

Cell Plasticity of Marine Mediterranean Diazotrophs to Climate Change Factors and Nutrient Regimes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Strains Tested
2.2. Experimental Culture Conditions
2.2.1. Effects of Variation in pH and Temperature Levels
2.2.2. Effects of Increased CO2 Levels
2.3. Flow Cytometry and Growth Measurement
2.4. Nitrogen Fixation Activity
2.5. Determination of the Alkaline Phosphatase Activity
2.6. Determination of Reactive Oxygen Species
2.7. Statistical Analysis
3. Results
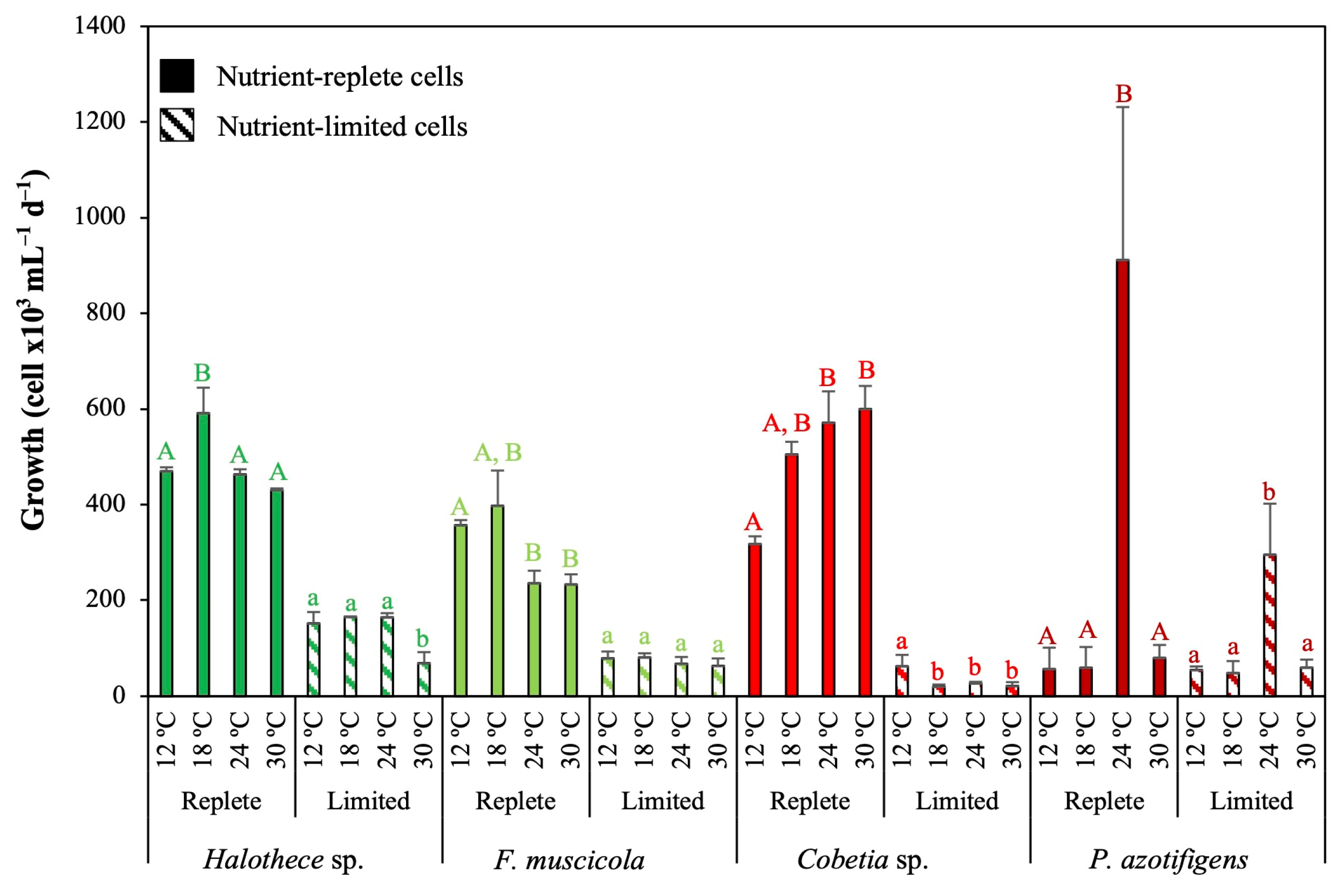
3.1. Impact of Climate Change on Growth Responses
3.1.1. Effect of pH and Temperature
3.1.2. Effect of CO2
3.2. Effect of Climate Change on N2 Fixation Rates
3.2.1. Effect of pH and Temperature
3.2.2. Effect of CO2
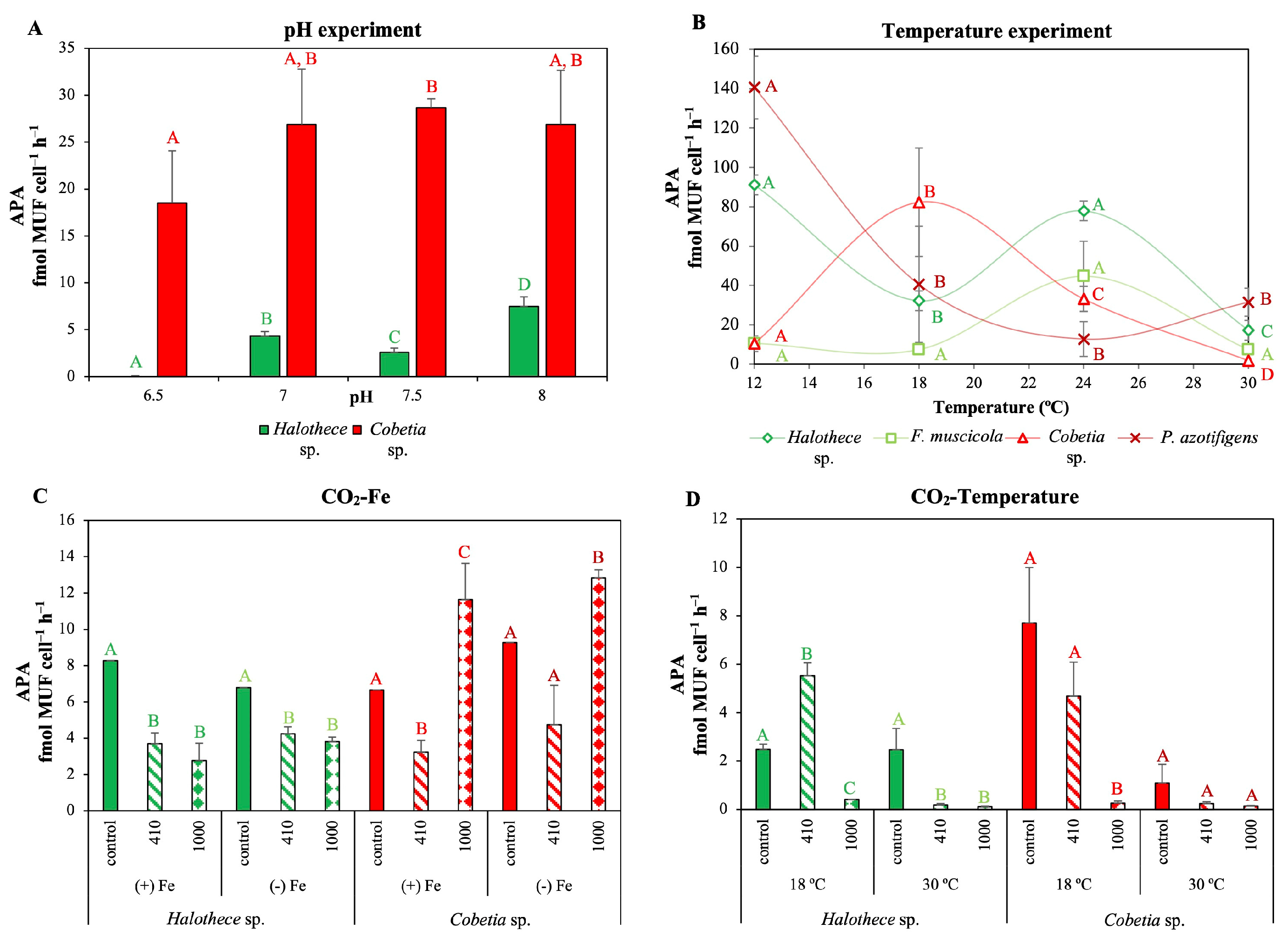
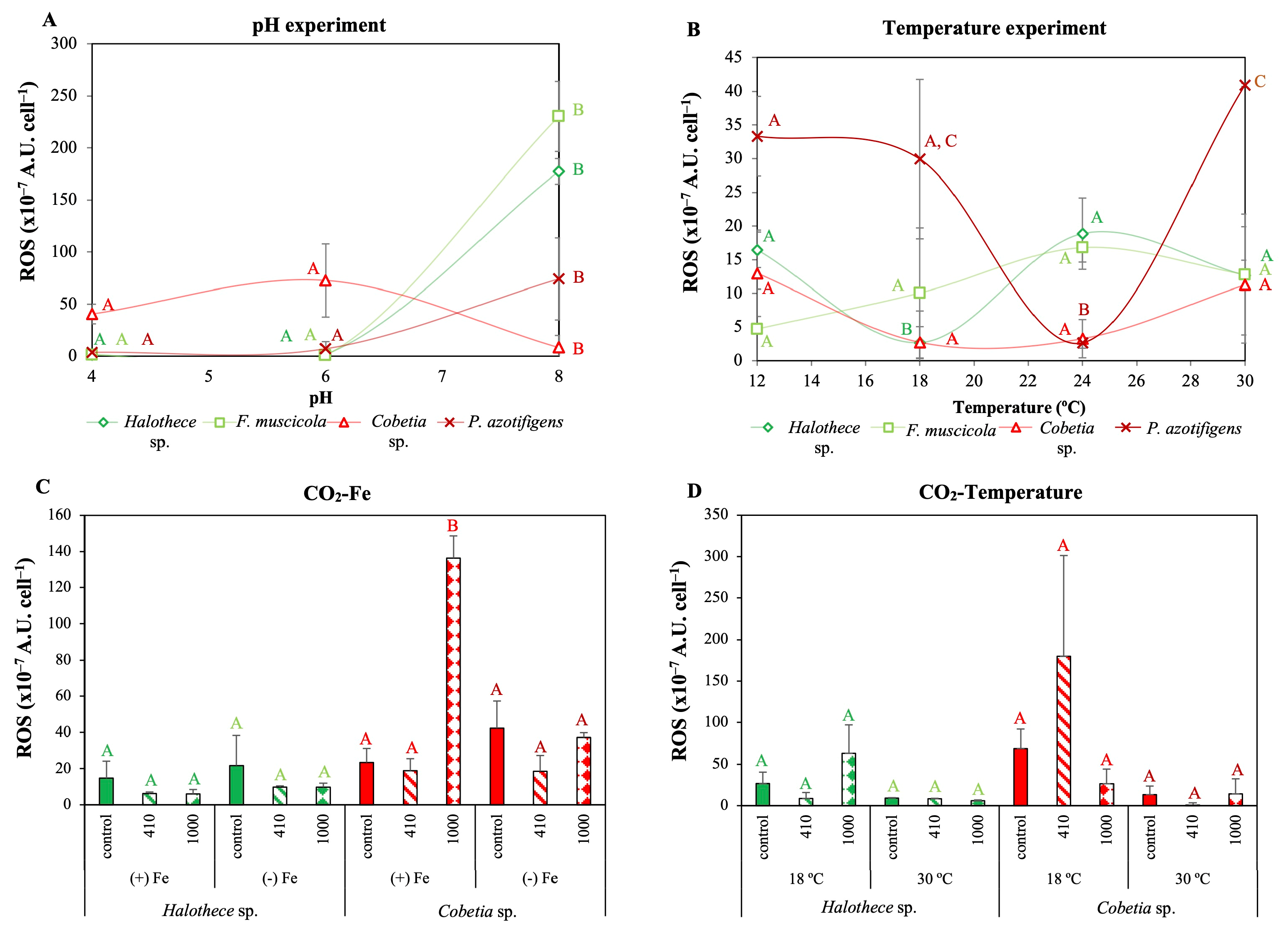
3.3. Plasticity of Diazotrophs against Climate Change: Changes in P-Acquisition Mechanisms and Oxidative Stress
3.3.1. Changes in P-Acquisition Mechanisms
3.3.2. Changes in ROS
3.3.3. APA and ROS as Molecular Biomarkers
4. Discussion
4.1. Dependence of Nutrient Status of N2 Fixers in Their Growth Responses to Climate Change Factors
4.2. Climate Change Effect on N2 Fixation
4.3. P-Acquisition Mechanisms and Oxidative Stress as Biomarkers for Predicting Microbial Perturbations Associated with Climate Change
4.3.1. P-Acquisition Mechanisms
4.3.2. Oxidative Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Etheridge, D.M.; Steele, L.P.; Langenfelds, R.L.; Francey, R.J.; Barnola, J.-M.; Morgan, V.I. Natural and anthropogenic changes in atmospheric CO2 over the last 1000 years from air in Antarctic ice and firn. J. Geophys. Res. Atmos. 1996, 101, 4115–4128. [Google Scholar] [CrossRef] [Green Version]
- Raupach, M.R.; Marland, G.; Ciais, P.; Le Quéré, C.; Canadell, J.G.; Klepper, G.; Field, C.B. Global and regional drivers of accelerating CO2 emissions. Proc. Natl. Acad. Sci. USA 2007, 104, 10288–10293. [Google Scholar] [CrossRef] [Green Version]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Hauck, J.; Olsen, A.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; et al. Global Carbon Budget 2020. Earth Syst. Sci. Data 2020, 12, 3269–3340. [Google Scholar] [CrossRef]
- Jiang, L.Q.; Carter, B.R.; Feely, R.A.; Lauvset, S.K.; Olsen, A. Surface ocean pH and buffer capacity: Past, present and future. Sci. Rep. 2019, 9, 18624. [Google Scholar] [CrossRef] [Green Version]
- Sarmento, H.; Montoya, J.M.; Vázquez-Domínguez, E.; Vaque, D.; Gasol, J.M. Warming effects on marine microbial food web processes: How far can we go when it comes to predictions? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2137–2149. [Google Scholar] [CrossRef] [Green Version]
- Berge, T.; Daugbjerg, N.; Andersen, B.B.; Hansen, P.J. Effect of lowered pH on marine phytoplankton growth rates. Mar. Ecol. Prog. Ser. 2010, 416, 79–91. [Google Scholar] [CrossRef]
