



Classification of Wetland Forests and Scrub in the Western Balkans

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Preparation
2.3. Data Analysis
3. Results
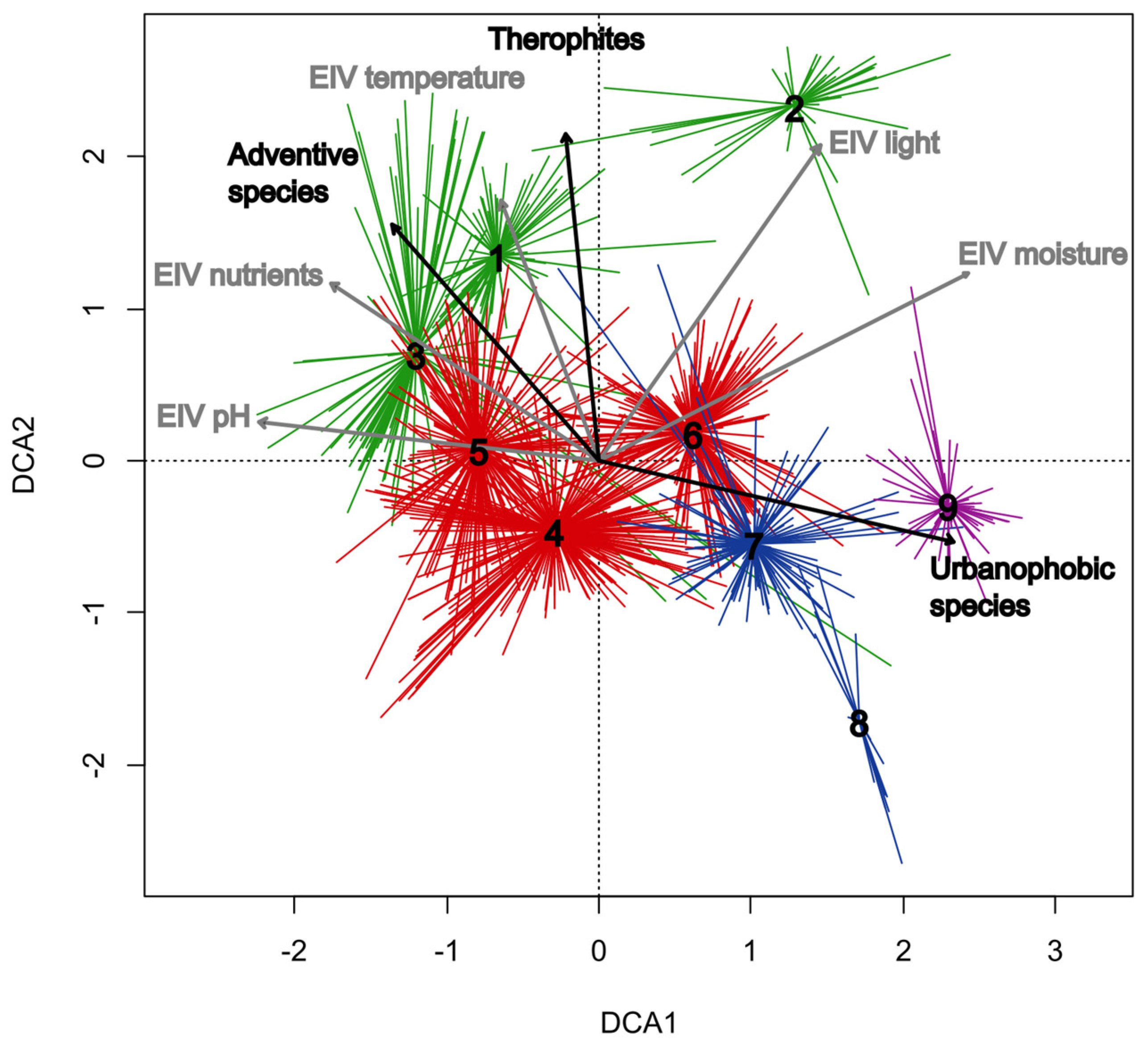
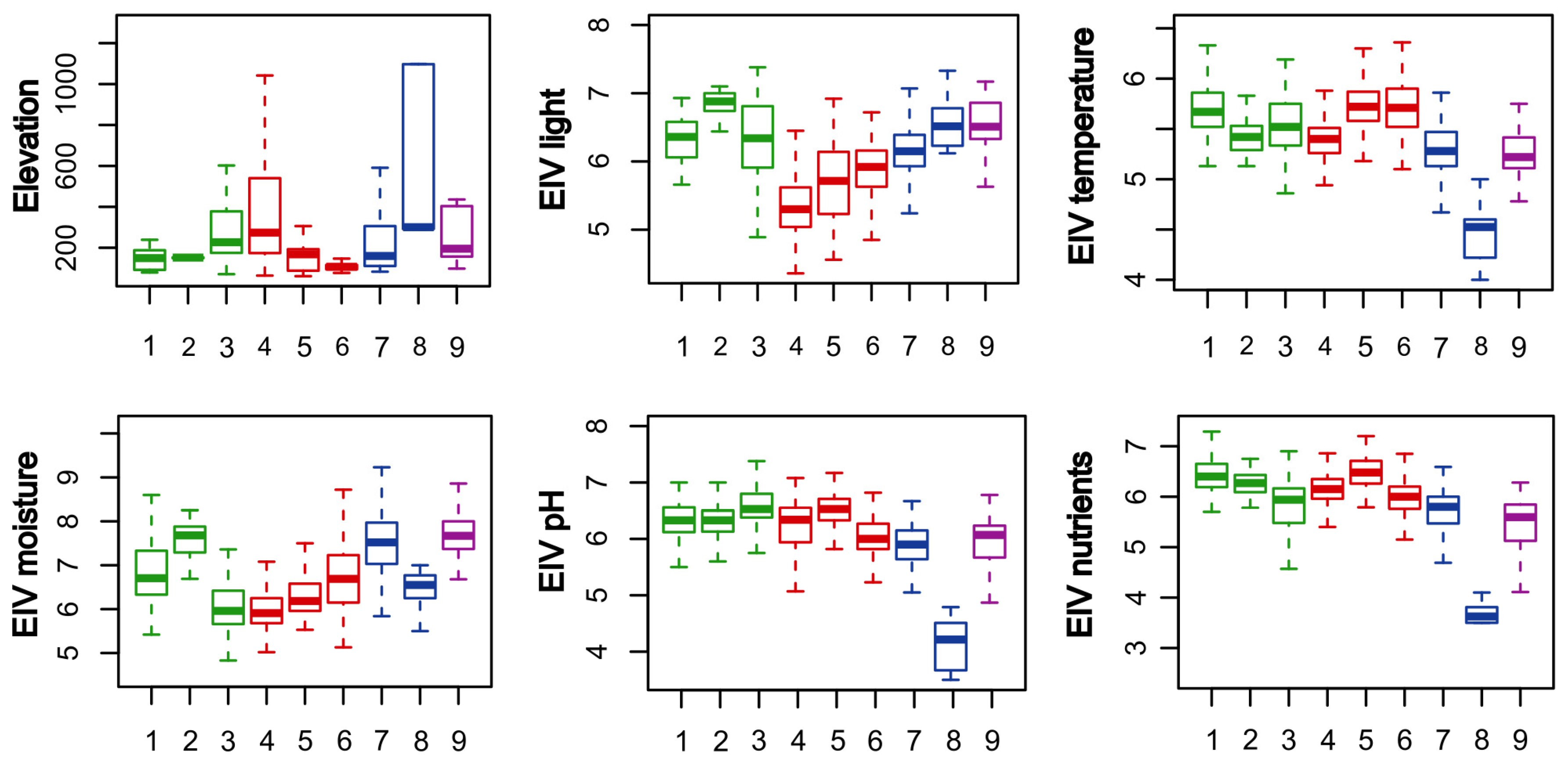
3.1. Classification and Ordination
3.2. Overview of the Classified Communities
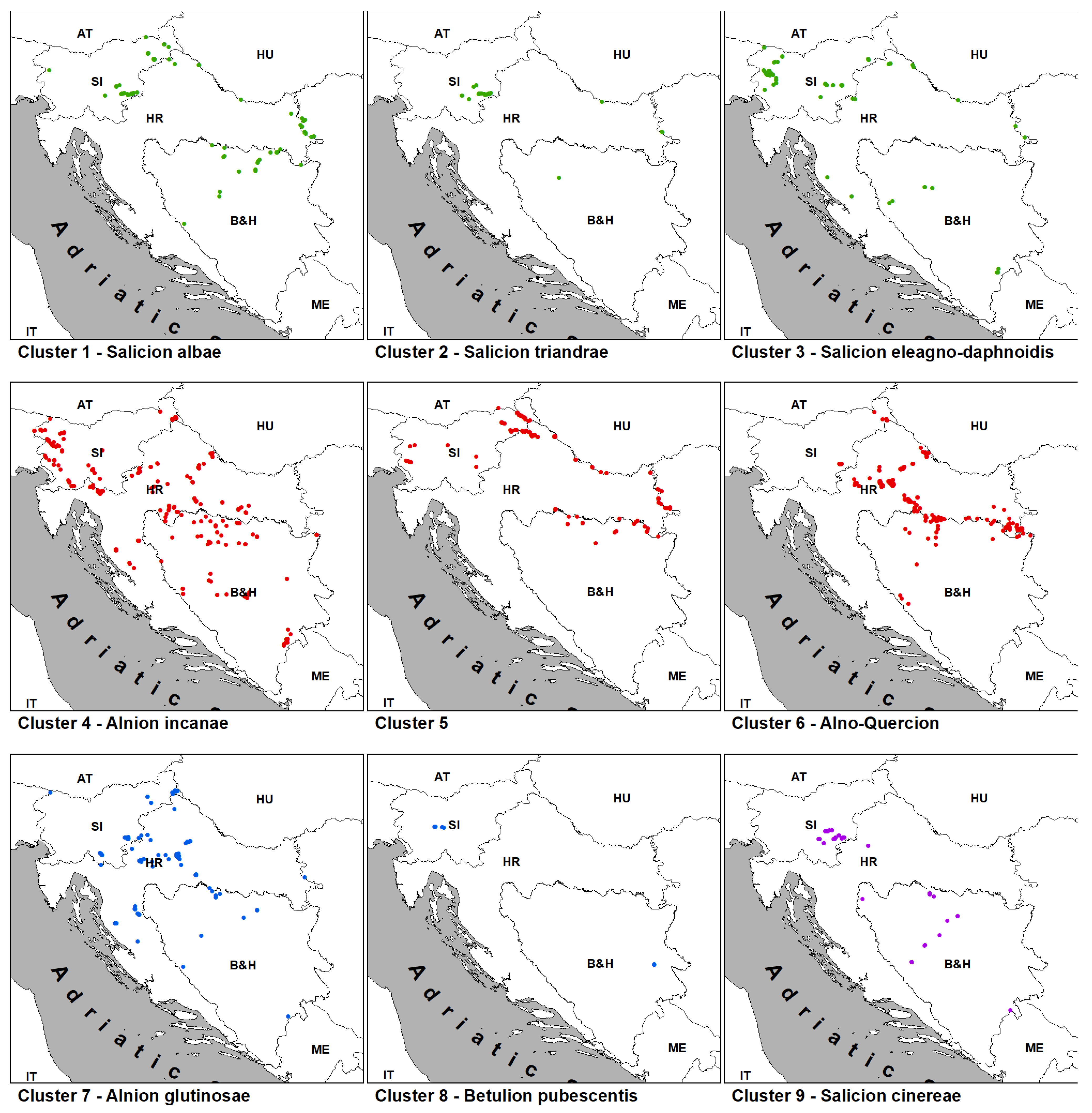
3.2.1. Salicetea purpureae Group of Clusters (Clusters 1–3; Table 1, Columns 1–3; Table A1)
3.2.2. Alno glutinosae-Populetea albae Group of Clusters (Clusters 4–6; Table 1, Columns 4–6; Table A2)
3.2.3. Alnetea glutinosae Group of Clusters (Clusters 7–8; Table 1, Columns 7–8; Table A3)
3.2.4. Franguletea Group of Clusters (Cluster 9; Table 1, Column 9)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Alnus glutinosa agg. (A. glutinosa and A. rohlenae) |
| Aquilegia vulgaris agg. (A. nigricans and A. vulgaris) |
| Aconitum variegatum agg. (A. variegatum and A. degenii) |
| Agrostis stolonifera agg. (A. gigantea and A. stolonifera) |
| Galium palustre agg. (G. palustre and G. elongatum) |
| Lamium galeobdolon agg. (L. galeobdolon ssp. argentatum, L. galeobdolon ssp. flavidum, L. galeobdolon ssp. galeobdolon and L. galeobdolon ssp. montanum) |
| Molinia caerulea agg. (M. caerulea and M. arundinacea) |
| Myosotis palustris agg. (M. scorpioides and Myosotis palustris) |
| Rubus fruticosus agg. (R. plicatus, R. silvaticus and Rubus fruticosus) |
| Crocus vernus agg. (Crocus vernus and C. vernus ssp. albiflorus) |
| Malus sylvestris agg. (M. sylvestris and M. pumila) |
| Ranunculus auricomus agg. (R. cassubicus. and R. auricomus) |
| Stellaria media agg. (S. media and S. neglecta) |
| Rosa canina (all species from Rosa canina group sensu Tutin et al. [103]) |
| Aconitum lycoctonum s.lat. (A. lycoctonum ssp. lycoctonum and A. lycoctonum ssp. vulparia) |
| Asarum europaeum s.lat. (A. europaeum ssp. caucasicum and A. europaeum ssp. europaeum) |
| Centaurea scabiosa s.lat (C. scabiosa ssp. scabiosa and Centaurea scabiosa ssp. fritschii) |
| Dactylis glomerata s.lat. (D. glomerata ssp. glomerata and D. glomerata ssp. lobata) |
| Fraxinus angustifolia s.lat. (F. angustifolia ssp. angustifolia and Fraxinus angustifolia ssp. oxycarpa) |
| Hesperis matronalis s.lat. (H. matronalis ssp. matronalis and H. matronalis ssp. candida) |
| Knautia drymeia s.lat. (K. drymeia ssp. drymeia and K. drymeia ssp. intermedia) |
| Phyteuma spicatum s.lat. (P. spicatum ssp. spicatum and P. spicatum ssp. coeruleum) |
| Plantago major s.lat (P. major ssp. major and P. major ssp. intermedia) |
| Prunus domestica s.lat. (P. domestica ssp. domestica and P. domestica ssp. insititia) |
| Solanum nigrum s.lat. (S. nigrum ssp. nigrum and S. nigrum ssp. schultesii) |
| Arabidopsis halleri s.lat. (A. halleri ssp. halleri and A. halleri ssp. ovirensis) |
| Carex divulsa s.lat. (C. divulsa ssp. divulsa and C. divulsa ssp. leersii) |
| Euphorbia esula s.lat (E. esula ssp. esula and E. esula ssp. tommasiniana) |
| Leucanthemum ircutianum s.lat. (L. ircutianum ssp. ircutianum and L. ircutianum ssp. leucolepis) |
| Pyrus communis s.lat. (P. communis ssp. communis and P. communis ssp. pyraster) |
| Rhamnus alpina s.lat. (R. alpina ssp. alpina and Rhamnus alpina ssp. fallax) |
| Helleborus dumetorum s.lat. (H. dumetorum ssp. dumetorum and H. dumetorum ssp. atrorubens) |
Appendix B
| Alnetum incanae Lüdi 1921 |
| Betulo-Quercetum roboris Martinčič 1987 |
| Cardamini parviflorae-Quercetum roboris Molnár Zs. 2010 |
| Carduo crispi–Populetum nigrae Kevey in Borhidi and Kevey 1996 |
| Carici acutiformis-Alnetum glutinosae Scamoni 1935 |
| Carici brizoides-Alnetum glutinosae Horvat 1938 |
| Carici elongatae-Alnetum glutinosae Koch 1926 |
| Carici elongatae-Quercetum Sokołowski 1972 |
| Carici paniculatae-Salicetum myrsinifoliae Dakskobler in Vreš, Seliškar et Dakskobler 2012 |
| Equiseto hyemali-Alnetum incanae Moor 1958 |
| Frangulo-Alnetum glutinosae Rauš 1971 (1973) |
| Fraxino excelsioris-Populetum albae Jurko 1958 |
