
Analysis of the Anomalies in a Middle Eocene Shell of Neochelys (Pleurodira, Podocnemididae) from the Duero Basin (Zamora, Spain)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
Institutional Abbreviations
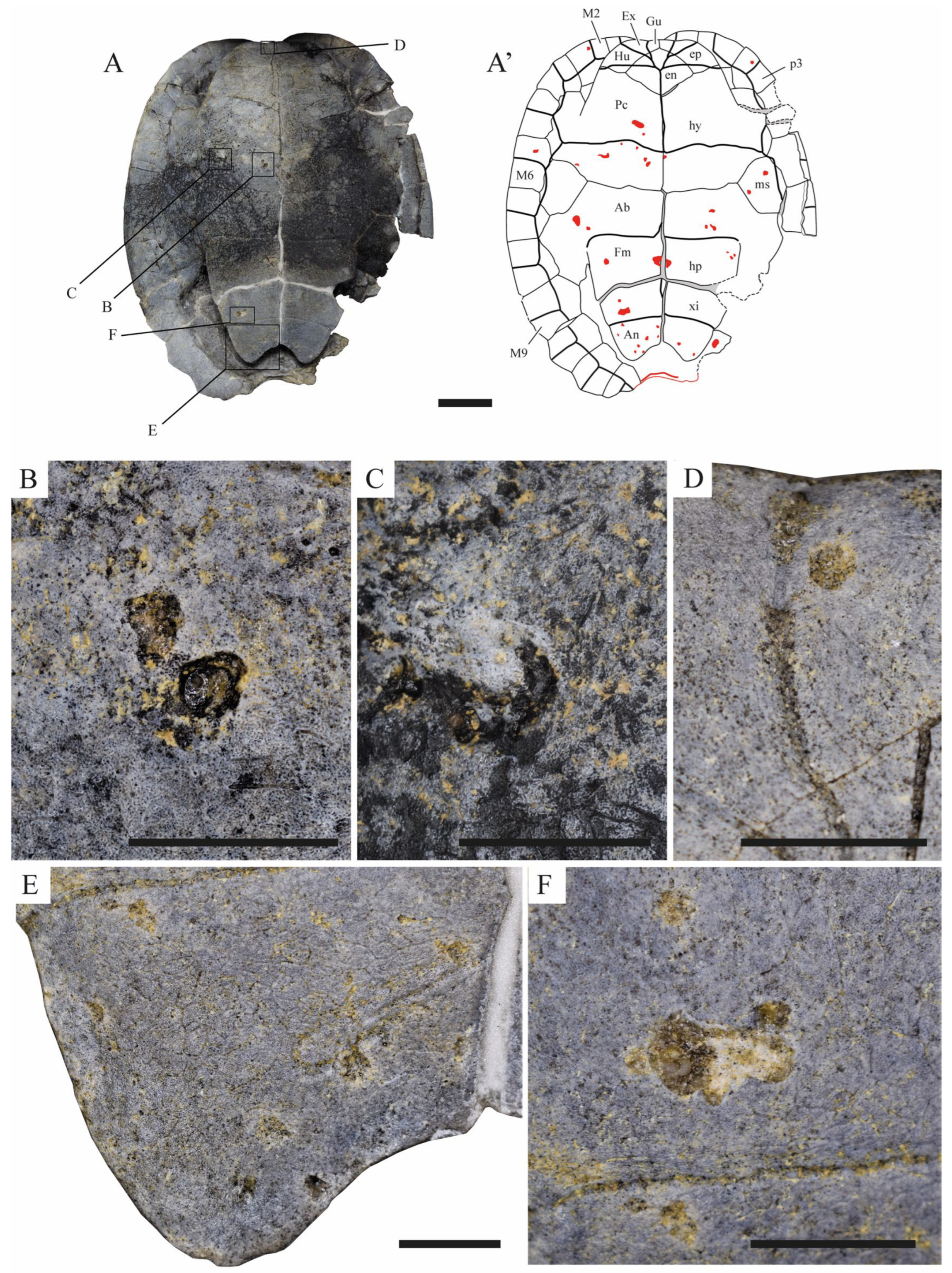
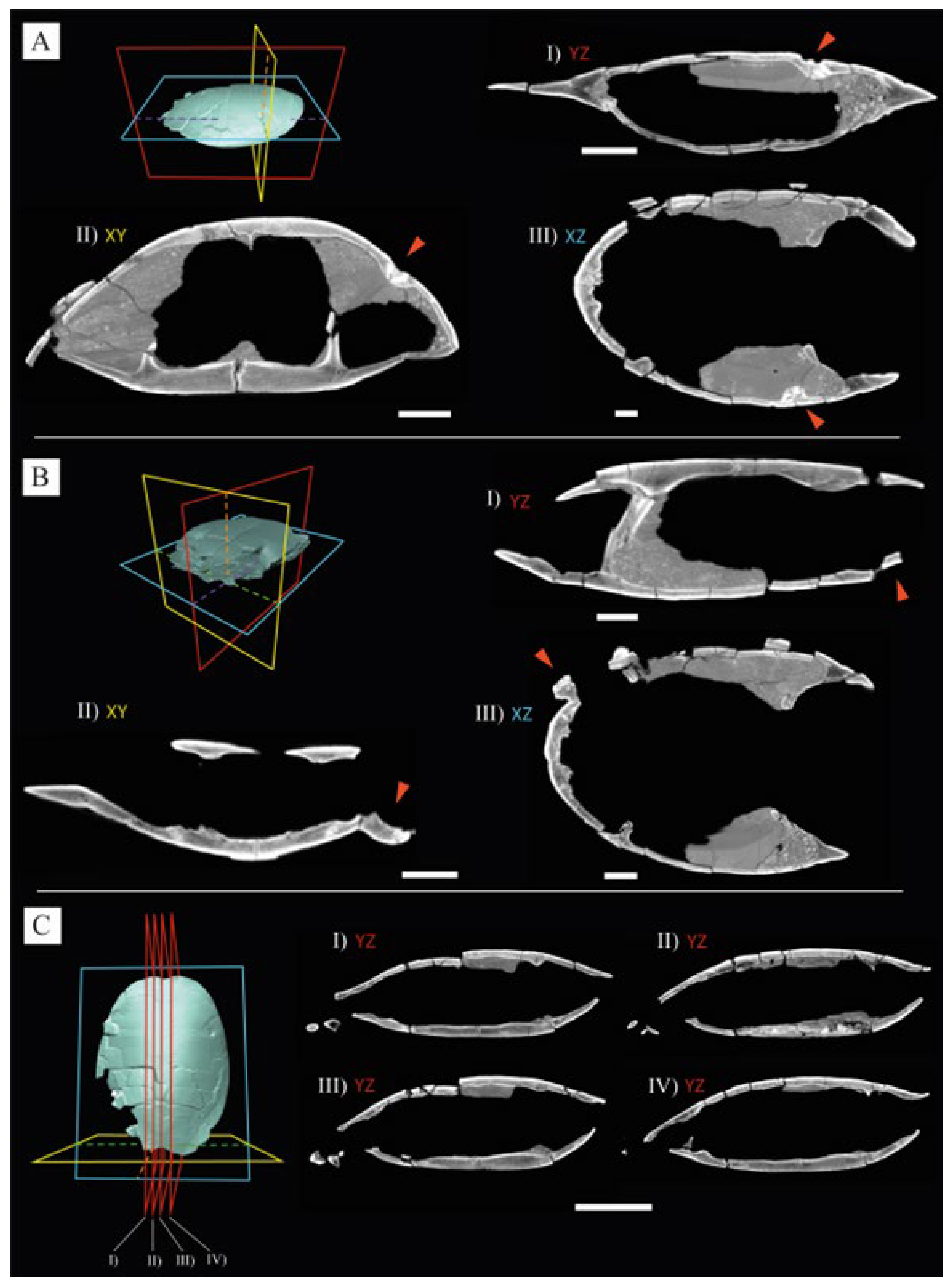
3. Results
4. Discussions
4.1. Etiology of the Anomalous Shell Marks of STUS 14024
4.2. Etiology of the Anomaly of the Posterior Region of the Carapace STUS 14024

5. Pathogenesis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collareta, A.; Casati, S.; Zuffi, M.A.; Di Cencio, A. First authentic record of the freshwater turtle Mauremys from the upper Pliocene of Italy, with a new occurrence of the rarely reported ichnotaxon Thatchtelithichnus holmani. Carnets Geol. 2020, 20, 301–313. [Google Scholar] [CrossRef]
- Reolid, M.; Santos, A.; Mayoral, E. Grazing activity as taphonomic record of necrobiotic interaction: A case study of a sea turtle carapace from the Upper Jurassic of the Prebetic (south Spain). Rev. Mex. de Cienc. Geol. 2015, 32, 21–28. [Google Scholar]
- Jagt, J.W.; Deckers, M.J.; De Leebeeck, M.; Donovan, S.K.; Nieuwenhuis, E. Episkeletozoans and bioerosional ichnotaxa on isolated bones of Late Cretaceous mosasaurs and cheloniid turtles from the Maastricht area, the Netherlands. Geologos 2020, 26, 39–49. [Google Scholar] [CrossRef]
- Zonneveld, J.P.; AbdelGawad, M.K.; Miller, E.R. Ectoparasite borings, mesoparasite borings, and scavenging traces in early Miocene turtle and tortoise shell: Moghra Formation, Wadi Moghra, Egypt. J. Paleontol. 2022, 96, 304–322. [Google Scholar] [CrossRef]
- Hutchison, J.H.; Frye, F. Evidence of pathology in early Cenozoic turtles. PaleoBios 2001, 21, 12–19. [Google Scholar]
- Homer, B.L.; Berry, K.H.; Brown, M.B.; Ellis, G.; Jacobson, E.R. Pathology of diseases in wild desert tortoises from California. J. Wildl. Dis. 1998, 34, 508–523. [Google Scholar] [CrossRef]
- Scheyer, T.M.; Delfino, M.; Klein, N.; Bunbury, N.; Fleischer-Dogley, F.; Hansen, D.M. Trophic interactions between larger crocodylians and giant tortoises on Aldabra Atoll, Western Indian Ocean, during the Late Pleistocene. R. Soc. Open Sci. 2018, 5, 171800. [Google Scholar] [CrossRef] [Green Version]
