
Patterns of European Ant Communities Reveal a Functionally Coherent Holarctic Fauna



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling
2.3. Analysis
2.4. Functional Group Classification and Re-Analysis of Previous Studies
3. Results
3.1. The Fauna
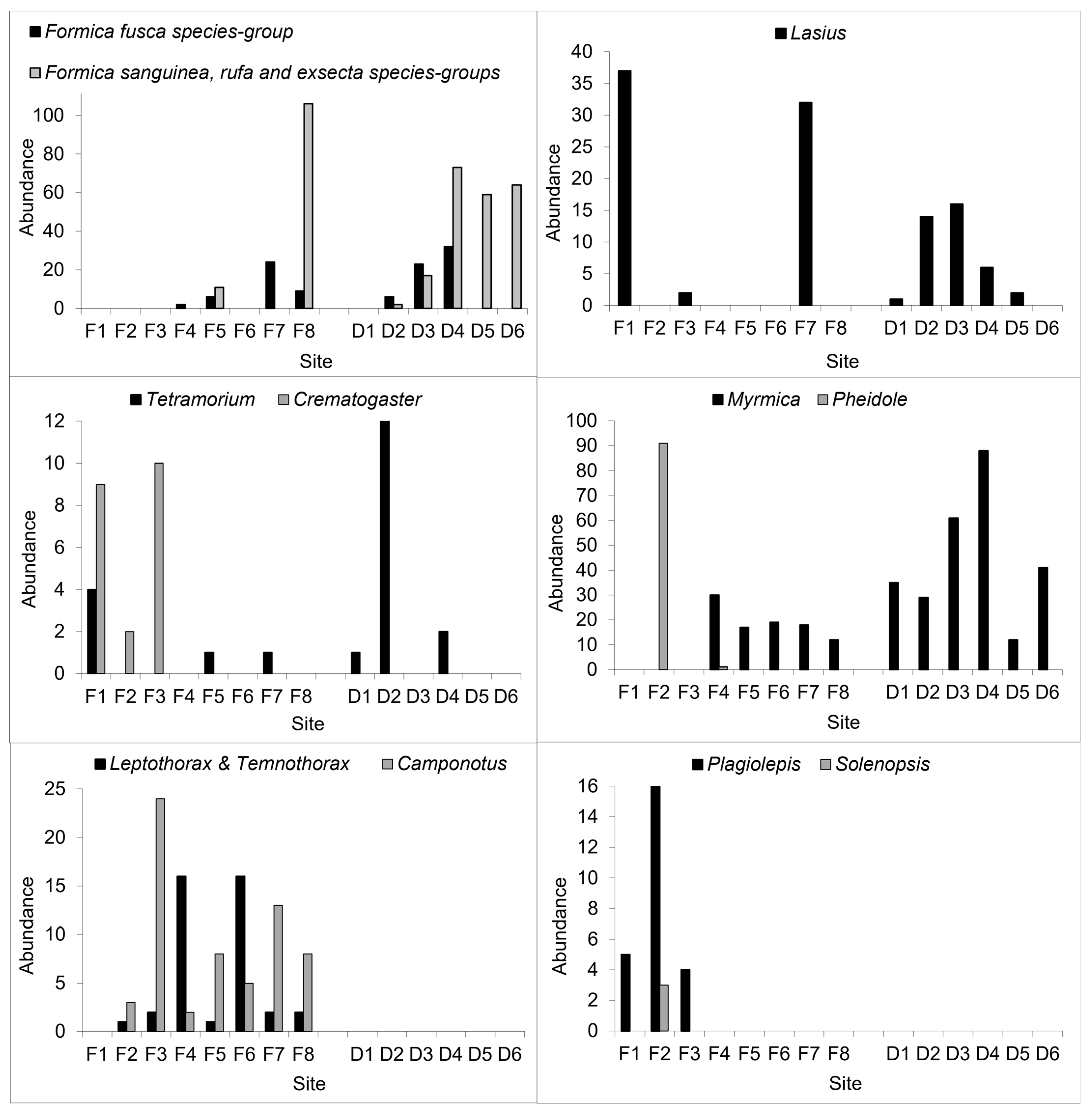
3.2. Geographic Patterns
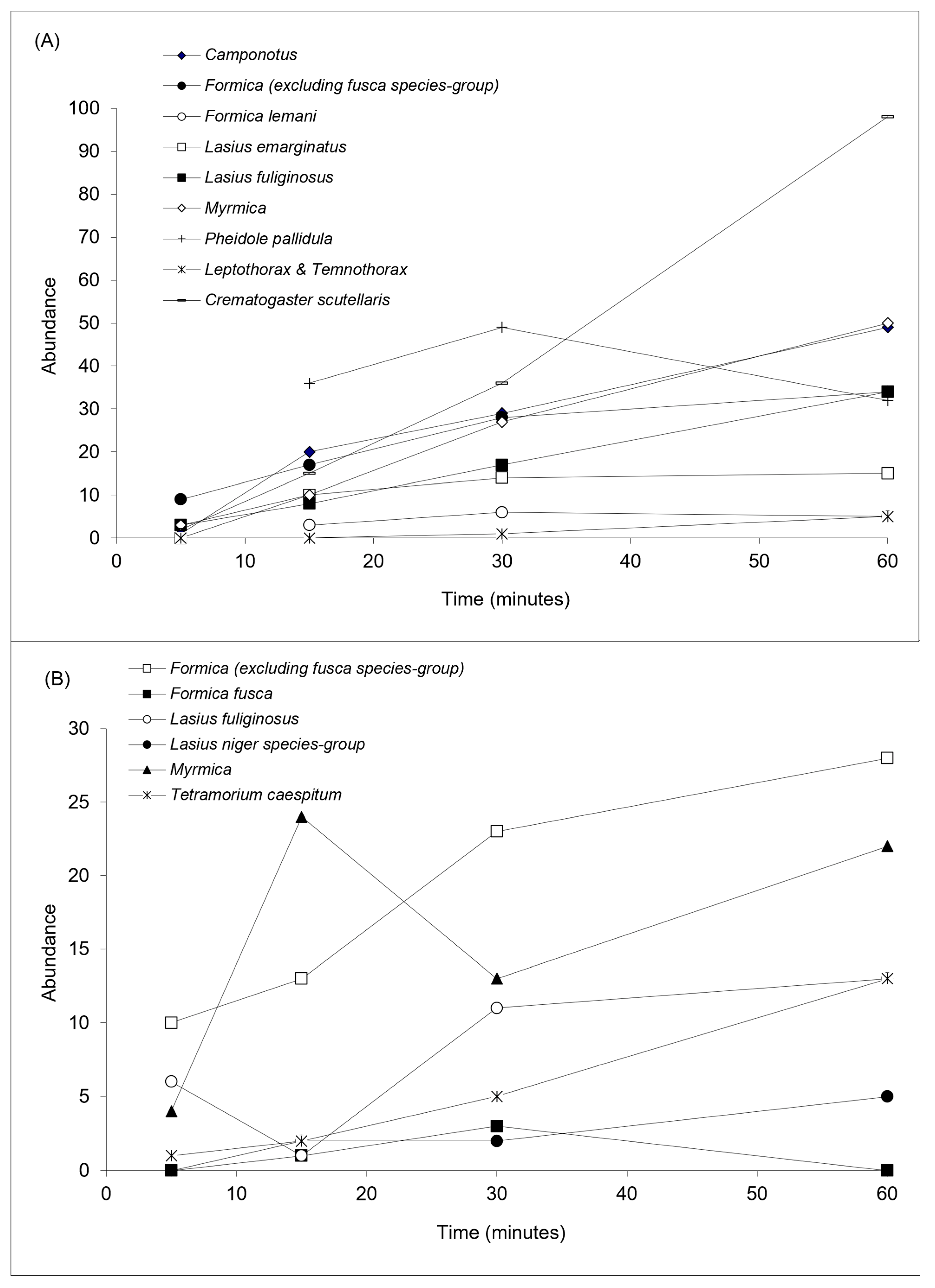
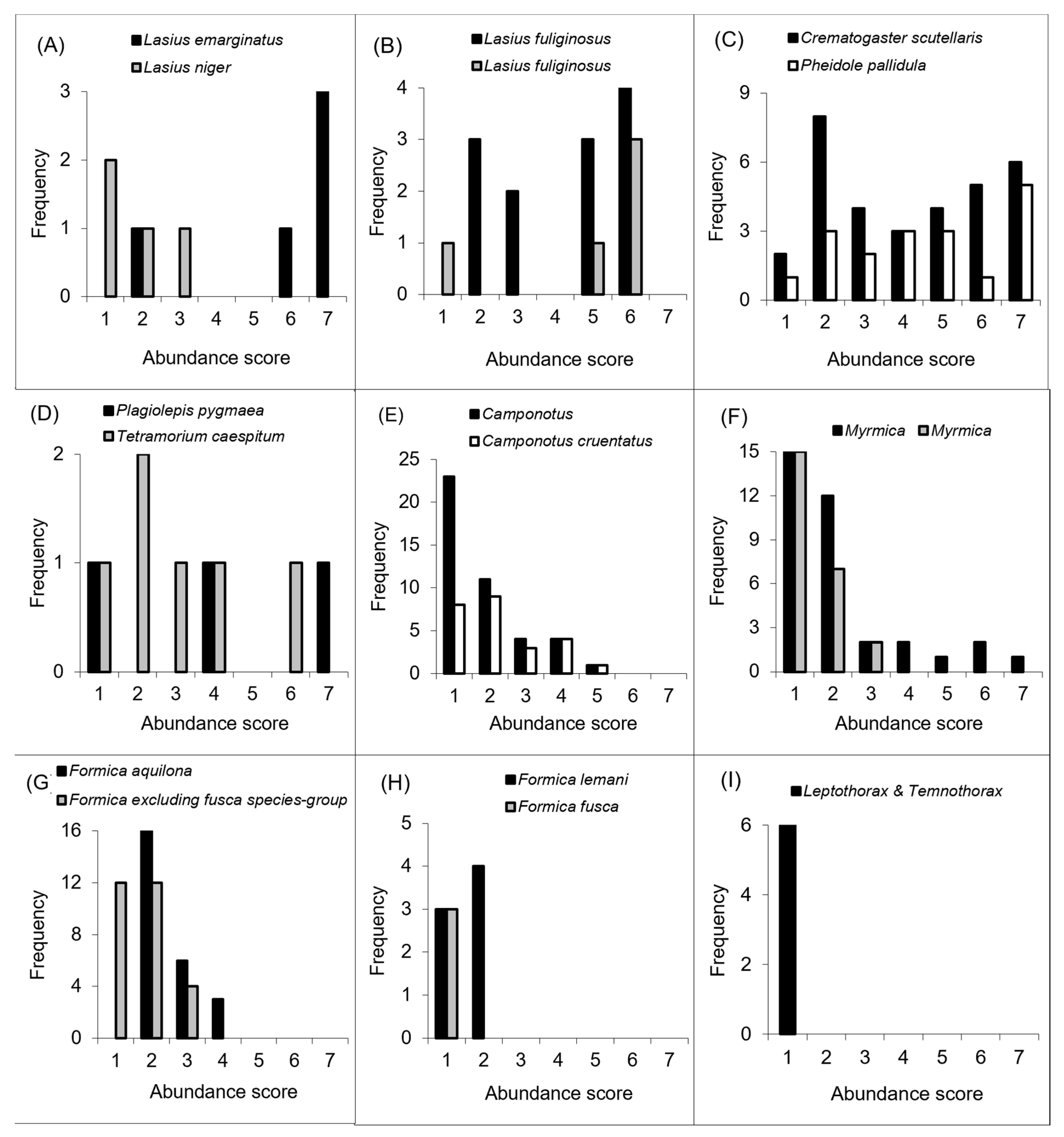
3.3. Behavioural Dominance
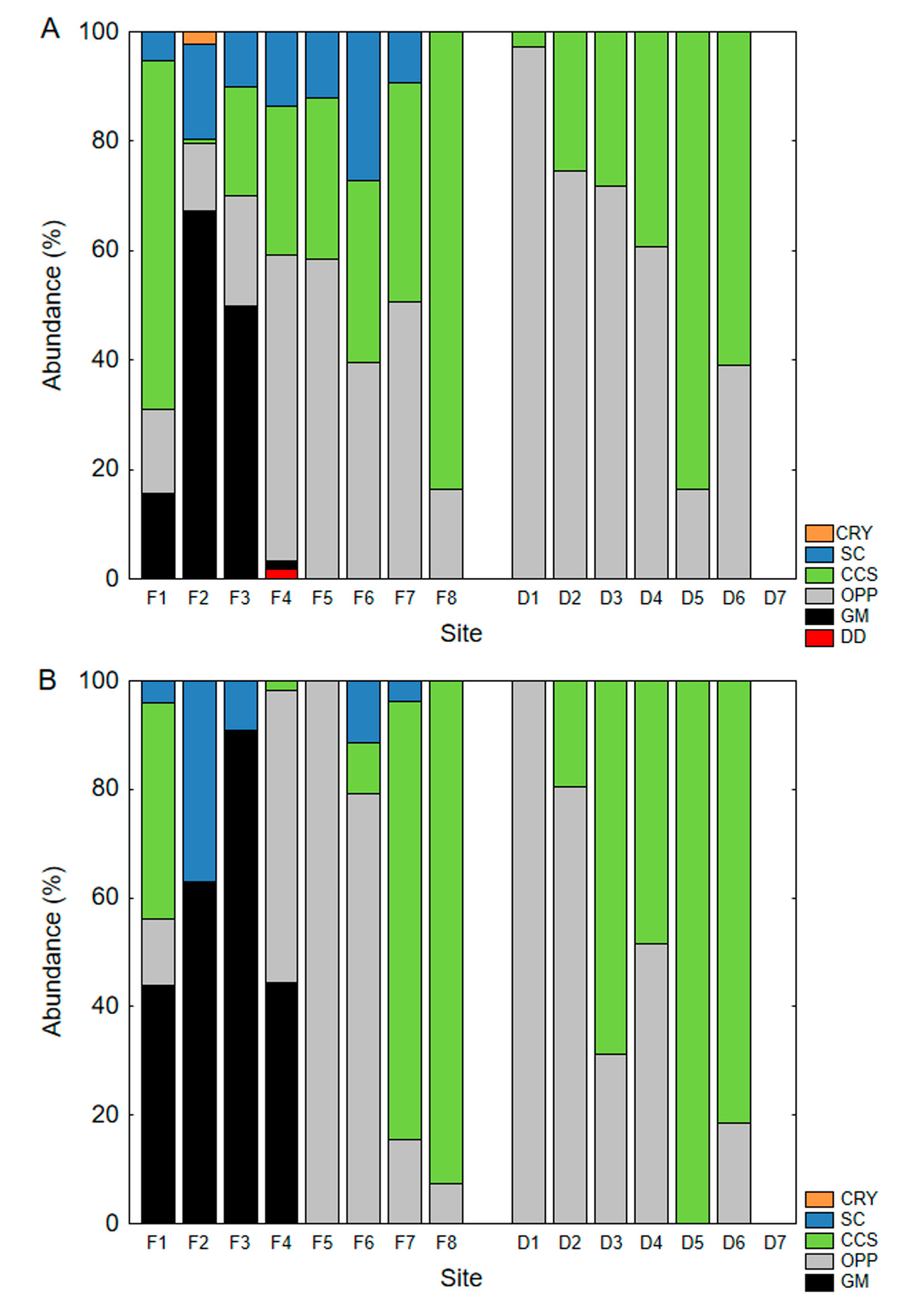
3.4. Functional Group Classification
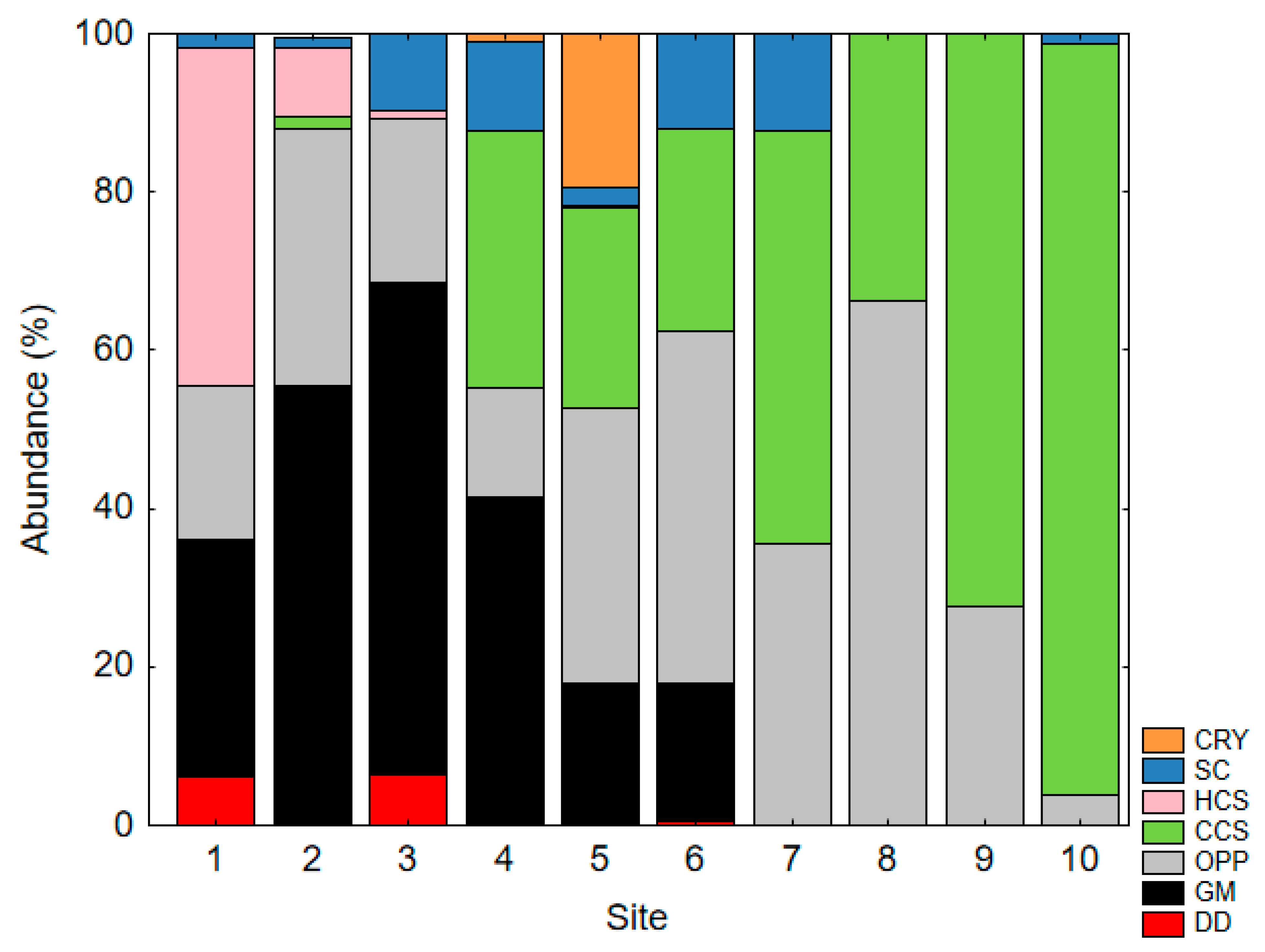
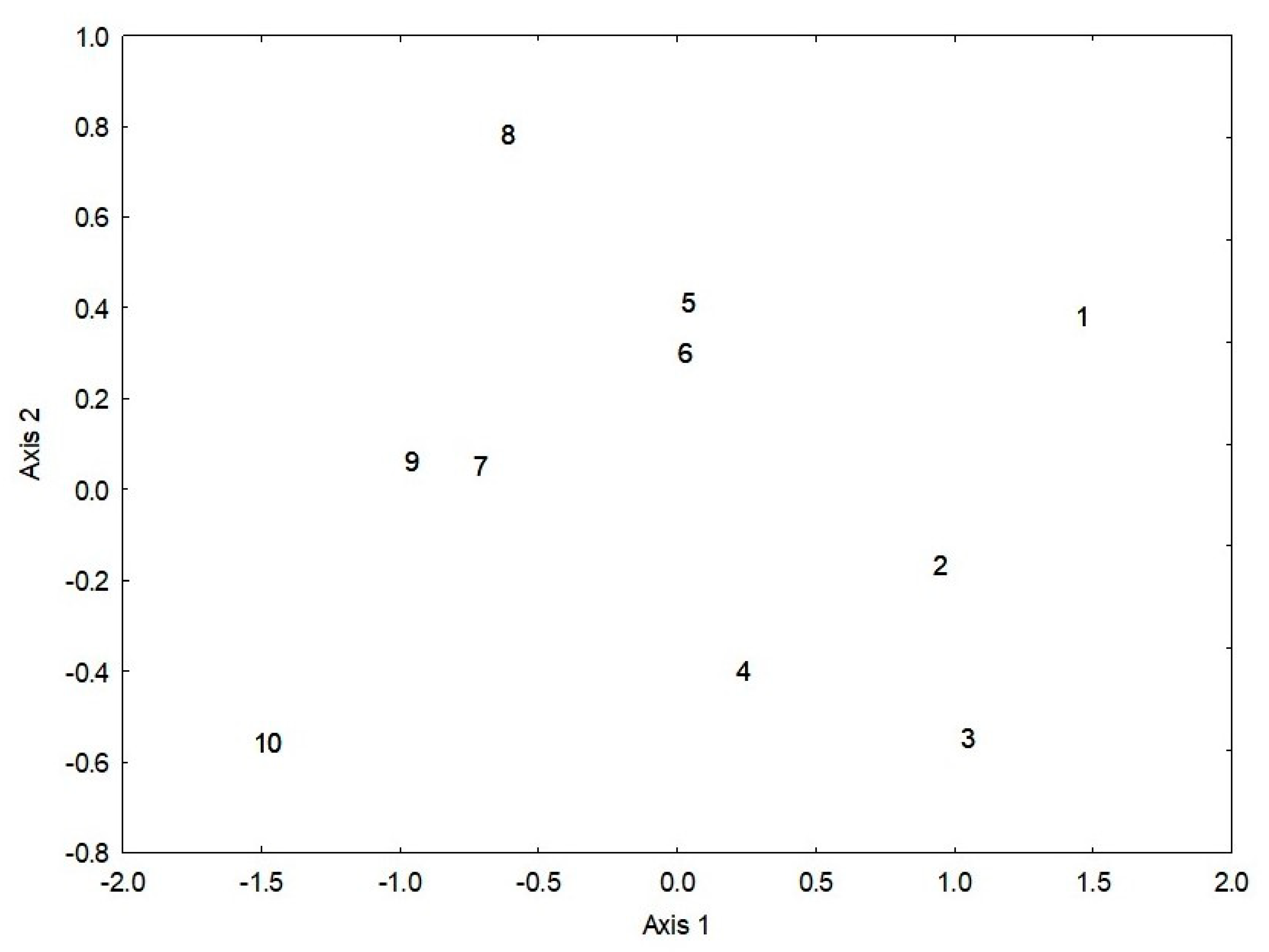
3.5. Biogeographic Patterns of Functional Group Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terborgh, J.; Robinson, S. Guilds and Their Utility in Ecology—Community Ecology: Patterns and Processes; Kikkawa, J., Anderson, D.J., Eds.; Blackwell Scientific Publications: Melbourne, Australia, 1986; pp. 65–90. [Google Scholar]
- Blaum, N.; Mosner, E.; Schwager, M.; Jeltsch, F. How functional is functional? Ecological groupings in terrestrial animal ecology: Towards an animal functional type approach. Biodivers. Conserv. 2011, 20, 2333–2345. [Google Scholar] [CrossRef]