| Fraxino angustifoliae-Ulmetum effusae Slavnić 1952 |
| Galio palustri–Salicetum albae Rauš 1973 |
| Genisto elatae-Quercetum roboris Horvat 1938 |
| Lamio orvalae-Alnetum glutinosae Dakskobler 2016 |
| Lamio orvalae-Alnetum incanae Dakskobler 2010 |
| Lamio orvalae-Salicetum albae Dakskobler 2016 |
| Lamio orvalae-Salicetum albae caricetosum pendulae Dakskobler 2016 |
| Lamio orvalae-Salicetum albae ranunculetosum lanuginosae Dakskobler 2016 |
| Lamio orvalae-Salicetum eleagni Dakskobler, Šilc and Čušin ex Dakskobler 2007 |
| Lamio orvalae-Salicetum purpureae nom. prov. (Dakskobler, 2016) |
| Leucojo aestivi-Fraxinetum angustifoliae Glavač 1959 |
| Petterio-Salicetum eleagni Redžić, Muratspahić and Lakušić 1992 |
| Pino-Betuletum pubescentis Stefanović 1961 |
| Populetum nigro-albae Slavnić 1952 |
| Pruno padi-Fraxinetum angustifoliae Čarni et al. 2008 |
| Pseudostellario europaeae-Quercetum roboris Accetto 1974 |
| Salicetum albae Issler 1926 |
| Salicetum cinereae Zólyomi 1931 |
| Salicetum cinereo-purpureae Pelcer 1975 prov. |
| Salicetum purpureae Wendelberger-Zelinka 1952 |
| Salicetum incano-purpureae Sillinger 1933 |
| Salicetum triandrae Malcuit ex Noirfalise in Lebrun et al. 1955 |
| Salici eleagni-Juniperetum communis Poldini, Francescato, Vidali and Castello 2020 |
| Salici purpureae-Myricarietum germanicae Moor 1958 |
| Salici-Populetum nigrae (R. Tx. 1931) Meyer Drees 1936 |
| Saponario officinalis-Salicetum purpureae Tchou 1948 |
| Sphagno nemorei-Betuletum pubescentis (Libbert 1933) Passarge 1968 |
| Stellario nemorum-Alnetum glutinosae Lohmeyer 1957 |
Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Number | 1 | 2 | 3 |
|---|---|---|---|
| Number of Relevés | 90 | 37 | 83 |
| Salicion albae | |||
| Salix alba | 9172.6 | 8 | 23 |
| Galium aparine | 5642.9 | 14 | 16 |
| Glechoma hederacea | 4441.1 | 11 | 7 |
| Iris pseudacorus | 4340.9 | 14 | 4 |
| Salix euxina | 3636.9 | . | 13 |
| Rubus caesius | 7333 | 19 | 58 |
| Euonymus europaeus | 3232.2 | 3 | 12 |
| Salicion triandrae | |||
| Salix triandra | 19 | 9778.6 | 11 |
| Alisma plantago-aquatica | 1 | 3850.8 | 1 |
| Rorippa sylvestris | 6 | 4948.3 | 8 |
| Solanum dulcamara | 38 | 8146.2 | 27 |
| Agrostis stolonifera agg. | 34 | 7042.2 | 18 |
| Persicaria hydropiper | 28 | 5741.7 | 5 |
| Bidens tripartitus | 14 | 4340 | 4 |
| Echinocystis lobata | 39 | 5937.3 | 5 |
| Lythrum salicaria | 27 | 5734.3 | 18 |
| Galium palustre agg. | 29 | 4932.8 | 6 |
| Phalaroides arundinacea | 56 | 7832 | 34 |
| Rorippa amphibia | 10 | 3031.9 | 2 |
| Calystegia sepium | 46 | 6230.9 | 14 |
| Salicion eleagno-daphnoidis | |||
| Salix eleagnos | 4 | . | 7073.9 |
| Salix purpurea | 18 | 16 | 8465 |
| Petasites hybridus | 1 | . | 5364.3 |
| Clematis vitalba | 3 | . | 4354.2 |
| Brachypodium sylvaticum | 13 | . | 5352.6 |
| Fraxinus excelsior | 2 | . | 3951.5 |
| Acer pseudoplatanus | . | . | 3551.3 |
| Deschampsia cespitosa | 4 | . | 4049.9 |
| Eupatorium cannabinum | 7 | . | 4249.6 |
| Salvia glutinosa | . | . | 3148.3 |
| Chaerophyllum hirsutum | 3 | . | 3547 |
| Equisetum arvense | 6 | . | 3746.6 |
| Ranunculus lanuginosus | 1 | . | 3045.6 |
| Saponaria officinalis | 3 | . | 3143.7 |
| Cirsium oleraceum | 11 | 3 | 3939.3 |
| Other species with high frequency | |||
| Urtica dioica | 8215.7 | 86 | 48 |
| Cornus sanguinea | 43 | . | 43 |
| Persicaria dubia | 12 | 43 | 29 |
| Angelica sylvestris | 24 | 11 | 4629.9 |
| Poa trivialis | 4327.6 | 14 | 22 |
| Lamium maculatum | 29 | 24 | 24 |
| Humulus lupulus | 4329 | 27 | 6 |
| Alliaria petiolata | 34 | 14 | 27 |
| Ranunculus repens | 23 | 35 | 16 |
| Aegopodium podagraria | 26 | 11 | 35 |
| Cluster Number | 4 | 5 | 6 |
|---|---|---|---|
| Number of Relevés | 281 | 178 | 226 |
| Alnion incanae | |||
| Acer pseudoplatanus | 5059.5 | 3 | 1 |
| Fraxinus excelsior | 4750.7 | 8 | 2 |
| Ranunculus lanuginosus | 3245.7 | 2 | 1 |
| Petasites hybridus | 3144.5 | 2 | . |
| Chaerophyllum hirsutum | 3043.5 | 2 | . |
| Equisetum arvense | 3943.2 | 9 | 1 |
| Corylus avellana | 6342.9 | 20 | 19 |