- Garner, M.M.; Herrington, R.; Howerth, E.W.; Homer, B.L.; Nettles, V.F.; Isaza, R.; Shotts, E.B.; Jacobson, E.R. Shell disease in river cooters (Pseudemys concinna) and yellow–bellied turtles (Trachemys scripta) in a Georgia (USA) lake. J. Wildl. Dis. 1997, 33, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Pereda-Suberbiola, X.; Astibia, H.; Murelaga, X.; Elorza, J.J.; Gomez-Alday, J.J. Taphonomy of the Late Cretaceous dinosaur-bearing beds of the Laño Quarry (Iberian Peninsula). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 157, 247–275. [Google Scholar] [CrossRef]
- Hayashi, R.; Chan, B.K.K.; Simon-Blecher, N.; Watanabe, H.; Guy-Haim, T.; Yonezawa, T.; Levy, Y.; Shuto, T.; Achituv, Y. Phylogenetic position and evolutionary history of the turtle and whale barnacles (Cirripedia: Balanomorpha: Coronuloidea). Mol. Phylogenet. Evol. 2013, 67, 9–14. [Google Scholar] [CrossRef]
- Agha, M.; Riley, M.K.; Sanford, E.; Carlton, J.T.; Newman, W.A.; Todd, B.D. A review of epizoic barnacles reported from freshwater turtles with a new record from California. Herpetol. Rev. 2018, 49, 25–28. [Google Scholar]
- Lafortune, M.; Wellehan, J.F.; Terrell, S.P.; Jacobson, E.R.; Heard, D.; Kimbrough, J.W. Shell and systemic hyalohyphomycosis in Fly River turtles, Carettochelys insculpta, caused by Paecilomyces lilacinus. J. Herpetol. Med. Surg 2005, 15, 15–19. [Google Scholar] [CrossRef]
- Pfaller, J.B.; Frick, M.G.; Reich, K.J.; Williams, K.L.; Bjorndal, K.A. Carapace epibionts of loggerhead turtles (Caretta caretta) nesting at Canaveral National Seashore, Florida. J. Nat. Hist. 2008, 42, 1095–1102. [Google Scholar] [CrossRef]
- McCoy, J.C.; Failey, E.L.; Price, S.J.; Dorcas, M.E. An assessment of leech parasitism on semi-aquatic turtles in the western Piedmont of North Carolina. Southeast. Nat. 2007, 6, 191–202. [Google Scholar] [CrossRef]
- Ryan, T.J.; Lambert, A. Prevalence and colonization of Placobdella on two species of freshwater turtles (Graptemys geographica and Sternotherus odoratus). J. Herpetol. 2005, 39, 284–287. [Google Scholar] [CrossRef]
- Ortega, F.; Armenteros, I.; de Celis, A.; Escaso, F.; Huerta, P.; Martín de Jesús, S.; Narváez, I.; Pérez-García, A.; Sanz, J.L. Crocodyliformes and Testudines from the Eocene of the Duero Basin (northwestern Spain): An update of their diversity and stratigraphic context. Hist Biol. 2022, 34, 1560–1581. [Google Scholar] [CrossRef]
- Jiménez-Fuentes, E. Predación crocodiliana a quelonios. Un Neochelys (Pelomedusidae), del Eoceno de Zamora, lisiado por un Asiatosuchus. Stud. Geol. Salmant. 2003, 39, 11–23. [Google Scholar]