- Rocha, M.R.; Gaedke, U.; Vasseur, D.A. Functionally similar species have similar dynamics. J. Ecol. 2011, 99, 1453–1459. [Google Scholar] [CrossRef]
- Andersen, A.N. A Classification of Australian Ant Communities, Based on Functional Groups Which Parallel Plant Life-Forms in Relation to Stress and Disturbance. J. Biogeogr. 1995, 22, 15. [Google Scholar] [CrossRef]
- Andersen, A. Functional groups and patterns of organization in North American ant communities: A comparison with Australia. J. Biogeogr. 1997, 24, 433–460. [Google Scholar] [CrossRef]
- Hernández-Ruiz, P.; Cano-Santana, Z.; Castaño-Meneses, G. Composition and functional groups of epiedaphic ants (Hymenoptera: Formicidae) in irrigated agroecosystem and in nonagricultural areas. Pesq. Agropec. Bras. 2009, 44, 904–910. [Google Scholar] [CrossRef]
- Castracani, C.; Grasso, D.A.; Fanfani, A.; Mori, A. The ant fauna of Castelporziano Presidential Reserve (Rome, Italy) as a model for the analysis of ant community structure in relation to environmental variation in Mediterranean ecosystems. J. Insect Conserv. 2010, 14, 585–594. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Wiens, J.A. The Effects of Land Use on the Structure of Ground-Foraging Ant Communities in the Argentine Chaco. Ecol. Appl. 1996, 6, 1225–1240. [Google Scholar] [CrossRef]