| Alnus glutinosa agg. | 8139.6 | 31 | 47 |
| Brachypodium sylvaticum | 6435.4 | 46 | 8 |
| Lamium galeobdolon agg. | 4935.1 | 24 | 8 |
| Lamium orvala | 3334.3 | 13 | . |
| Aegopodium podagraria | 7033 | 57 | 14 |
| Sambucus nigra | 7431.5 | 61 | 21 |
| Angelica sylvestris | 3731.1 | 12 | 10 |
| Cluster 5 | |||
| Populus nigra | 7 | 5457 | 2 |
| Prunus padus | 5 | 4045.7 | 4 |
| Populus alba | 1 | 3843.9 | 8 |
| Salix alba | 21 | 4739.8 | 1 |
| Acer negundo | 3 | 3038.5 | 4 |
| Solidago gigantea | 14 | 4438.3 | 7 |
| Impatiens glandulifera | 8 | 3037 | . |
| Galium aparine | 27 | 6236.2 | 23 |
| Ulmus laevis | 4 | 5433.6 | 388.7 |
| Alno-Quercion | |||
| Fraxinus angustifolia s.lat. | 9 | 43 | 8454.5 |
| Galium palustre agg. | 8 | 10 | 5853.1 |
| Quercus robur | 11 | 37 | 7952.5 |
| Stachys palustris | 2 | 10 | 4646.8 |
| Ulmus minor | 10 | 14 | 5243.5 |
| Crataegus laevigata | 9 | 3 | 3942 |
| Persicaria hydropiper | 15 | 10 | 5040.6 |
| Iris pseudacorus | 6 | 29 | 5438.5 |
| Myosotis palustris agg. | 10 | 2 | 3538.1 |
| Lysimachia nummularia | 23 | 24 | 6237.2 |
| Leucojum aestivum | 2 | 9 | 3336.3 |
| Acer tataricum | 8 | 1 | 3136.3 |
| Rumex sanguineus | 12 | 12 | 4234.5 |
| Carex remota | 33 | 17 | 6134.5 |
| Lythrum salicaria | 6 | 7 | 3132.9 |
| Other species with high frequency | |||
| Rubus caesius | 70 | 9023.4 | 69 |
| Urtica dioica | 59 | 66 | 61 |
| Cornus sanguinea | 6513.6 | 64 | 37 |
| Euonymus europaeus | 5514.7 | 47 | 31 |
| Crataegus monogyna | 48 | 34 | 46 |
| Acer campestre | 5721.1 | 21 | 48 |
| Geum urbanum | 4716.9 | 30 | 29 |
| Ranunculus repens | 37 | 16 | 4821.1 |
| Hedera helix | 4119.5 | 22 | 23 |
| Ligustrum vulgare | 34 13.9 | 30 | 13 |
| Cluster Number | 7 | 8 |
|---|---|---|
| Number of Relevés | 121 | 14 |
| Alnion glutinosae | ||
| Alnus glutinosa agg. | 9494.4 | . |
| Lycopus europaeus | 6569.6 | . |
| Carex elongata | 5561.2 | . |
| Iris pseudacorus | 5259.3 | . |
| Rubus caesius | 4654.9 | . |
| Filipendula ulmaria | 4554.2 | . |
| Lythrum salicaria | 4453 | . |
| Urtica dioica | 4352.3 | . |
| Betulion pubsecentis | ||
| Betula pubescens | . | 8686.6 |
| Molinia caerulea agg. | 7 | 9386.2 |
| Pinus sylvestris | . | 7174.5 |
| Sorbus aucuparia | 2 | 5760.9 |
| Betula pendula | 1 | 5056.5 |
| Rubus hirtus s.lat. | 1 | 4350.9 |
| Salix aurita | 2 | 4349.5 |
| Calamagrostis villosa | . | 3646.6 |
| Lonicera nigra | . | 3646.6 |
| Cirsium palustre | . | 3646.6 |
| Parnassia palustris | . | 3646.6 |
| Knautia sarajevensis | . | 3646.6 |
| Carex rostrata | . | 3646.6 |
| Salix pentandra | 1 | 3645.1 |
| Equisetum sylvaticum | 1 | 3645.1 |
| Other species with high frequency | ||
| Frangula alnus | 66 | 100 |
| Dryopteris carthusiana | 53 | 71 |
| Lysimachia vulgaris | 67 | 43 |
| Galium palustre agg. | 60 | 36 |
| Viburnum opulus | 53 | 36 |
| Peucedanum palustre | 52 | 36 |
| Caltha palustris | 48 | 36 |
| Solanum dulcamara | 54 | 29 |
| Valeriana dioica s.lat. | 50 | 21 |
| Quercus robur | 33 | 36 |
References
- Dierschke, H. Pflanzensoziologie; Eugen Ulmer Verlag: Stuttgart, DE, USA, 1994. [Google Scholar]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Stupar, V.; Čarni, A. Ecological, Floristic and Functional Analysis of Zonal Forest Vegetation in Bosnia and Herzegovina. Acta Bot. Croat. 2017, 76, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Junk, W.J.; Piedade, M.T.F. An Introduction to South American Wetland Forests: Distribution, Definitions and General Characterization. In Amazonian Floodplain Forests; Junk, W.J., Piedade, M.T.F., Wittmann, F., Schöngart, J., Parolin, P., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; pp. 3–25. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical Floristic Classification System of Vascular Plant, Bryophyte, Lichen, and Algal Communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Eremiášová, R.; Skokanová, H. Response of Vegetation on Gravel Bars to Management Measures and Floods: Case Study from the Czech Republic. Ekologia 2014, 33, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Mandžukovski, D.; Čarni, A.; Biurrun, I.; Douda, J.; Škvorc, Ž.; Stupar, V.; Slezák, M.; Ćušterevska, R.; Rodríguez-González, P.; Mendías, C.; et al. Interpretative Manual of European Riparian Forests and Shrublands; Ss Cyril and Methodius University in Skopje, Hans Em Faculty of Forest Sciences, Landscape Architecture and Environmental Engineering: Skopje, Republic of Macedonia, 2021; ISBN 978-9989-132-22-3. [Google Scholar]
