [CrossRef]
- Thomas, J.; El-Sheikh, M.A.; Alfarhan, A.H.; Alatar, A.A.; Sivadasan, M.; Basahi, M.; Al-Obaid, S.; Rajakrishnan, R. Impact of alien invasive species on habitats and species richness in Saudi Arabia. J. Arid Environ. 2016, 127, 53–65. [Google Scholar] [CrossRef]
- Rahmonov, O.; Abramowicz, A.; Pukowiec-Kurda, K.; Fagiewicz, K. The link between a high-mountain community and ecosystem services of juniper forests in Fann Mountains (Tajikistan). Ecosyst. Serv. 2021, 48, 101255. [Google Scholar] [CrossRef]
- Brooks, W.; Mandil, K. Vegetation dynamics in the Asir woodlands of south-western Saudi Arabia. J. Arid Environ. 1983, 6, 357–362. [Google Scholar] [CrossRef]
- El-Sheikh, M.A.; Thomas, J.; Arif, I.A.; El-Sheikh, H.M. Ecology of inland sand dunes “nafuds” as a hyper-arid habitat, Saudi Arabia: Floristic and plant associations diversity. Saudi J. Biol. Sci. 2021, 28, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Elmefregy, M.; El-Sheikh, M. Ecological status of sand binder plant white saxaul (Haloxylon persicum) at the managed area of Al-Qassim, Saudi Arabia: Plant associations and population structure. Appl. Ecol. Environ. Res. 2020, 18, 2781–2794. [Google Scholar] [CrossRef]
- Valentini, E.; Taramelli, A.; Cappucci, S.; Filipponi, F.; Nguyen Xuan, A. Exploring the Dunes: The Correlations between Vegetation Cover Pattern and Morphology for Sediment Retention Assessment Using Airborne Multisensor Acquisition. Remote Sens. 2020, 12, 1229. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VG * | # Stands | % Study Area | Stands | Habitats (%) + | 1st Dominant sp. | C (%) | P (%) | 2nd Dominant sp. | C (%) | P (%) | No. Species |
|---|---|---|---|---|---|---|---|---|---|---|---|
| VG I | 11 | 18.96 | 22–24, 26, 51, 53–58 | F = 0 M = 100 P = 0 W = 0 C = 0 R = 0 | Opuntia ficus-indica | 90 | 67 | Juniperus phoenicea | 30 | 67 | Native: 32 Casual: 0 Naturalized: 11 Invasive: 2 |
| VG II | 25 | 43.10 | 1–21, 25, 27, 48, 52 | F = 24 M = 40 P = 0 W = 28 C = 0 R = 8 | Reichardia tingitana | 10 | 63 | Heliotropium aegyptiacum | 5 | 58 | Native: 50 Casual: 2 Naturalized: 23 Invasive: 2 |
| VG III | 17 | 29.31 | 28, 29, 35–39, 40–47, 49, 50 | F = 17.65 M = 0 P = 5.88 W = 64.71 C = 11.67 R = 0 | Prosopis juliflora | 85 | 76 | Acacia seyal | 30 | 47 | Native: 38 Casual: 2 Naturalized: 33 Invasive: 2 |
| Abutilon pannosum | 20 | 35 | |||||||||
| VG IV | 5 | 5.63 | 30–34 | F = 0 M = 0 P = 0 W = 100 C = 0 R = 0 | Suaeda monoica | 43 | 80 | Acacia ehrenbergiana | 10 | 60 | Native: 6 Casual: 0 Naturalized: 8 Invasive: 1 |
| Variable | Axis 1 | Axis 2 |
|---|---|---|
| Diversity indices | ||
| Species number | −0.10 | −0.04 |
| Species coverage (%) | 0.01 | −0.11 |
| Species richness | −0.10 | −0.02 |
| Species evenness | −0.43 * | 0.10 |
| Shannon (Ĥ) | −0.44 * | 0.02 |
| Simpson-C | 0.50 * | −0.01 |
| Soil | ||
| pH | 0.21 | 0.02 |
| EC (mmohs cm−3) | −0.40 * | 0.26 |
| Bulk soil (%) | ||
| Sand | −0.20 | 0.14 |
| Silt | 0.30 | −0.13 |
| Clay | −0.04 | −0.12 |
| Organic matter | 0.52 ** | 0.10 |
| Minerals | ||
| N (mg g−1) | 0.003 | 0.14 |
| P (mg g−1) | 0.05 | 0.142 |
| K (mg g−1) | −0.64 *** | −0.29 |
| Mg (ppm) | −0.50 * | −0.05 |
| Ca (ppm) | −0.44 * | −0.21 |
| Fe (ppm) | −0.03 | −0.10 |
| Na (ppm) | −0.61 *** | −0.0.15 |
| Mn (ppm) | −0.44 | 0.51 ** |
| Variable | pH | Sand (%) | Silt (%) | Clay (%) | OM (%) | Fe (ppm) | Mn (ppm) |
|---|---|---|---|---|---|---|---|