- Hoffmann, B.D.; Andersen, A.N. Responses of ants to disturbance in Australia, with particular reference to functional groups. Austral Ecol. 2003, 28, 444–464. [Google Scholar] [CrossRef]
- Linksvayer, T.A.; Janssen, M.A. Traits underlying the capacity of ant colonies to adapt to disturbance and stress regimes. Syst. Res. Behav. Sci. 2009, 26, 315–329. [Google Scholar] [CrossRef]
- Vepsäläinen, K.; Pisarski, B. Assembly of island ant communities. Ann. Zool. Fennici 1982, 19, 327–335. [Google Scholar]
- Savolainen, R.; Vepsäläinen, K. A Competition Hierarchy among Boreal Ants: Impact on Resource Partitioning and Community Structure. Oikos 1988, 51, 135. [Google Scholar] [CrossRef]
- Savolainen, R.; Vepsäläinen, K. Niche Differentiation of Ant Species within Territories of the Wood Ant Formica polyctena. Oikos 1989, 56, 3–16. [Google Scholar] [CrossRef]
- Savolainen, R. Colony success of the submissive ant Formica fusca within territories of the dominant Formica polyctena. Ecol. Èntomol. 1990, 15, 79–85. [Google Scholar] [CrossRef]
- Sheard, J.K.; Nelson, A.S.; Berggreen, J.D.; Boulay, R.; Dunn, R.R.; Sanders, N.J. Testing trade-offs and the dominance-impoverishment rule among ant communities. J. Biogeogr. 2020, 47, 1899–1909. [Google Scholar] [CrossRef]
- Cerdá, X.; Retana, J.; Manzaneda, A. The role of competition by dominants and temperature in the foraging of subordinate species in Mediterranean ant communities. Oecologia 1998, 117, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Luque, G.M.; López, J.R. Effect of experimental small-scale spatial heterogeneity on resource use of a Mediterranean ground-ant community. Acta Oecol. 2007, 32, 42–49. [Google Scholar] [CrossRef]
- Santini, G.; Tucci, L.; Ottonetti, L.; Frizzi, F. Competition trade-offs in the organisation of a Mediterranean ant assemblage. Ecol. Èntomol. 2007, 32, 319–326. [Google Scholar] [CrossRef]
- Cushman, J.H.; Lawton, J.H.; Manly, B.F.J. Latitudinal patterns in European ant assemblages: Variation in species richness and body size. Oecologia 1993, 95, 30–37. [Google Scholar] [CrossRef]