- Vogt, K.; Rasran, L.; Jensen, K. Water-Borne Seed Transport and Seed Deposition during Flooding in a Small River-Valley in Northern Germany. Flora–Morphol. Distrib. Funct. Ecol. Plants 2004, 199, 377–388. [Google Scholar] [CrossRef]
- Vukelić, J. Šumska Vegetacija Hrvatske; Šumarski fakultet, Sveučilište u Zagrebu, DZZP: Zagreb, Croatia, 2012; ISBN 978-953-292-024-6. [Google Scholar]
- Slezák, M.; Hrivnák, R.; Petrášová, A. Numerical Classification of Alder Carr and Riparian Alder Forests in Slovakia. Phytocoenologia 2014, 44, 283–308. [Google Scholar] [CrossRef]
- Douda, J.; Boublík, K.; Slezák, M.; Biurrun, I.; Nociar, J.; Havrdová, A.; Doudová, J.; Aćić, S.; Brisse, H.; Brunet, J.; et al. Vegetation Classification and Biogeography of European Floodplain Forests and Alder Carrs. Appl. Veg. Sci. 2016, 19, 147–163. [Google Scholar] [CrossRef]
- Rodríguez-González, P.M.; Abraham, E.; Aguiar, F.; Andreoli, A.; Baležentienė, L.; Berisha, N.; Bernez, I.; Bruen, M.; Bruno, D.; Camporeale, C.; et al. Bringing the Margin to the Focus: 10 Challenges for Riparian Vegetation Science and Management. WIREs Water 2022, 9, e1604. [Google Scholar] [CrossRef]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global Overview of Ecosystem Services Provided by Riparian Vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A. Wetlands in Europe: Perspectives for Restoration of a Lost Paradise. Ecol. Eng. 2014, 66, 6–9. [Google Scholar] [CrossRef]
- Cestarić, D.; Škvorc, Ž.; Franjić, J.; Sever, K.; Krstonošić, D. Forest Plant Community Changes in the Spačva Lowland Area. Plant Biosyst. 2017, 151, 584–597. [Google Scholar] [CrossRef]
- Havrdová, A.; Douda, J.; Doudová, J. Threats, Biodiversity Drivers and Restoration in Temperate Floodplain Forests Related to Spatial Scales. Sci. Total Environ. 2023, 854, 158743. [Google Scholar] [CrossRef]
- Janssen, J.A.M.; Rodwell, J.S.; Garcia Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssymank, A.; et al. European Red List of Habitats—Part 2. Terrestrial and Freshwater Habitats; European Commission: Brussels, Belgium, 2016; ISBN 978-92-79-61588-7. [Google Scholar]
- Council of the European Communities. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Off. J. Eur. Communities 1992, L206, 7–50. [Google Scholar]
- Kalníková, V.; Chytrý, K.; Biţa-Nicolae, C.; Bracco, F.; Font, X.; Lakushenko, D.; Kącki, Z.; Kudrnovsky, H.; Landucci, F.; Lustyk, P.; et al. Vegetation of the European Mountain River Gravel Bars: A Formalized Classification. Appl. Veg. Sci. 2021, 24, e12542. [Google Scholar] [CrossRef]
- Biurrun, I.; Campos, J.A.; García-Mijangos, I.; Herrera, M.; Loidi, J. Floodplain Forests of the Iberian Peninsula: Vegetation Classification and Climatic Features. Appl. Veg. Sci. 2016, 19, 336–354. [Google Scholar] [CrossRef]
- Borhidi, A.; Kevey, B.; Lendvai, G. Plant Communities of Hungary; Akadémiai Kiadó: Budapest, Hungary, 2012. [Google Scholar]
- Chytry, M.; Douda, J.; Roleček, J.; Sádlo, J.; Boublík, K.; Hédl, R.; Vítková, M.; Zelený, D.; Navrátilová, J.; Neuhäuslová, Z.; et al. Vegetace České Republiky 4. Lesní a Křovinná Vegetace (Vegetation of the Czech Republic 4. Forest and Scrub Vegetation); Chytrý, M., Ed.; Academia: Praha, CZ, USA, 2013. [Google Scholar]
- Poldini, L.; Vidali, M.; Ganis, P. Riparian Salix Alba: Scrubs of the Po Lowland (N-Italy) from an European Perspective. Plant Biosyst. 2011, 145, 132–147. [Google Scholar] [CrossRef]