- Hallock, L.A.; McMillan, A.; Wiles, G.J. Draft Periodic Status Review for the Western Pond Turtle in Washington; Washington Department of Fish and Wildlife: Olympia, WA, USA, 2016; p. 18. [Google Scholar]
- Rothschild, B.M.; Martin, L.D. Skeletal impact of disease. Bull. N. M. Mus. Nat. Hist. Sci. 2006, 33, 1–226. [Google Scholar]
- Zonneveld, J.P.; Bartels, W.S. The occurrence of bone modification features in the carapace and plastron of the extant red-eared Slider Trachemys scripta elegans (Wied-Neuwied, 1839): Implications for paleoecological analyses of fossil turtle assemblages. Palaios 2022, 37, 499–519. [Google Scholar] [CrossRef]
- Bader, K.S.; Hasiotis, S.T.; Martin, L.D. Application of forensic science techniques to trace fossils on dinosaur bones from a quarry in the Upper Jurassic Morrison Formation, northeastern Wyoming. Palaios 2009, 24, 140–158. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification; Springer: Dordrecht, The Netherlands, 2016; p. 359. [Google Scholar] [CrossRef]
- Höpner, S.; Bertling, M. Holes in bones: Ichnotaxonomy of bone borings. Ichnos 2017, 24, 259–282. [Google Scholar] [CrossRef]
- Emmons, L.H. Jaguar predation on chelonians. J. Herpetol. 1989, 23, 311–314. [Google Scholar] [CrossRef]
- Frazier, J.G.; Winston, J.E.; Ruckdeschel, C.A. Epizoan communities on marine turtles. III. Bryozoa. Bull. Mar. Sci. 1992, 51, 1–8. [Google Scholar]
- Jackson, C.G.; Ross, A. The occurrence of barnacles on the alligator snapping turtle, Macroclemys temminckii (Troost). J. Herpetol. 1971, 5, 188–189. [Google Scholar] [CrossRef]
- Bielecki, A.; Cichocka, J.; Jabłonski, A.; Jelén, I.; Ropelewska, E.; Biedunkiewicz, A.; Terlecki, J.; Nowakowski, J.J.; Pakulnicka, J.; Szlachciak, J. Coexistence of Placobdella costata (Fr. Müller, 1846) (Hirudinida: Glossiphoniidae) and mud turtle Emys orbicularis. Biologia 2012, 67, 731–738. [Google Scholar] [CrossRef]
- Jacobson, E.R. Infectious Diseases and Pathology of Reptiles: Color Atlas and Text; CRC Press: Boca Raton, FL, USA, 2007; p. 736. [Google Scholar] [CrossRef]
- Mader, D.R. Reptile Medicine and Surgery; Saunders Elsevier: St. Louis, MI, USA, 2006; p. 1242. [Google Scholar]
- Johnson, C.A.; Griffith, J.W.; Tenorio, P.; Hytrek, S.; Lang, C.M. Fatal trematodiasis in research turtles. Comp. Med. 1998, 48, 340–343. [Google Scholar]
- Schumacher, J. Fungal diseases of reptiles. Vet. Clin. N. Am.-Exot. Anim. Pract. 2003, 6, 327–335. [Google Scholar] [CrossRef]