- Kumschick, S.; Schmidt-Entling, M.H.; Bacher, S.; Hickler, T.; Espadaler, X.; Nentwig, W. Determinants of local ant (Hymenoptera: Formicidae) species richness and activity density across Europe. Ecol. Èntomol. 2009, 34, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Arnan, X.; Cerda, X.; Retana, J. Distinctive life traits and distribution along environmental gradients of dominant and subordinate Mediterranean ant species. Oecologia 2012, 170, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Arnan, X.; Cerdá, X.; Retana, J. Ant functional responses along environmental gradients. J. Anim. Ecol. 2014, 83, 1398–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernadou, A.; Espadaler, X.; Le Goff, A.; Fourcassié, V. Ant community organization along elevational gradients in a temperate ecosystem. Insectes Sociaux 2015, 62, 59–71. [Google Scholar] [CrossRef]
- Blatrix, R.; Lebas, C.; Galkowski, C.; Wegnez, P.; Pimenta, R.; Morichon, D. Vegetation cover and elevation drive diversity and composition of ant communities (Hymenoptera: Formicidae) in a Mediterranean ecosystem. Myrmecol. News 2016, 22, 119–127. [Google Scholar]
- Wendt, C.F.; Ceia-Hasse, A.; Nunes, A.; Verble, R.; Santini, G.; Boieiro, M.; Branquinho, C. Local environmental variables are key drivers of ant taxonomic and functional beta-diversity in a Mediterranean dryland. Sci. Rep. 2021, 11, 2292. [Google Scholar] [CrossRef]