- Valachovič, M.; Kliment, J.; Hegedüšová Vantarová, K. Rastlinné Spoločenstvá Slovenska 6. Vegetácia Lesov a Krovín; Veda vydavateľstvo Slovenskej akadémie vied: Bratislava, Slovakia, 2021. [Google Scholar]
- Willner, W.; Grabherr, G.; Drescher, A.; Eichberger, C.; Exner, A.; Franz, W.R.; Grabner, S.; Heiselmayer, P.; Karner, P.; Steiner, G.M.; et al. Die Wälder und Gebüsche Österreichs: Ein Bestimmungswerk mit Tabellen (in zwei Bänden); Willner, W., Grabherr, G., Eds.; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Barudanović, S.; Macanović, A.; Topalić-Trivunović, L.; Cero, M. Ekosistemi Bosne i Hercegovine u Funkciji Održivog Razvoja; Univerzitet u Sarajevu, Prirodno-matematički fakultet: Sarajevo, BA, USA, 2015; ISBN 978-9958-592-60-7. [Google Scholar]
- Šilc, U.; Čarni, A. Conspectus of Vegetation Syntaxa in Slovenia. Hacquetia 2012, 11, 113–164. [Google Scholar] [CrossRef] [Green Version]
- Škvorc, Ž.; Jasprica, N.; Alegro, A.; Kovačić, S.; Franjić, J.; Krstonošić, D.; Vraneša, A.; Čarni, A. Vegetation of Croatia: Phytosociological Classification of the High-Rank Syntaxa. Acta Bot. Croat. 2017, 76, 200–224. [Google Scholar] [CrossRef] [Green Version]
- Trinajstić, I. Biljne Zajednice Republike Hrvatske; Akademija šumarskih znanosti: Zagreb, Croatia, 2008. [Google Scholar]
- Fabijanić, B.; Fukarek, P.; Stefanović, V. Lepenica: Pregled osnovnih tipova šumske vegetacije. Naučn. Druš. BH Poseb. Izd. 1963, 3, 85–129. [Google Scholar]
- Fukarek, P. Šumske zajednice prašumskog rezervata Perućice u Bosni. Akad. Nauk. Umjet. BH Od. Prir. Druš. Nauk. Poseb. Izd. 1970, 15, 157–262. [Google Scholar]
- Fukarek, P. Hrastove šume bosanskog posavlja u prošlosti i sadašnjosti. Jugosl. Akad. Znan. Umjet. Poseb. Izd. 1975, 2, 371–379. [Google Scholar]
- Stefanović, V.; Sokač, A. Fitocenoza bijelog bora i maljave breze na rubu tresetišta kod Han-krama (Pineto-Betuletum pubescentis, Stef.). Rad. Naučn. Druš. BH 1962, 5, 97–126. [Google Scholar]
- Horvat, I. Biljnosociološka Istraživanja Šuma u Hrvatskoj. Glas. Šumske Pokuse 1938, 6, 127–279. [Google Scholar]
- Horvat, I.; Glavač, V.; Ellenberg, H. Vegetation Südosteuropas; Geobotanica selecta; Gustav Fischer Verlag: Stuttgart, DE, USA, 1974. [Google Scholar]
- Preislerová, Z.; Jiménez-Alfaro, B.; Mucina, L.; Berg, C.; Bonari, G.; Kuzemko, A.; Landucci, F.; Marcenò, C.; Monteiro-Henriques, T.; Novák, P.; et al. Distribution Maps of Vegetation Alliances in Europe. Appl. Veg. Sci. 2022, 25, e12642. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). Biogeographic Regions in Europe; European Environment Agency (EEA): Copenhagen, Denmark, 2011. [Google Scholar]
- Mrgić, J. Severna Bosna: 13–16. Vek; Istorijski Institut: Beograd, RS, USA, 2008; ISBN 1019-469X. [Google Scholar]
- Mrgić, J. Lijevče polje–beleške o naseljima i prirodi 15-19. vek. Hist. Rev. 2007, 5, 171–199. [Google Scholar]
- Dakskobler, I.; Kutnar, L.; Šilc, U. Poplavni, Močvirni in Obrežni Gozdovi v Sloveniji. Gozdovi vrb, jelš, Dolgopecljatega Bresta, Velikega in Ozkolistnega Jesena, doba in Rdečega bora ob Rekah in Potokih; Gozdarski inštitut Slovenije: Ljubljana, SI, USA, 2013. [Google Scholar]
- Košir, P.; Čarni, A.; Marinšek, A.; Šilc, U. Floodplain Forest Communities along the Mura River (NE Slovenia). Acta Bot. Croat. 2013, 72, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Dakskobler, I. Phytosociological Analysis of Riverine Forests in the Vipava and Reka Valleys (Southwestern Slovenia). Folia Biol. Geol. 2016, 57, 5–61. [Google Scholar] [CrossRef] [Green Version]
- Dakskobler, I.; Šilc, U.; Čušin, B. Riverine Forests in the Upper Soča Valley (the Julian Alps, Western Slovenia). Hacquetia 2004, 3, 51–80. [Google Scholar]
- Hennekens, S.M.; Schaminée, J.H.J. TURBOVEG, a Comprehensive Data Base Management System for Vegetation Data. J. Veg. Sci. 2001, 12, 589–591. [Google Scholar] [CrossRef] [Green Version]
- Tichý, L. JUICE, Software for Vegetation Classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Euro+Med Euro+Med PlantBase–the Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/query.asp (accessed on 12 October 2015).