- Granados, J.L.; Moreno, O.G.; Brieva, C.I. Lesiones ulcerativas cutaneas en tortugas dulceaquícolas. Rev. Med. Vet. Zoot. 2013, 60, 61–70. [Google Scholar]
- Fayolle, S.; Moriconi, C.; Oursel, B.; Koenig, C.; Suet, M.; Ficheux, S.; Logez, M.; Olivier, A. Epizoic algae distribution on the carapace and plastron of the European pond turtle (Emys orbicularis, Linnaeus, 1758): A study from the Camargue, France. Cryptogam. Algol. 2016, 37, 221–232. [Google Scholar] [CrossRef]
- Jiménez-Fuentes, E. Los reptiles fósiles del Valle del Duero: Podocnemis carbajosai nov. sp. del Eoceno de Salamanca. Estud. Geol. 1971, 27, 85–93. [Google Scholar]
- Erickson, B.R. Chelonivorous habits of the Paleocene crocodile Leidyosuchus formidabilis; Science Museum of Minnesota: Saint Paul, MN, USA, 1984; Volume 5, pp. 3–9. [Google Scholar]
- Antunes, M.T.; De Broin, F. Le Crétacé terminal de Beira Litoral, Portugal: Remarques stratigraphiques et écologiques, étude complémentaire de Rosasia soutoi (Chelonii, Bothremydidae). Earth Sci. 1998, 9, 153–200. [Google Scholar]
- Jiménez-Fuentes, E.; Martín de Jesús, S.; Mulas Alonso, E. Malformaciones y deformaciones patológicas en tortugas fósiles. Notas Informativas Sala de las Tortugas 1987, 3, 1–4. [Google Scholar]
- Mead, J.I.; Cubero, R.; Zamora, A.L.V.; Swift, S.L.; Laurito, C.; Gomez, L.D. Plio-Pleistocene Crocodylus (Crocodylia) from southwestern Costa Rica. Stud. Neotrop. Fauna Environ. 2006, 41, 1–7. [Google Scholar] [CrossRef]
- Steadman, D.W.; Franz, R.; Morgan, G.S.; Albury, N.A.; Kakuk, B.; Broad, K.; Franz, S.E.; Tinker, K.; Pateman, M.P.; Lott, T.A.; et al. Exceptionally well preserved late Quaternary plant and vertebrate fossils from a blue hole on Abaco, The Bahamas. Proc. Natl. Acad. Sci. USA 2007, 104, 19897–19902. [Google Scholar] [CrossRef] [Green Version]
- Lehman, T.M.; Wick, S.L. Chupacabrachelys complexus, n. gen. n. sp. (Testudines: Bothremydidae), from the Aguja Formation (Campanian) of West Texas. J. Vertebr. Paleontol. 2010, 30, 1709–1725. [Google Scholar] [CrossRef]
- Schwimmer, D.R. Bite marks of the giant crocodylian Deinosuchus on Late Cretaceous (Campanian) bones. N. M. Mus. Nat. Hist. Sci. Bull. 2010, 51, 183–190. [Google Scholar]
- MCCoy, M.R.; Karl, H.V.; Tichy, G.; Steinbacher, J.; Aigner, G.; Cemper-Kisslich, J. Radiological evaluation of a fossil turtle trauma from the Upper Jurassic of Eichstätt (Testudines: Cryptodira). Stud. Geol. Salmant. 2012, 48, 37–44. [Google Scholar]