| Sp. number | −0.023 | −0.30 * | 0.33 * | 0.01 | 0.21 | 0.02 | −0.10 |
| Plant coverage (%) | −0.4 ** | 0.30 | −0.24 | −0.23 | −0.33 * | −0.34 ** | 0.30 * |
| Sp. richness | 0.20 | −0.35 ** | 0.40 ** | 0.20 | 0.33 * | 0.20 | −0.22 |
| Evenness | 0.30 * | −0.26 | 0.23 | 0.30 * | 0.30 * | 0.31 * | −0.23 * |
| Shannon (Ĥ) | 0.20 | −0.34 | 0.30 ** | 0.25 | 0.40 ** | 0.30 * | −0.32 * |
| Simpson-C | −0.26 | 0.30 * | −0.30 * | −0.30 * | −0.30 * | −0.30 * | 0.40 ** |
| Variable | VG I | VG II | VG III | VG IV | F-Value | p-Value |
|---|---|---|---|---|---|---|
| Diversity indices | ||||||
| Species number | 6.82 ± 2.70 | 9.84 ± 2.90 | 8.8 ± 2.61 | 4.4 ± 1.70 | 7.2 | 0.00 *** |
| Species coverage (%) | 86.73 ± 30.85 | 32.84 ± 31.97 | 94.24 ± 33.65 | 81.60 ± 37.85 | 14.6 | 0.00 *** |
| Species richness | 1.40 ± 0.10 | 2.70 ± 0.10 | 1.80 ± 0.70 | 0.90 ±0.50 | 16.6 | 0.00 *** |
| Species evenness | 0.50 ±0.16 | 0.84 ± 0.14 | 0.62 ± 0.16 | 0.70 ± 0.10 | 15.8 | 0.00 *** |
| Shannon (Ĥ) | 0.40 ± 0.15 | 0.81 ± 0.16 | 0.60 ± 0.18 | 0.40 ± 0.10 | 22.11 | 0.00 *** |
| Simpson-C | 0.60 ± 0.15 | 0.20 ± 0.12 | 0.40 ± 0.16 | 0.45 ± 0.10 | 22.6 | 0.00 *** |
| Soil | ||||||
| pH | 7.70 ± 0.34 | 7.82 ± 0.25 | 7.50 ± 0.30 | 7.9 ± 0.20 | 6.9 | 0.001 *** |
| EC (mmohs cm−3) | 0.30 ± 0.50 | 0.33 ± 0.42 | 1.30 ± 1.54 | 0.14 ± 0.10 | 4.5 | 0.007 *** |
| Bulk soil (%) | ||||||
| Sand | 72.01 ± 7.8 | 62.10 ± 19.5 | 70.90 ± 13.3 | 80.50 ± 12.12 | 2.71 | 0.054 * |
| Silt | 17.5 ± 6.90 | 24.7 ± 15.02 | 17.7 ± 11.10 | 8.9 ± 5.60 | 3 | 0.038 * |
| Clay | 10.5 ± 2.3 | 13.2 ± 5.3 | 11.5 ± 3.6 | 10.6 ± 6.5 | 1.3 | 0.30 |
| Organic matter | 8.28 ± 3.72 | 8.70 ± 2.90 | 5.58 ± 3.80 | 2.19 ± 1.60 | 7.5 | 0.00 *** |
| Minerals | ||||||
| N (mg g−1) | 4.50 ± 1.80 | 5.80 ± 4.04 | 5.60 ± 2.90 | 2.92 ± 1.02 | 1.4 | 0.30 |
| P (mg g−1) | 0.2 ± 0.01 | 0.2 ± 0.02 | 0.2 ± 0.01 | 0.1 ± 0.03 | 0.8 | 0.53 |
| K (mg g−1) | 2.70 ± 1.99 | 3.10 ± 1.97 | 7.84 ± 2.70 | 5.02 ± 1.94 | 13.4 | 0.00 *** |
| Mg (ppm) | 3.54 ± 1.3 | 2.53 ± 1.4 | 5.50 ± 1.93 | 2.50 ± 1.5 | 7.6 | 0.00 *** |
| Ca (ppm) | 13.80 ± 6.58 | 12.75 ± 3.10 | 20.20 ± 4.43 | 9.30 ± 3.30 | 2.73 | 0.053 * |
| Fe (ppm) | 0.12 ± 0.01 | 0.70 ± 0.05 | 0.10 ± 0.03 | 0.30 ± 0.11 | 3.7 | 0.018 * |
| Na (ppm) | 9.83 ± 2.23 | 12.01 ± 3.10 | 15.72 ± 5.14 | 11.80 ± 0.97 | 1.5 | 0.227 |
| Mn (ppm) | 0.04 ± 0.01 | 0.01 ± 0.01 | 0.07 ± 0.01 | 0.04 ± 0.01 | 7.2 | 0.00 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharthi, S.T.; El-Shiekh, M.A.; Alfarhan, A.A. Alien Plant Invasions of the Natural Habitat in the Western Region of Saudi Arabia: Floristic Diversity and Vegetation Structure. Diversity 2023, 15, 309. https://doi.org/10.3390/d15030309
Alharthi ST, El-Shiekh MA, Alfarhan AA. Alien Plant Invasions of the Natural Habitat in the Western Region of Saudi Arabia: Floristic Diversity and Vegetation Structure. Diversity. 2023; 15(3):309. https://doi.org/10.3390/d15030309
Chicago/Turabian StyleAlharthi, Saud T., Mohamed A. El-Shiekh, and Ahmed A. Alfarhan. 2023. "Alien Plant Invasions of the Natural Habitat in the Western Region of Saudi Arabia: Floristic Diversity and Vegetation Structure" Diversity 15, no. 3: 309. https://doi.org/10.3390/d15030309
APA StyleAlharthi, S. T., El-Shiekh, M. A., & Alfarhan, A. A. (2023). Alien Plant Invasions of the Natural Habitat in the Western Region of Saudi Arabia: Floristic Diversity and Vegetation Structure. Diversity, 15(3), 309. https://doi.org/10.3390/d15030309