- Vít, P.; Douda, J.; Krak, K.; Havrdová, A.; Mandák, B. Two New Polyploid Species Closely Related to Alnus Glutinosa in Europe and North Africa–An Analysis Based on Morphometry, Karyology, Flow Cytometry and Microsatellites. Taxon 2017, 66, 567–583. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data. Version 5.0 1999; Fundacion BBVA: Bilbao, Spain, 2014. [Google Scholar]
- Tichý, L.; Hennekens, S.M.; Novák, P.; Rodwell, J.S.; Schaminée, J.H.J.; Chytrý, M. Optimal Transformation of Species Cover for Vegetation Classification. Appl. Veg. Sci. 2020, 23, 710–717. [Google Scholar] [CrossRef]
- Botta-Dukát, Z.; Chytrý, K.; Hájková, P.; Havlová, M. Vegetation of Lowland Wet Meadows along a Climatic Continentality Gradient in Central Europe. Preslia 2005, 77, 89–111. [Google Scholar]
- Tichý, L.; Chytrý, M. Statistical Determination of Diagnostic Species for Site Groups of Unequal Size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di bioindicazione delle piante vascolari della flora d’Italia. Braun-Blanquetia 2005, 39, 1–97. [Google Scholar]
- Zelený, D.; Schaffers, A.P. Too Good to Be True: Pitfalls of Using Mean Ellenberg Indicator Values in Vegetation Analyses. J. Veg. Sci. 2012, 23, 419–431. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Gajić, M. Pregled vrsta flore SR Srbije sa biljnogeografskim oznakama. Glas. Šumar. Fak. 1980, 54, 111–141. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. (Eds.) BIOLFLOR–Eine Datenbank Zu Biologisch-Ökologischen Merkmalen Der Gefäßpflanzen in Deutschland; Bundesamt für Naturschutz: Bonn, DE, USA, 2002. [Google Scholar]
- Novák, P.; Willner, W.; Zukal, D.; Kollár, J.; Roleček, J.; Świerkosz, K.; Ewald, J.; Wohlgemuth, T.; Csiky, J.; Onyshchenko, V.; et al. Oak-Hornbeam Forests of Central Europe: A Formalized Classification and Syntaxonomic Revision. Preslia 2020, 92, 1–34. [Google Scholar] [CrossRef]
- Kočí, M.; Chytrý, M.; Tichý, L. Formalized Reproduction of an Expert-Based Phytosociological Classification: A Case Study of Subalpine Tall-Forb Vegetation. J. Veg. Sci. 2003, 14, 601–610. [Google Scholar] [CrossRef]
- Willner, W.; Jiménez-Alfaro, B.; Agrillo, E.; Biurrun, I.; Campos, J.A.; Čarni, A.; Casella, L.; Csiky, J.; Ćušterevska, R.; Didukh, Y.P.; et al. Classification of European Beech Forests: A Gordian Knot? Appl. Veg. Sci. 2017, 20, 494–512. [Google Scholar] [CrossRef]
- Čarni, A.; Košir, P.; Marinček, L.; Marinšek, A.; Šilc, U.; Zelnik, I. Komentar k Vegetacijski Karti Gozdnih Združb Slovenije v Merilu 1: 50.000-List Murska Sobota; Pomurska Akademsko Znanstvena Unija-PAZU: Murska Sobota, Slovenia, 2008; ISBN 961-91503-6-8. [Google Scholar]
- Dakskobler, I. Development of Vegetation on Gravel Sites of the Idrijca River in Western Slovenia. Folia Biol. Geol. 2010, 51, 5–90. [Google Scholar]
- Dakskobler, I.; Rozman, A. Phytosociological Analysis of Riverine Forests along the Sava Bohinjka, Radovna, Učja and Slatenik Rivers in Northwestern Slovenia. Folia Biol. Geol. 2013, 54, 37–105. [Google Scholar]
- Šilc, U. Vegetation of the Class Salicetea Purpureae in Dolenjska (SE Slovenia). Fitosociologia 2003, 40, 3–27. [Google Scholar]
- Rauš, Đ. Vegetacija Ritskih Šuma Dijela Podunavlja Od Aljmaša Do Iloka. Glas. Šum. Pokuse 1976, 19, 5–75. [Google Scholar]
- Rauš, Đ. Vegetacija Podravskih Ritskih Šuma u Okolici Legrada Na Ušću Mure u Dravu; Trinajstić, I., Ed.; Šumarski fakultet i Hrvatske šume po Zagreb: Zagreb, Croatia, 1994; pp. 87–100. [Google Scholar]
- Vukelić, J.; Baričević, D.; Perković, Z. Vegetacijske i Druge Značajke Zaštičenog Dijela “Slatinskih Podravskih Šuma”. Šumar. List 1999, 123, 287–299. [Google Scholar]
- Milanović, Đ.; Stupar, V. Riparian forest communities along watercourses in the Sutjeska National Park (SE Bosnia and Herzegovina). Glas. Šumar. Fak. Univ. Banjal. 2017, 26, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Vilhar, U.; Čarni, A.; Božič, G. Growth and vegetation characteristics of European black poplar (Populus nigra L.) in a floodplain forest along the river Sava and temperature differencies among selected sites. Folia Biol. Geol. 2013, 54, 193–214. [Google Scholar]
- Poldini, L.; Vidali, M.; Castello, M.; Sburlino, G. A Novel Insight into the Remnants of Hygrophilous Forests and Scrubs of the Po Plain Biogeographical Transition Area (Northern Italy). Plant Soc. 2020, 57, 17–69. [Google Scholar] [CrossRef]
- Redžić, S.; Muratspahić, D.; Lakušić, R. Neke fitocenoze šuma i šikara iz doline Neretve. Poljopr. Šumar. Podgor. 1992, 38, 95–101. [Google Scholar]
- Accetto, M. Močvirni in Poplavni Gozdovi. Zasnova Rajonizacije Ekosistemov Slovenije; Oddelek za Biologijo, Biotehniška Fakulteta: Ljubljana, Slovenia, 1994. [Google Scholar]
- Plišo Vusić, I.; Šapić, I.; Vukelić, J. Prepoznavanje i Kartiranje Šumskih Staništa Natura 2000 u Hrvatskoj (I)–91E0*, Aluvijalne Šume s Crnom Johom Alnus Glutinosa i Običnim Jasenom Fraxinus Excelsior (Alno-Padion, Alnion Incanae, Salicion Albae). Šumar. List 2019, 5–6, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Vukelić, J.; Šapić, I.; Alegro, A.; Šegota, V.; Stankić, I.; Baričević, D. Phytocoenological Analysis of Grey Alder (Alnus incana L.) Forests in the Dinarides of Croatia and Their Relationship with Affiliated Communities. Tuexenia 2017, 37, 65–78. [Google Scholar] [CrossRef]
- Vukelić, J.; Baričević, D.; Poljak, I.; Vrček, M.; Šapić, I. Fitocenološka analiza šuma bijele johe (Alnus incana/L./Moench subsp. incana) u Hrvatskoj. Šumar. List 2018, 142, 123–135. [Google Scholar] [CrossRef]
- Brullo, S.; Spampinato, G. Syntaxonomy of Hygrophilous Woods of the Alno-Quercion Roboris. Ann. Bot. 1999, 57, 133–146. [Google Scholar]
- Rauš, Đ. Šuma crne johe (Frangulo-Alnetum glutinosae Rauš 68) u bazenu Spačva. Šumar. List 1975, 99, 431–446. [Google Scholar]
- Sciandrello, S.; Angiolini, C.; Bacchetta, G.; Cutini, M.; Dumoulin, J.; Fois, M.; Gabellini, A.; Gennai, M.; Gianguzzi, L.; Landi, M.; et al. Alnus glutinosa Riparian Woodlands of Italy and Corsica: Phytosociological Classification and Floristic Diversity. Land 2023, 12, 88. [Google Scholar] [CrossRef]