- Nielsen, L.T.; Hallegraeff, G.M.; Wright, S.W.; Hansen, P.J. Effects of experimental seawater acidification on an estuarine plankton community. Aquat. Microb. Ecol. 2012, 65, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.E.; Newbold, L.K.; Whiteley, A.S.; van der Gast, C.J. Marine bacterial communities are resistant to elevated carbon dioxide levels. Environ. Microbiol. Rep. 2014, 6, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Lee, K.; Shin, K.; Kang, J.H.; Lee, H.W.; Kim, M.; Jang, P.G.; JanG, M.C. The effect of seawater CO2 concentration on growth of a natural phytoplankton assemblage in a controlled mesocosm experiment. Limnol. Oceanogr. 2006, 51, 1629–1636. [Google Scholar] [CrossRef] [Green Version]
- Krause, E.; Wichels, A.; Giménez, L.; Lunau, M.; Schilhabel, M.B.; Gerdts, G. Small changes in pH have direct effects on marine bacterial community composition: A microcosm approach. PLoS ONE 2012, 7, e47035. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Capone, D.G. Changing perspectives in marine nitrogen fixation. Science 2020, 368, eaay9514. [Google Scholar] [CrossRef]
- Shi, D.; Kranz, S.A.; Kim, J.M.; Morel, F.M.M. Ocean acidification slows nitrogen fixation and growth in the dominant diazotroph Trichodesmium under low-iron conditions. Proc. Natl. Acad. Sci. USA 2012, 109, E3094–E3100. [Google Scholar] [CrossRef] [Green Version]
- Breitbarth, E.; Oschlies, A.; LaRoche, J. Physiological constraints on the global distribution of Trichodesmium—Effect of temperature on diazotrophy. Biogeosciences 2007, 4, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Eichner, M.; Kranz, S.A.; Rost, B. Combined effects of different CO2 levels and N sources on the diazotrophic cyanobacterium Trichodesmium. Physiol. Plant. 2014, 152, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Boatman, T.G.; Lawson, T.; Geider, R.J. A key marine diazotroph in a changing ocean: The interacting effects of temperature, CO2 and light on the growth of Trichodesmium erythraeum IMS101. PLoS ONE 2017, 12, e0168796. [Google Scholar] [CrossRef] [Green Version]
- Eichner, M.; Rost, B.; Kranz, S.A. Diversity of ocean acidification effects on marine N2 fixers. J. Exp. Mar. Biol. Ecol. 2014, 457, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Riemann, L.; Farnelid, H.; Steward, G.F. Nitrogenase genes in non-cyanobacterial plankton: Prevalence, diversity and regulation in marine waters. Aquat. Microb. Ecol. 2010, 61, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Moisander, P.H.; Benavides, M.; Bonnet, S.; Berman-Frank, I.; White, A.E.; Riemann, L. Chasing after non-cyanobacterial nitrogen fixation in marine pelagic environments. Front. Microbiol. 2017, 8, 1736. [Google Scholar] [CrossRef]
- Chakraborty, S.; Andersen, K.H.; Follows, M.J.; Riemann, L. Quantifying nitrogen fixation by heterotrophic bacteria in sinking marine particles. Nat. Commun. 2021, 12, 4085. [Google Scholar] [CrossRef]