- Noto, C.R.; Main, D.J.; Drumheller, S.K. Feeding traces and paleobiology of a Cretaceous (Cenomanian) crocodyliform: Example from the Woodbine Formation of Texas. PALAIOS 2012, 27, 105–115. [Google Scholar] [CrossRef]
- Smith, H.F.; Hutchison, J.H.; Townsend, K.E.B.; Adrian, B.; Jager, D. Morphological variation, phylogenetic relationships, and geographic distribution of the Baenidae (Testudines), based on new specimens from the Uinta Formation (Uinta Basin), Utah (USA). PLoS ONE 2017, 12, e0180574. [Google Scholar] [CrossRef] [Green Version]
- Pujos, F.; Salas-Gismondi, R. Predation of the giant Miocene caiman Purussaurus on a mylodontid ground sloth in the wetlands of proto-Amazonia. Biol. Lett. 2020, 16, 20200239. [Google Scholar] [CrossRef]
- Joyce, W.G. The first complete skeleton of Solnhofia parsonsi (Cryptodira, Eurysternidae) from the Upper Jurassic of Germany and its taxonomic implications. J. Paleontol. 2000, 74, 684–700. [Google Scholar] [CrossRef]
- Alonso-Santiago, L.; Alonso-Andres, L.; Jiménez-Fuentes, E. Análisis de varios casos de zoopaleopatología del Eoceno medio de Zamora (Espana). Stud. Geol. Salmant. 2006, 42, 97–112. [Google Scholar]
- Karl, H.-V.; Tichy, G. The structure of fossil teeth of chelonophagous crocodiles (Diapsida: Crocodylia). Stud. Geol. Salmant. 2004, 40, 115–124. [Google Scholar]
- Karl, H.-V. Bite traces in a turtle shell fragment from the Kimmeridgian (Upper Jurassic) of northern Germany. Stud. Geol. Salmant. 2012, 9, 25–30. [Google Scholar]
- Ortiz, R.M.; Plotkin, P.T.; Owens, D.W. Predation upon olive ridley sea turtles (Lepidochelys olivacea) by the American crocodile (Crocodylus acutus) at Playa Nancite, Costa Rica. Chelonian Conserv. Biol. 1997, 2, 585–586. [Google Scholar]
- Milàn, J.; Kofoed, J.; Bromley, R.G. Crocodylian-chelonian carnivory: Bite traces of Dwarf Caiman, Paleosuchus palpebrosus in red-eared slider, Trachemys scripta carapaces. M. Mus Nat. Hist. Sci. 2010, 51, 195–200. [Google Scholar]
- Drumheller-Horton, S.K. An Actualistic and Phylogenetic Approach to Identifying and Interpreting Crocodylian Bite Marks. Ph.D. Thesis, University of Iowa, Iowa City, IA, USA, 2012; p. 170. [Google Scholar] [CrossRef]
- Njau, J.K.; Blumenschine, R.J. A diagnosis of crocodile feeding traces on larger mammal bone, with fossil examples from the Plio-Pleistocene Olduvai Basin, Tanzania. J. Hum. Evol. 2006, 50, 142–162. [Google Scholar] [CrossRef]