- Poldini, L.; Sburlino, G. Lamio orvalae-Alnetum glutinosae Dakskobler 2016, nuova associazione ripariale per l’Italia settentrionale (Friuli Venezia Giulia, Veneto e Lombardia) con note sulle cenosi corrispondenti poste a sud del Po. Gortania 2020, 42, 5–21. [Google Scholar]
- Baričević, D. Ecological-Vegetational Properties of Forest “Žutica”. Glas. Šum. Pokuse 1998, 35, 1–91. [Google Scholar]
- Glavač, V. O Vlažnom Tipu Hrasta Lužnjaka i Običnog Graba. Šumar. List 1961, 85, 342–347. [Google Scholar]
- Rauš, Đ. Fitocenološke Značajke i Vegetacijska Karta Fakultetskih Šuma Lubardenik i Opeke. Šumar. List 1973, 97, 190–221. [Google Scholar]
- Stefanović, V. Fitocenologija Sa Pregledom Šumskih Fitocenoza Jugoslavije, 2nd ed.; Svjetolost, Zavod za udžbenike i nastavna sredstva: Sarajevo, Bosnia and Herzegovina, 1986. [Google Scholar]
- Vukelić, J.; Baričević, D. The Association of Spreading Elm and Narrow-Leaved Ash (Fraxino-Ulmetum Laevis Slav. 1952) in Floodplain Forests of the Podravina and Podunavlje. Hacquetia 2004, 3, 49–60. [Google Scholar]
- Plišo Vusić, I.; Šapić, I.; Vukelić, J. Prepoznavanje i kartiranje šumskih staništa Natura 2000 u Hrvatskoj (II)–91F0, poplavne šume s vrstama Quercus robur, Ulmus laevis, Ulmus minor, Fraxinus angustifolia; 91L0, hrastovo-grabove šume ilirskoga područja. Šumar. List. 2019, 143, 461–467. [Google Scholar] [CrossRef]
- Horvat, I. Pregled Šumske Vegetacije u Hrvatskoj. Šumar. List 1937, 61, 337–344. [Google Scholar]
- Dekanić, I. Utjecaj podzemne vode na pridolazak i uspijevanje šumskog drveća u posavskim šumama kod Lipovljana. Glas. Šum. Pokuse 1962, 15, 5–117. [Google Scholar]
- Caudullo, G.; Houston Durrant, T. Fraxinus Angustifolia in Europe: Distribution, Habitat, Usage and Threats. In European atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, UK, 2016; Volume 97. [Google Scholar]
- Douda, J. Formalized Classification of the Vegetation of Alder Carr and Floodplain Forests in the Czech Republic. Preslia 2008, 80, 199–224. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Casavecchia, S.; Galdenzi, D.; Gasparri, R.; Pesaresi, S.; Poldini, L.; Sburlino, G.; Vagge, I.; Venanzoni, R. New Syntaxonomic Contribution to the Vegetation Prodrome of Italy. Plant Biosyst. 2015, 149, 603–615. [Google Scholar] [CrossRef]
- Gennai, M.; Gabellini, A.; Viciani, D.; Venanzoni, R.; Dell’Olmo, L.; Giunti, M.; Lucchesi, F.; Monacci, F.; Mugnai, M.; Foggi, B. The Floodplain Woods of Tuscany: Towards a Phytosociological Synthesis. Plant Soc. 2021, 58, 1–28. [Google Scholar] [CrossRef]
- Glavač, V. O šumi poljskog jasena sa kasnim drijemovcem (Leucoieto-Fraxinetum angustifoliae ass. nov.). Šumar. List 1959, 1–3, 39–45. [Google Scholar]
- Petrášová-Šibíková, M.; Bacigál, T.; Jarolímek, I. Fragmentation of Hardwood Floodplain Forests—How Does It Affect Species Composition? Community Ecol. 2017, 18, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Petrášová, M.; Jarolímek, I. Hardwood Floodplain Forests in Slovakia: Syntaxonomical Revision. Biologia 2012, 67, 889–908. [Google Scholar] [CrossRef]
- Poldini, L.; Sburlino, G.; Vidali, M. New Syntaxonomic Contribution to the Vegetation Prodrome of Italy. Plant Biosyst. 2017, 151, 1111–1119. [Google Scholar] [CrossRef]
- Rauš, D. Fitocenoloska Osnova i Vegetacijska Karta Nizinskih Suma Srednje Hrvatske. Glas. Šum. Pokuse 1993, 29, 335–364. [Google Scholar]
- Rauš, Đ. Nizinske Šume Pokupskog Bazena. Šumar. Inst. Jastrebar. Rad. 1996, 31, 17–37. [Google Scholar]
- Sokołowski, W.A. Zespół Carici Elongatae-Quercetum–Dębniak Turzycowy. Acta Soc. Bot. Pol. 1972, 41, 113–120. [Google Scholar] [CrossRef]
- Martinčič, A. Fragmenti Visokega Barja Na Ljubljanskem Barju. Scopolia 1987, 14, 1–53. [Google Scholar]
- Šilc, U. Asociacija Salicetum Cinereae Zölyomi 1931 v JV Sloveniji. Hacquetia 2002, 1, 165–184. [Google Scholar]
- Milanović, Đ.; Stupar, V. Checklist of Vegetation Classes of Bosnia and Herzegovina: How Much Do We Know? Rad. Šumar. Fak. Univ. Sarajev. 2019, 49, 9–20. [Google Scholar] [CrossRef]
- Slezák, M.; Hrivnák, R.; Petrášová, A.; Dítě, D. Variability of Alder-Dominated Forest Vegetation along a Latitudinal Gradient in Slovakia. Acta Soc. Bot. Pol. 2013, 81, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Cambridge University Press: Cambridge, UK, 1968. [Google Scholar]




| Cluster Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| No. of Relevés | 90 | 37 | 83 | 281 | 178 | 226 | 121 | 14 | 56 |
| Salicion albae | |||||||||
| Salix alba | 9158.1 | 8 | 23 | 21 | 47 20.4 | 1 | 2 | . | 9 |
| Salix euxina | 36 33.3 | . | 13 | 14 | 10 | . | 5 | . | 2 |
| Salicion triandrae | |||||||||
| Salix triandra | 19 4 | 97 82 | 11 | 2 | 1 | . | . | . | 4 |
| Rorippa sylvestris | 6 | 49 56.5 | 8 | 1 | 2 | 1 | . | . | . |
| Echinocystis lobata | 39 24.2 | 59 44.7 | 5 | 2 | 11 | 5 | 3 | . | 7 |
| Phalaroides arundinacea | 56 25.6 | 78 44.3 | 34 7.6 | 7 | 15 | 4 | 19 | . | 7 |
| Agrostis stolonifera agg. | 34 10.9 | 70 41.7 | 18 | 3 | 19 | 8 | 2 | 29 | 12 |
| Calystegia sepium | 46 25.1 | 62 40.3 | 14 | 5 | 11 | 7 | 4 | . | 14 |
| Persicaria dubia | 12 | 43 36.7 | 29 20.4 | 6 | 2 | 1 | 2 | . | 2 |
| Rorippa amphibia | 10 | 30 35.3 | 2 | . | 1 | 6 | 3 | . | 2 |
| Bidens tripartitus | 14 | 43 30.9 | 4 | 4 | 4 | 26 12.7 | 12 | . | 14 |
| Salicion eleagno-daphnoidis | |||||||||
| Salix eleagnos | 4 | . | 70 70.4 | 14 | 2 | . | . | . | . |
| Salix purpurea | 18 | 16 | 84 62.9 | 9 | 8 | . | 5 | . | 14 |
| Petasites hybridus | 1 | . | 53 50.8 | 31 24.4 | 2 | . | 1 | . | 2 |
| Saponaria officinalis | 3 | . | 31 46.4 | 2 | 3 | . | . | . | . |
| Clematis vitalba | 3 | . | 43 37.8 | 27 19.4 | 17 | 1 | 2 | . | . |
| Taraxacum sect. Taraxacum | 8 | 16 | 42 37.7 | 6 | 8 | 4 | 1 | . | 5 |
| Galium mollugo | 8 | 11 | 37 35.5 | 6 | 7 | 1 | 4 | . | 5 |
| Chaerophyllum hirsutum | 3 | . | 35 30.2 | 30 23.8 | 2 | . | 12 | . | 5 |
| Alnion incanae | |||||||||
| Corylus avellana | 2 | . | 27 | 63 44.8 | 20 | 19 | 15 | . | . |
| Acer pseudoplatanus | . | . | 35 26.1 | 50 42.6 | 3 | 1 | 13 | . | 2 |