- Santruckova, H.; Bird, M.I.; Elhottová, D.; Novák, J.; Picek, T.; Simek, M.; Tykva, R. Heterotrophic fixation of CO2 in soil. Microb. Ecol. 2005, 49, 218–225. [Google Scholar] [CrossRef]
- Jiang, H.; Fu, F.; Rivero-Calle, S.; Levine, N.M.; Sañudo-Wilhelmy, S.A.; Qu, P.; Wang, X.; Pinedo-Gonzalez, P.; Zhu, Z.; Hutchins, D.A. Ocean warming alleviates iron limitation of marine nitrogen fixation. Nat. Clim. Chang. 2018, 8, 709–712. [Google Scholar] [CrossRef]
- Boyd, P.W.; Doney, S.C. Modelling regional responses by marine pelagic ecosystems to global climate change. Geophys. Res. Lett. 2002, 29, 53-1–53-4. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.; Yu, E.; Garcia, N.; Gale, J.; Luo, Y.; Webb, E.; Hutchins, D.A. Differing responses of marine N2 fixers to warming and consequences for future diazotroph community structure. Aquat. Microb. Ecol. 2014, 72, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Lagus, A.; Suomela, J.; Weithoff, G.; Heikkilä, K.; Helminen, H.; Sipura, J. Species-specific differences in phytoplankton responses to N and P enrichments and the N:P ratio in the Archipelago Sea, northern Baltic Sea. J. Plankton Res. 2004, 26, 779–798. [Google Scholar] [CrossRef] [Green Version]
- Striebel, M.; Schabhüttl, S.; Hodapp, D.; Hingsamer, P.; Hillebrand, H. Phytoplankton responses to temperature increases are constrained by abiotic conditions and community composition. Oecologia 2016, 182, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Thackeray, S.J.; Jones, I.D.; Maberly, S.C. Long-term change in the phenology of spring phytoplankton: Species-specific responses to nutrient enrichment and climatic change. J. Ecol. 2008, 96, 523–535. [Google Scholar] [CrossRef]
- Wrightson, L.; Tagliabue, A. Quantifying the impact of climate change on marine diazotrophy: Insights from earth system models. Front. Mar. Sci. 2020, 7, 635. [Google Scholar] [CrossRef]
- Ward, B.A.; Dutkiewicz, S.; Moore, C.M.; Follows, M.J. Iron, phosphorus, and nitrogen supply ratios define the biogeography of nitrogen fixation. Limnol. Oceanogr. 2013, 58, 2059–2075. [Google Scholar] [CrossRef]
- Falkowski, P.G. Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 1997, 387, 272–275. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, B.M.; Lukoyanov, D.; Yang, Z.Y.; Dean, D.R.; Seefeldt, L.C. Mechanism of nitrogen fixation by nitrogenase: The next Stage. Chem. Rev. 2014, 114, 4041–4062. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Ferriol, P.; Cryer, C.; Alcon, E.; Busquets, A.; Sintes, E.; Vidal, C.; Moyà, G. Significant nitrogen fixation activity associated with the phyllosphere of Mediterranean seagrass Posidonia oceanica: First report. Mar. Ecol. Prog. Ser. 2016, 551, 53–62. [Google Scholar] [CrossRef]