- Fisher, B.L.; Cover, S.P. Ants of North America: A Guide to the Genera; University of California Press: Berkeley, CA, USA, 2007. [Google Scholar]
- Fauna Europaea. Fauna Europaea Version 2.4. 2011. Available online: http://www.faunaeur.org (accessed on 5 July 2012).
- Gómez, C.; Casellas, D.; Oliveras, J.; Bas, J.M. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region. Biodivers. Conserv. 2003, 12, 2135–2146. [Google Scholar] [CrossRef]
- Ottonetti, L.; Tucci, L.; Santini, G. Recolonization Patterns of Ants in a Rehabilitated Lignite Mine in Central Italy: Potential for the Use of Mediterranean Ants as Indicators of Restoration Processes. Restor. Ecol. 2006, 14, 60–66. [Google Scholar] [CrossRef]
- Roig, X.; Espadaler, X. Proposal of functional groups of ants for the Iberian Peninsula and Balearic Islands, and their use as bioindicators. Iberomyrmex 2010, 2, 28–29. (In Spanish) [Google Scholar]
- Gómez, C.; Abril, S. Selective logging in public pine forests of the central Iberian Peninsula: Effects of the recovery process on ant assemblages. For. Ecol. Manag. 2011, 262, 1061–1066. [Google Scholar] [CrossRef]