| Lamium galeobdolon agg. | 3 | . | 11 | 49 42.4 | 24 | 8 | 6 | . | . |
| Sambucus nigra | 30 | 3 | 12 | 74 38.9 | 61 28.4 | 21 | 21 | . | 12 |
| Acer campestre | 2 | . | 12 | 57 38.2 | 21 | 48 30 | 9 | . | . |
| Fraxinus excelsior | 2 | . | 39 25.9 | 47 35 | 8 | 2 | 20 | . | 4 |
| Brachypodium sylvaticum | 13 | . | 53 25.7 | 64 34.8 | 46 19.8 | 8 | 16 | . | 4 |
| Symphytum tuberosum agg. | 3 | . | 4 | 35 34.2 | 27 24.1 | 1 | 4 | . | . |
| Geum urbanum | 2 | . | 7 | 47 33.6 | 30 | 29 | 10 | . | . |
| Lamium orvala | . | . | 23 | 33 33 | 13 | . | 2 | . | . |
| Carex sylvatica | 2 | . | 5 | 35 31.8 | 17 | 15 | 8 | . | . |
| Aegopodium podagraria | 26 | 11 | 35 | 70 31.1 | 57 20.9 | 14 | 26 | 21 | 11 |
| Cluster 5 | |||||||||
| Populus alba | 7 | . | . | 1 | 38 47.5 | 8 | 1 | . | . |
| Ulmus laevis | 13 | . | 1 | 4 | 54 43.1 | 38 26 | 7 | . | . |
| Populus nigra | 22 | . | 33 20.2 | 7 | 54 42.6 | 2 | 1 | . | . |
| Prunus padus | 6 | . | 4 | 5 | 40 37.9 | 4 | 21 | . | 4 |
| Arum maculatum | 2 | . | 1 | 17 14.7 | 30 34.2 | 6 | . | . | 2 |
| Solidago gigantea | 16 | . | 18 | 14 | 44 33.9 | 7 | 11 | . | 2 |
| Acer negundo | 23 22.4 | . | 2 | 3 | 30 32.1 | 4 | 1 | . | . |
| Galium aparine | 56 26.4 | 14 | 16 | 27 | 62 32 | 23 | 13 | . | 4 |
| Pulmonaria officinalis | 1 | . | 10 | 34 27.5 | 37 30.9 | 8 | 2 | . | 2 |
| Alno-Quercion | |||||||||
| Fraxinus angustifolia s.lat. | 7 | . | 1 | 9 | 43 19.9 | 84 55.3 | 37 | . | 4 |
| Ulmus minor | 4 | . | 1 | 10 | 14 | 52 47.3 | 15 | . | . |
| Quercus robur | 1 | . | 4 | 11 | 37 | 79 46.5 | 33 | 36 | 9 |
| Crataegus laevigata | 1 | . | 1 | 9 | 3 | 39 42.6 | 14 | . | . |
| Acer tataricum | 2 | . | . | 8 | 1 | 31 40.4 | 5 | . | . |
| Rumex sanguineus | 10 | . | 2 | 12 | 12 | 42 37.2 | 8 | . | 5 |
| Carex remota | 14 | . | . | 33 | 17 | 61 35.6 | 26 | 29 | 2 |
| Glechoma hederacea | 44 | 11 | 7 | 31 | 37 | 65 34.5 | 11 | . | 9 |
| Stachys palustris | 11 | 5 | 1 | 2 | 10 | 46 34.3 | 3422 | . | 7 |
| Lysimachia nummularia | 24 | 5 | 5 | 23 | 24 | 62 30.7 | 27 | 21 | 27 |
| Alnion glutinosae | |||||||||
| Carex elongata | . | . | . | 1 | . | 20 12 | 55 53.1 | . | 12 |
| Carex vesicaria | 7 | . | . | 2 | 3 | 11 | 38 39.3 | . | 11 |
| Peucedanum palustre | . | . | . | 6 | 2 | 23 8.1 | 52 36.2 | 36 | 18 |
| Carex riparia | 3 | . | . | 2 | 3 | 24 20.8 | 36 36 | . | 4 |
| Valeriana dioica s.lat. | 1 | . | 4 | 10 | 5 | 16 | 50 35.4 | 21 | 25 |
| Lycopus europaeus | 14 | 8 | 6 | 24 | 8 | 46 16.3 | 65 32.3 | . | 57 25.7 |
| Betulion pubescentis | |||||||||
| Betula pubescens | . | . | . | . | . | 1 | . | 86 91.5 | . |
| Molinia caerulea agg. | . | 5 | 11 | 2 | . | 1 | 7 | 93 81.8 | 4 |
| Pinus sylvestris | . | . | 4 | . | . | . | . | 71 80.7 | . |
| Sorbus aucuparia | . | . | . | 3 | . | . | 2 | 57 70.2 | . |
| Betula pendula | . | . | . | 1 | . | 1 | 1 | 50 66.5 | . |
| Salix aurita | . | . | . | 1 | . | 1 | 2 | 43 61.2 | . |
| Knautia sarajevensis | . | . | . | . | . | . | . | 36 57.5 | . |
| Calamagrostis villosa | . | . | . | . | . | . | . | 36 57.5 | . |
| Carex rostrata | . | . | . | . | . | . | . | 36 57.5 | . |
| Frangula alnus | 1 | . | 20 | 21 | 7 | 41 7.6 | 66 26.7 | 100 52.5 | 23 |
| Epilobium palustre | . | 5 | . | 1 | 1 | . | 2 | 36 50.3 | . |
| Rubus hirtus s.lat. | . | . | 1 | 14 8.8 | . | 4 | 1 | 43 49.1 | 2 |
| Salicion cinereae | |||||||||
| Salix cinerea | 3 | 8 | 4 | 2 | . | 8 | 28 6.3 | 36 | 100 68.8 |
| Filipendula ulmaria | 14 | . | 8 | 26 | 12 | 8 | 45 23.9 | . | 55 32.8 |
| Diagnostic species for more than one cluster | |||||||||
| Alnus glutinosa agg. | 12 | . | 11 | 81 37.6 | 31 | 47 | 94 47.8 | . | 7 |
| Other species with high frequency | |||||||||
| Urtica dioica | 82 21 | 86 24 | 48 | 59 | 66 | 61 | 43 | . | 27 |
| Rubus caesius | 73 | 19 | 58 | 70 | 90 29.5 | 69 | 46 | . | 12 |
| Solanum dulcamara | 38 | 81 29.6 | 27 | 22 | 13 | 38 | 54 9.9 | 29 | 59 13.7 |
| Galium palustre agg. | 29 | 49 | 6 | 8 | 10 | 58 16.1 | 60 17.3 | 36 | 70 24.7 |
| Ranunculus repens | 23 | 35 | 16 | 37 | 16 | 48 10.6 | 49 11.3 | 14 | 6423 |
| Cornus sanguinea | 43 | . | 43 | 65 24.7 | 64 24.2 | 37 | 22 | . | 14 |
| Lysimachia vulgaris | 28 | 19 | 8 | 16 | 10 | 33 | 67 28 | 43 | 50 15 |
| Angelica sylvestris | 24 | 11 | 46 14.8 | 37 8.1 | 12 | 10 | 31 | 29 | 45 |
| Lythrum salicaria | 27 | 57 23.9 | 18 | 6 | 7 | 31 3.6 | 44 13.5 | . | 52 19.9 |
| Iris pseudacorus | 43 | 14 | 4 | 6 | 29 | 54 22.3 | 52 20.4 | . | 38 |
| Viburnum opulus | 10 | . | 13 | 43 13.1 | 35 | 25 | 53 21 | 36 | 25 |
| Euonymus europaeus | 32 | 3 | 12 | 55 25.2 | 47 18.9 | 31 | 26 | . | 12 |
| Persicaria hydropiper | 28 | 57 28.9 | 5 | 15 | 10 | 50 22.8 | 31 | . | 9 |
| Caltha palustris | 7 | . | 11 | 30 | 8 | 31 | 48 21.5 | 36 | 34 |
| Humulus lupulus | 43 17.4 | 27 | 6 | 32 | 39 13.6 | 10 | 26 | . | 21 |
| Poa trivialis | 43 19.8 | 14 | 22 | 20 | 38 14.8 | 21 | 11 | . | 18 |
| Deschampsia cespitosa | 4 | . | 40 17.1 | 29 7.5 | 13 | 22 | 29 | 36 | 11 |
| Crataegus monogyna | 7 | . | 22 | 48 26.1 | 34 | 46 24.4 | 14 | . | 2 |
| Myosotis palustris agg. | 12 | 30 | 4 | 10 | 2 | 35 15.8 | 40 20.4 | 7 | 23 |
| Circaea lutetiana | 9 | . | . | 44 25.7 | 35 | 38 20.3 | 22 | . 2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koljanin, D.; Brujić, J.; Čarni, A.; Milanović, Đ.; Škvorc, Ž.; Stupar, V. Classification of Wetland Forests and Scrub in the Western Balkans. Diversity 2023, 15, 370. https://doi.org/10.3390/d15030370
Koljanin D, Brujić J, Čarni A, Milanović Đ, Škvorc Ž, Stupar V. Classification of Wetland Forests and Scrub in the Western Balkans. Diversity. 2023; 15(3):370. https://doi.org/10.3390/d15030370
Chicago/Turabian StyleKoljanin, Dragan, Jugoslav Brujić, Andraž Čarni, Đorđije Milanović, Željko Škvorc, and Vladimir Stupar. 2023. "Classification of Wetland Forests and Scrub in the Western Balkans" Diversity 15, no. 3: 370. https://doi.org/10.3390/d15030370
APA StyleKoljanin, D., Brujić, J., Čarni, A., Milanović, Đ., Škvorc, Ž., & Stupar, V. (2023). Classification of Wetland Forests and Scrub in the Western Balkans. Diversity, 15(3), 370. https://doi.org/10.3390/d15030370