- García-Márquez, M.G.; Fernández-Juárez, V.; Rodríguez-Castañeda, J.C.; Agawin, N.S.R. Response of Posidonia oceanica (L.) Delile and Its Associated N2 Fixers to Different Combinations of Temperature and Light Levels. Front. Mar. Sci. 2022, 8, 1966. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Gil Atorrasagasti, M.G.; Frank Comas, A.; Fernández-Juárez, V.; López-Alforja, X.; Hendriks, I. EResponse of the seagrass Posidonia oceanica and its associated N2 fixers to high business-as-usual climate change scenario in winter. Limnol. Oceanogr. 2021, 66, 2346–2359. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Ferriol, P.; Sintes, E.; Moyà, G. Temporal and spatial variability of in situ nitrogen fixation activities associated with the Mediterranean seagrass Posidonia oceanica meadows. Limnol. Oceanogr. 2017, 62, 2575–2592. [Google Scholar] [CrossRef]
- Fernández-Juárez, V.; Bennasar-Figueras, A.; Sureda-Gomila, A.; Ramis-Munar, G.; Agawin, N.S.R. Differential Effects of Varying Concentrations of Phosphorus, Iron, and Nitrogen in N2 Fixing Cyanobacteria. Front. Microbiol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Fernández-Juárez, V.; Bennasar-Figueras, A.; Tovar-Sanchez, A.; Agawin, N.S. The role of iron in the P-acquisition mechanisms of the unicellular N2 -fixing cyanobacteria Halothece sp., found in association with the Mediterranean seagrass Posidonia oceanica. Front. Microbiol. 2019, 10, 1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Juárez, V.; López-alforja, X.; Frank-comas, A.; Echeveste, P. “The good, the bad and the double-sword ” effects of microplastics and their organic additives in marine bacteria. Front. Microbiol. 2021, 11, 581118. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Juárez, V.; Jaén-Luchoro, D.; Brito-Echeverría, J.; Agawin, N.S.; Bennasar-Figueras, A.; Echeveste, P. Everything Is Everywhere: Physiological Responses of the Mediterranean Sea and Eastern Pacific Ocean Epiphyte Cobetia Sp. to Varying Nutrient Concentration. Microb. Ecol. 2021, 83, 296–313. [Google Scholar] [CrossRef]
- Moßhammer, M.; Brodersen, K.E.; Kühl, M.; Koren, K. Nanoparticle- and microparticle-based luminescence imaging of chemical species and temperature in aquatic systems: A review. Microchim. Acta 2019, 186, 20–23. [Google Scholar] [CrossRef]
- Knapp, A.N. The sensitivity of marine N2 fixation to dissolved inorganic nitrogen. Front. Microbiol. 2012, 3, 374. [Google Scholar] [CrossRef] [Green Version]
- Agawin, N.S.R.; Benavides, M.; Busquets, A.; Ferriol, P.; Stal, L.J.; Arístegui, J. Dominance of unicellular cyanobacteria in the diazotrophic community in the Atlantic Ocean. Limnol. Oceanogr. 2014, 59, 623–637. [Google Scholar] [CrossRef]
- Stal, L.J. Acetylene reduction technique for assay of nitrogenase. Methods Enzymol. 1988, 167, 474–484. [Google Scholar]
- Jensen, B.B.; Cox, R.P. Direct measurements of steady-state kinetics of cyanobacterial N2uptake by membrane-leak mass spectrometry and comparisons between nitrogen fixation and acetylene reduction. Appl. Environ. Microbiol. 1983, 45, 1331–1337. [Google Scholar] [CrossRef] [Green Version]