- Dunn, R.R.; Agosti, D.; Andersen, A.; Arnan, X.; Brühl, C.; Cerda, X.; Ellison, A.; Fisher, B.; Fitzpatrick, M.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef]
- Arnan, X.; Andersen, A.N.; Gibb, H.; Parr, C.; Sanders, N.J.; Dunn, R.; Angulo, E.; Baccaro, F.; Bishop, T.; Boulay, R.; et al. Dominance-diversity relationships in ant communities differ with invasion. Glob. Change Biol. 2018, 24, 4614–4625. [Google Scholar] [CrossRef]
- Della Santa, E. Fourmis de Provence. Faune Provence 1995, 16, 5–38. [Google Scholar]
- Collingwood, C.A. Fauna Entomologica Scandinavica—Volume 8: The Formicidae (Hymenoptera) of Fennoscandia and Denmark; Scandinavian Science Press Ltd.: Klampenborg, Denmark, 1979. [Google Scholar]
- Seifert, B. A Taxonomic Revision of The Palaearctic Members of the Ant Subgenus Lasius s. str. (Hymenoptera, Formicidae). Abh. Ber. Nat. Görlitz 1992, 66, 1–67. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS-Statistical Estimation of Species Richness and Shared Species from Samples. 2005. Available online: http://www.purl.oclc.org/estimates (accessed on 1 March 2013).