- Drumheller, S.K.; Brochu, C.A. A Diagnosis of Alligator mississippiensis bite marks with comparisons to existing crocodylian datasets. Ichnos 2014, 21, 131–146. [Google Scholar] [CrossRef]
- Njau, J.; Gilbert, H. Standardizing terms for crocodile-induced bite marks on bone surfaces in light of the frequent bone modification equifinality found to result from crocodile feeding behavior, stone tool modification, and trampling. FOROST Occas. Publ. 2016, 3, 1–13. [Google Scholar]
- Lescano, J.; Quevedo, M.; Fernández, V. Enfermedad ulcerativa cutánea septicémica en una colección multi-especie de tortugas semiacuáticas. Rev. Investig. Vet. Perú 2013, 24, 561–564. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.H. Husbandry and medicine of aquatic reptiles. Semin. Avian Exot. Pet Med. 2004, 13, 223–228. [Google Scholar] [CrossRef]
- Aleksić-Kovačević, S.; Oezvegy, J.; Krstić, N.; Rusvai, M.; Jakab, C.; Stanimirović, Z.; Becskei, Z. Skin and skeletal system lesions of european pond turtles (Emys orbicularis) from natural habitats. Acta Vet. Hung. 2014, 62, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Assan Kasim, M.; Senthil Kumar, K.; Palanivelrajan, M. Shell rot infection in red eared turtle. Int. J. Adv. Biol. Res. 2017, 7, 634–635. [Google Scholar]
- Barnett, S. Shell infections: When there are chinks in the armor. In Terrapin Tales (USA); MAATS: Highland, MD, USA, 2003; pp. 18–42. [Google Scholar]
- Wallach, J.D. The pathogenesis and etiology of Ulcerative Shell Disease in turtles. J. Zoo Anim. Med. 1975, 6, 11–13. [Google Scholar] [CrossRef]
- Woodburn, D.B.; Kinsel, M.J.; Poll, C.P.; Langan, J.N.; Haman, K.; Gamble, K.C.; Madox, C.; Jeon, A.B.; Wellehan, J.F.X.; Ossiboff, R.J.; et al. Shell lesions associated with Emydomyces testavorans infection in freshwater aquatic turtles. Vet. Pathol. 2021, 58, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Lovich, J.E.; Gotte, S.W.; Ernst, C.H.; Harshbarger, J.C.; Laemmerzahl, A.F.; Gibbons, J.W. Prevalence and histopathology of shell disease in turtles from Lake Blackshear, Georgia. J. Wildl. Dis. 1996, 32, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.R.; Hernández-Gómez, O.; Krohn, A.R.; Mutlow, A.; Patterson, L.; Rosenblum, E.B.; Timmer, M.; Willis, J.; Bushell, J. Turtle Shell Disease Fungus (Emydomyces testavorans): First documented occurrence in California and prevalence in free-living turtles. Ichthyol. Herpetol. 2021, 109, 958–962. [Google Scholar] [CrossRef]