- Sanudo-Wilhelmy, S.A.; Kustka, A.B.; Gobler, C.J.; Hutchins, D.A.; Yang, M.; Lwiza, K.; Burns, J.; Capone, D.G.; Raven, J.A.; Carpenter, J.E. Phosphorus limitation of nitrogen fixation by Trichodesmium in the central Atlantic Ocean. Nature 2001, 411, 66–69. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Cullen, J.T.; Shaked, Y.; Sherrell, R.M.; Falkowski, P.G. Iron availability, cellular iron quotas, and nitrogen fixation in Trichodesmium. Limnol. Oceanogr. 2001, 46, 1249–1260. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Quigg, A.; Finkel, Z.V.; Irwin, A.J.; Haramaty, L. Nitrogen-fixation strategies and Fe requirements in cyanobacteria. Limnol. Oceanogr. 2007, 52, 2260–2269. [Google Scholar] [CrossRef]
- Schubert, E.; Mengel, K.; Schubert, S. Soil pH and calcium effect on nitrogen fixation and growth of board bean. Agron. J. 1990, 82, 969–972. [Google Scholar] [CrossRef]
- Pham, D.N.; Burgess, B.K. Nitrogenase reactivity: Effects of pH on substrate reduction and CO Inhibition. Biochemistry 1993, 32, 13725–13731. [Google Scholar] [CrossRef]
- Fernandez Valiente, E.; Leganes, F. Regulatory effect of pH and incident irradiance on the levels of nitrogenase activity in the cyanobacterium nostoc UAM 205. J. Plant Physiol. 1990, 135, 623–627. [Google Scholar] [CrossRef]
- Millero, F.J.; Woosey, R.; Ditrolio BWater, J. effect of Ocean acidification on the Speciation of Metals in Seawater. Oceanography 2009, 22, 72–85. [Google Scholar] [CrossRef]
- Shi, D.; Xu, Y.; Hopkinson, B.M.; Morel, F.M.M. Effect of ocean acidification on iron availability to marine phytoplankton. Science 2010, 327, 676–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Chen, Y. Effects of elevated carbon dioxide on environmental microbes and its mechanisms: A review. Sci. Total Environ. 2019, 655, 865–879. [Google Scholar] [CrossRef]
- Waditee, R.; Hibino, T.; Tanaka, Y.; Nakamura, T.; Incharoensakdi, A.; Takabe, T. Halotolerant Cyanobacterium Aphanothece halophytica Contains an Na+/H+ Antiporter, Homologous to Eukaryotic Ones, with Novel Ion Specificity Affected by C-terminal Tail. J. Biol. Chem. 2001, 276, 36931–36938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeroy, L.R.; Wiebe, W.J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Schabhüttl, S.; Hingsamer, P.; Weigelhofer, G.; Hein, T.; Weigert, A.; Striebel, M. Temperature and species richness effects in phytoplankton communities. Temperature and species richness effects in phytoplankton communities. Oecologia 2013, 171, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Marañón, E.; Lorenzo, M.P.; Cermeño, P.; Mouriño-Carballido, B. Nutrient limitation suppresses the temperature dependence of phytoplankton metabolic rates. ISME J. 2018, 12, 1836–1845. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.K.; Aranguren-Gassis, M.; Kremer, C.T.; Gould, M.R.; Anderson, K.; Klausmeier, C.A.; Litchman, E. Temperature–nutrient interactions exacerbate sensitivity to warming in phytoplankton. Glob. Chang. Biol. 2017, 23, 3269–3280. [Google Scholar] [CrossRef]