- Guénard, B.; Weiser, M.D.; Dunn, R.R. Ant Genera of the World. 2010. Available online: http://www.antmacroecology.org/ant_genera/index.htm (accessed on 7 February 2011).
- Cerdá, X.; Retana, J.; Cros, S. Thermal Disruption of Transitive Hierarchies in Mediterranean Ant Communities. J. Anim. Ecol. 1997, 66, 363. [Google Scholar] [CrossRef]
- Abel, M.A.; Pons, P. Effect of prescribed burning on ground-foraging ant community in a Mediterranean Maquis. In Fire Management and Landscape Ecology; Trabaud, L., Ed.; International Association of Wildland Fire: Washington, USA, 1998; pp. 253–259. [Google Scholar]
- Punttila, P.; Haila, Y.; Pajunen, T.; Tukia, H. Colonisation of Clearcut Forests by Ants in the Southern Finnish Taiga: A Quantitative Survey. Oikos 1991, 61, 250. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v5: User Manual Tutorial: Primer-e; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Brown, W.L., Jr. Diversity of ants. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 45–79. [Google Scholar]
- Machac, A.; Janda, M.; Dunn, R.R.; Sanders, N.J. Elevational gradients in phylogenetic structure of ant communities reveal the interplay of biotic and abiotic constraints on diversity. Ecography 2011, 34, 364–371. [Google Scholar] [CrossRef]
- Silvestre, M.; Carmona, C.P.; Azcárate, F.M.; Seoane, J. Diverging facets of grassland ant diversity along a Mediterranean elevational gradient. Ecol. Èntomol. 2021, 46, 1301–1314. [Google Scholar] [CrossRef]
- Andersen, A. Diversity, Seasonality and Community Organization of Ants at Adjacent Heath and Woodland Sites in Southeastern Australia. Aust. J. Zool. 1986, 34, 53–64. [Google Scholar] [CrossRef]
- Mezger, D.; Pfeiffer, M. Partitioning the impact of abiotic fators and spatial patterns on species richness and community structure of ground ant assemblages in four Bornean rainforests. Ecography 2010, 34, 39–48. [Google Scholar] [CrossRef]
- Savolainen, R.; Wuorenrinne, H.; Vepsäläinen, K. Ant assemblages in the taiga biome: Testing the role of territorial wood ants. Oecologia 1989, 81, 481–486. [Google Scholar] [CrossRef]
- Gallé, L. Structure and succession of ant assemblages in a north European sand dune area. Ecography 1991, 14, 31–37. [Google Scholar] [CrossRef]
- Braschler, B.; Baur, B. Experimental small-scale grassland fragmentation alters competitive interactions among ant species. Oecologia 2005, 143, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, A.N. Using ants as bioindicators: Multi-scale issues in ant community ecology. Conserv. Ecol. 1997, 1, 1–17. [Google Scholar]
- Creighton, W.S. The ants of North America. Bull. Mus. Comp. Zool. 1950, 24, 1–585. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, B.D.; Andersen, A.N. Patterns of European Ant Communities Reveal a Functionally Coherent Holarctic Fauna. Diversity 2023, 15, 341. https://doi.org/10.3390/d15030341
Hoffmann BD, Andersen AN. Patterns of European Ant Communities Reveal a Functionally Coherent Holarctic Fauna. Diversity. 2023; 15(3):341. https://doi.org/10.3390/d15030341
Chicago/Turabian StyleHoffmann, Benjamin D., and Alan N. Andersen. 2023. "Patterns of European Ant Communities Reveal a Functionally Coherent Holarctic Fauna" Diversity 15, no. 3: 341. https://doi.org/10.3390/d15030341
APA StyleHoffmann, B. D., & Andersen, A. N. (2023). Patterns of European Ant Communities Reveal a Functionally Coherent Holarctic Fauna. Diversity, 15(3), 341. https://doi.org/10.3390/d15030341