- Frye, F.L.; Williams, D.L. Self-Assessment Colour Review of Reptiles and Amphibians; Manson Publishing: Stanford-le-Hope, UK, 1995; p. 233. [Google Scholar]
- Rothschild, B.M.; Schultze, H.-P.; Pellegrini, R. Osseous and other hard tissue pathologies in turtles and abnormalities of mineral deposition. In Morphology and Evolution of Turtles, Vertebrate Paleobiology and Paleoanthropology; Brinkman, D.B., Ed.; Springer Science+Business Media: Heidelberg, Germany, 2013; pp. 501–534. [Google Scholar]
- Fediras, S.; Roang, R.; Ziane, N.; Olivier, A.; Bechet, A.; Benyacoub, S. Prevalence of Placobdella costata (Fr. Mûller, 1846) (Hirudinida: Glossiphoniidae) on the European pond turtle (Emys orbicularis) in northeast Algeria. Herpetol. Notes 2017, 10, 3–8. [Google Scholar]
- Trivalairat, P.; Chiangkul, K.; Purivirojkul, W. Parasitism of Placobdelloides siamensis (Oka, 1917) (Glossiphoniidae: Hirudinea) in snail-eating turtles, Malayemys spp., and the effects of host and aquatic environmental factors. Biodivers. Data J. 2020, 8, e57237. [Google Scholar] [CrossRef]
- Chiangkul, K.; Trivalairat, P.; Kunya, K.; Purivirojkul, W. Placobdelloides tridens sp. n., a new species of glossiphoniid leech (Hirudinea: Rhynchobdellida) found feeding on captive Orlitia borneensis in Thailand, and an update to the host distribution of P. siamensis. Syst. Parasitol. 2021, 98, 141–154. [Google Scholar] [CrossRef]
- Guerrero, A.; Ortega, F.; Pérez-García, A. Mark of ectoparasitic activity on a bothremydid (Pleurodira) turtle shell from the Lo Hueco site (Upper Cretaceous, Central Spain). Cretac. Res. 2022, 138, 105297. [Google Scholar] [CrossRef]
- Ernst, C.H.; Ernst, E.M. Ectoparasites associated with neotropical turtles of the genus Callopsis (Testudines, Emydidae, Batagurinae). Biotropica 1977, 9, 139–142. [Google Scholar] [CrossRef]
- Siddall, M.E.; Gaffney, E.S. Observations on the leech Placobdella ornata feeding from bony tissues of turtles. J. Parasitol. 2004, 90, 1186–1188. [Google Scholar] [CrossRef]
- Zonneveld, J.-P.; Bartels, W.S.; Gunnell, G.F.; McHugh, L.P. Borings in early Eocene turtle shell from the Wasatch Formation, South Pass, Wyoming. J. Paleontol. 2015, 89, 802–820. [Google Scholar] [CrossRef]