- Akpor, O.; Olaolu, T.D.; Okolie, E.C. The effect of temperature on nitrate and phosphate uptake from synthetic wastewater by selected bacteria species. Br. Microbiol. Res. J. 2014, 4, 328–342. [Google Scholar] [CrossRef]
- Huete-Stauffer, T.M.; Arandia-Gorostidi, N.; Alonso-Sáez, L.; Morán, X.A. Experimental warming decreases the average size and nucleic acid content of marine bacterial communities. Front. Microbiol. 2016, 7, 730. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Carroll, A.L.; Atsumi, S. Carbon recycling by cyanobacteria: Improving CO2 fixation through chemical production. FEMS Microbiol. Lett. 2017, 364, fnx165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcelos e Ramos, J.; Biswas, H.; Schulz, K.G.; LaRoche, J.; Riebesell, U. Effect of rising atmospheric carbon dioxide on the marine nitrogen fixer Trichodesmium. Glob. Biogeochem. Cycles 2007, 21, GB2028. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.X.; Mulholland, M.R.; Garcia, N.S.; Beck, A.; Bernhardt, P.W.; Warner, M.E.; Sañudo-Wilhelmy, S.A.; Hutchins, D.A. Interactions between changing pCO2, N2 fixation, and Fe limitation in the marine unicellular cyanobacterium Crocosphaera. Limnol. Oceanogr. 2008, 53, 2472–2484. [Google Scholar] [CrossRef] [Green Version]
- Kranz, S.A.; Sültemeyer, D.; Richter, K.U.; Rost, B. Carbon acquisition by Trichodesmium: The effect of pCO2 and diurnal changes. Limnol. Oceanogr. 2009, 54, 548–559. [Google Scholar] [CrossRef]
- Garcia, N.S.; Fu, F.X.; Breene, C.L.; Bernhardt, P.W.; Mulholland, M.R.; Sohm, J.A.; Hutchins, D. AInteractive effects of irradiance and CO2 on CO2 fixation and N2 fixation in the diazotroph Trichodesmium erythraeum (cyanobacteria). J. Phycol. 2011, 47, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.S.; Fu, F.X.; Breene, C.L.; Yu, E.K.; Bernhardt, P.W.; Mulholland, M.R.; Hutchins, D. A Combined effects of CO2 and light on large and small isolates of the unicellular N2 fixing cyanobacterium Crocosphaera watsonii from the western tropical Atlantic Ocean. Eur. J. Phycol. 2013, 48, 128–139. [Google Scholar] [CrossRef]
- Garcia, N.S.; Fu, F.X.; Hutchins, D.A. Colimitation of the unicellular photosynthetic diazotroph Crocosphaera watsonii by phosphorus, light, and carbon dioxide. Limnol. Oceanogr. 2013, 58, 1501–1512. [Google Scholar] [CrossRef]
- Levitan, O.; Rosenberg, G.; Setlik, I.; Grigel, J.; Klepetar, J.; Prasil, O.; Berkman-Frank, I. Elevated CO2 enhances nitrogen fixation and growth in the marine cyanobacterium Trichodesmium. Glob. Chang. Biol. 2007, 13, 531–538. [Google Scholar] [CrossRef]
- Wannicke, N.; Endres, S.; Engel, A.; Grossart, H.P.; Nausch, M.; Unger, J.; Voss, M. Response of Nodularia spumigena to pCO2—Part 1: Growth, production and nitrogen cycling. Biogeosciences 2012, 9, 2973–2988. [Google Scholar] [CrossRef] [Green Version]
- Teira, E.; Fernández, A.; Álvarez-Salgado, X.A.; García-Martín, E.; Serret, P.; Sobrino, C. Response of two marine bacterial isolates to high CO2 concentration. Mar. Ecol. Prog. Ser. 2012, 453, 27–36. [Google Scholar] [CrossRef] [Green Version]
- James, A.K.; Kelly, L.W.; Nelson, C.E.; Wilbanks, E.G.; Carlson, C.A. Elevated pCO2 alters marine heterotrophic bacterial community composition and metabolic potential in response to a pulse of phytoplankton organic matter. Environ. Microbiol. 2019, 21, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Grossart, H.P.; Allgaier, M.; Passow, U.; Riebesell, U. Testing the effect of CO2 concentration on the dynamics of marine heterotrophic bacterioplankton. Limnol. Oceanogr. 2006, 51, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bristow, L.A.; Mohr, W.; Ahmerkamp, S.; Kuypers, M.M.M. Nutrients that limit growth in the ocean. Curr. Biol. 2017, 27, R474–R478. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.J.; Kirkegaard, R.; Szul, M.J.; Johnson, Z.I.; Zinser, E.R. Facilitation of robust growth of Prochlorococcus colonies and dilute liquid cultures by “helper” heterotrophic bacteria. Appl. Environ. Microbiol. 2008, 74, 4530–4534. [Google Scholar] [CrossRef] [Green Version]
- Kranz, S.A.; Eichner, M.; Rost, B. Interactions between CCM and N2 fixation in Trichodesmium. Photosynth. Res. 2010, 109, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Spona-Friedl, M.; Braun, A.; Huber, C.; Eisenreich, W.; Griebler, C.; Kappler, A.; Elsner, M. Substrate-dependent CO2 fixation in heterotrophic bacteria revealed by stable isotope labelling. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Liu, W.; Gao, K. Effects of temperature, pH, and UV radiation on alkaline phosphatase activity in the terrestrial cyanobacterium Nostoc flagelliforme. J. Appl. Phycol. 2013, 25, 1031–1038. [Google Scholar] [CrossRef]
- Yu Plisova, E.; Balabanova, L.A.; Ivanova, E.P.; Kozhemyako, V.B.; Mikhailov, V.V.; Agafonova, E.V.; Rasskazov, V.A. A highly active alkaline phosphatase from the marine bacterium Cobetia. Mar. Biotechnol. 2005, 7, 173–178. [Google Scholar] [CrossRef]
- Noskova, Y.; Likhatskaya, G.; Terentieva, N.; Son, O.; Tekutyeva, L.; Balabanova, L. A novel alkaline phosphatase/phosphodiesterase, CamPhoD, from marine bacterium Cobetia amphilecti KMM 296. Mar. Drugs 2019, 17, 657. [Google Scholar] [CrossRef] [Green Version]
- Cerozi B da, S.; Fitzsimmons, K. The effect of pH on phosphorus availability and speciation in an aquaponics nutrient solution. Bioresour. Technol. 2016, 219, 778–781. [Google Scholar] [CrossRef] [Green Version]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant production and characterization of a highly active alkaline phosphatase from marine bacterium Cobetia marina. Mar. Biotechnol. 2015, 17, 130–143. [Google Scholar] [CrossRef]