- Chiangkul, K.; Trivalairat, P.; Purivirojkul, W. Redescription of the Siamese shield leech Placobdelloides siamensis with new host species and geographic range. Parasite 2018, 25, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchgessner, M.; Mitchell, M.A. Chelonians. In Manual of Exotic Pet Practice; Mitchell, M., Tully, T.N., Eds.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009; p. 560. [Google Scholar]
- Girling, S.J.; Raiti, P. BSAVA Manual of Reptiles; No. Ed. 3; British Small Animal Veterinary Association: Quedgeley, UK, 2019; p. 512. [Google Scholar]
- Vella, D. Management of aquatic turtle shell fractures. Lab Anim. 2009, 38, 52–53. [Google Scholar] [CrossRef]
- de Valais, S.; Díaz-Martínez, I.; Citton, P.; Maniel, I.; de la Fuente, M. A predation attempt in a Late Cretaceous pleurodire turtle from Patagonia. Cretac. Res. 2020, 107, 104290. [Google Scholar] [CrossRef]
- Garner, M.M.; Jacobson, E.R. Noninfectious Diseases and Pathology of Reptiles: Color Atlas and Text, Diseases and Pathology of Reptiles; CRC Press: Boca Raton, FL, USA, 2020; Volume 2, p. 512. [Google Scholar]
- Dodd Jr, C.K.; Franz, R.; Johnson, S.A. Shell injuries and anomalies in an insular population of Florida box turtles (Terrapene carolina bauri). Herpetol. Nat. Hist. 1997, 51, 66–72. [Google Scholar]
- Ghiasi, M.S.; Chen, J.; Vaziri, A.; Rodriguez, E.K.; Nazarian, A. Bone fracture healing in mechanobiological modeling: A review of principles and methods. Bone Rep. 2017, 6, 87–100. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero, A.; Ortega, F.; Martín de Jesús, S.; Pérez-García, A. Analysis of the Anomalies in a Middle Eocene Shell of Neochelys (Pleurodira, Podocnemididae) from the Duero Basin (Zamora, Spain). Diversity 2023, 15, 314. https://doi.org/10.3390/d15030314
Guerrero A, Ortega F, Martín de Jesús S, Pérez-García A. Analysis of the Anomalies in a Middle Eocene Shell of Neochelys (Pleurodira, Podocnemididae) from the Duero Basin (Zamora, Spain). Diversity. 2023; 15(3):314. https://doi.org/10.3390/d15030314
Chicago/Turabian StyleGuerrero, Andrea, Francisco Ortega, Santiago Martín de Jesús, and Adán Pérez-García. 2023. "Analysis of the Anomalies in a Middle Eocene Shell of Neochelys (Pleurodira, Podocnemididae) from the Duero Basin (Zamora, Spain)" Diversity 15, no. 3: 314. https://doi.org/10.3390/d15030314
APA StyleGuerrero, A., Ortega, F., Martín de Jesús, S., & Pérez-García, A. (2023). Analysis of the Anomalies in a Middle Eocene Shell of Neochelys (Pleurodira, Podocnemididae) from the Duero Basin (Zamora, Spain). Diversity, 15(3), 314. https://doi.org/10.3390/d15030314