- Hernández, I.; Niell, F.X.; Whitton, B.A. Phosphatase activity of benthic marine algae. An overview. J. Appl. Phycol. 2002, 14, 475–487. [Google Scholar] [CrossRef]
- Ferrat, L.; Pergent-Martini, C.; Roméo, M. Assessment of the use of biomarkers in aquatic plants for the evaluation of environmental quality: Application to seagrasses. Aquat. Toxicol. 2003, 65, 187–204. [Google Scholar] [CrossRef]
- Benedetti, M.; Giuliani, M.E.; Regoli, F. Oxidative metabolism of chemical pollutants in marine organisms: Molecular and biochemical biomarkers in environmental toxicology. Ann. N. Y. Acad. Sci. 2015, 1340, 8–19. [Google Scholar] [CrossRef]
- Litchman, E.; Edwards, K.F.; Klausmeier, C.A.; Thomas, M.K. Phytoplankton niches, traits and eco-evolutionary responses to global environmental change. Mar. Ecol. Prog. Ser. 2012, 470, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Au, D.W.T.; Anderson, D.M.; Lam, P.K.S.; Wu, R.S.S. Effects of nutrients, salinity, pH and light:dark cycle on the production of reactive oxygen species in the alga Chattonella marina. J. Exp. Mar. Biol. Ecol. 2007, 346, 76–86. [Google Scholar] [CrossRef]
- Twiner, M.J. Possible physiological mechanisms for production of hydrogen peroxide by the ichthyotoxic flagellate Heterosigma akashiwo. J. Plankton Res. 2000, 22, 1961–1975. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, M. Mechanism of bacterial adaptation to low temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef]
- Diaz, J.M.; Plummer, S. Production of extracellular reactive oxygen species by phytoplankton: Past and future directions. J. Plankton Res. 2018, 40, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Hennon, G.M.; Morris, J.J.; Haley, S.T.; Zinser, E.R.; Durrant, A.R.; Entwistle, E.; Dokland, T.; Dyhrman, S.T. The impact of elevated CO2 on Prochlorococcus and microbial interactions with a ‘helper’ bacterium Alteromonas. ISME J. 2017, 12, 520–531. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Juárez, V.; Zech, E.H.; Pol-Pol, E.; Agawin, N.S.R. Cell Plasticity of Marine Mediterranean Diazotrophs to Climate Change Factors and Nutrient Regimes. Diversity 2023, 15, 316. https://doi.org/10.3390/d15030316
Fernández-Juárez V, Zech EH, Pol-Pol E, Agawin NSR. Cell Plasticity of Marine Mediterranean Diazotrophs to Climate Change Factors and Nutrient Regimes. Diversity. 2023; 15(3):316. https://doi.org/10.3390/d15030316
Chicago/Turabian StyleFernández-Juárez, Víctor, Elisa H. Zech, Elisabet Pol-Pol, and Nona S. R. Agawin. 2023. "Cell Plasticity of Marine Mediterranean Diazotrophs to Climate Change Factors and Nutrient Regimes" Diversity 15, no. 3: 316. https://doi.org/10.3390/d15030316
APA StyleFernández-Juárez, V., Zech, E. H., Pol-Pol, E., & Agawin, N. S. R. (2023). Cell Plasticity of Marine Mediterranean Diazotrophs to Climate Change Factors and Nutrient Regimes. Diversity, 15(3), 316. https://doi.org/10.3390/d15030316