
Variation in Leaf Pigment Complex Traits of Wetland Plants Is Related to Taxonomy and Life Forms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Plant Material
2.2. Plant Sizes and Canopy Parameters
2.3. Leaf Traits
2.4. Leaf Pigment Content and CO2 Uptake Rate
2.5. Data Analysis
3. Results
3.1. Variation of Leaf Pigment Complex Traits across Wetland Plant Species
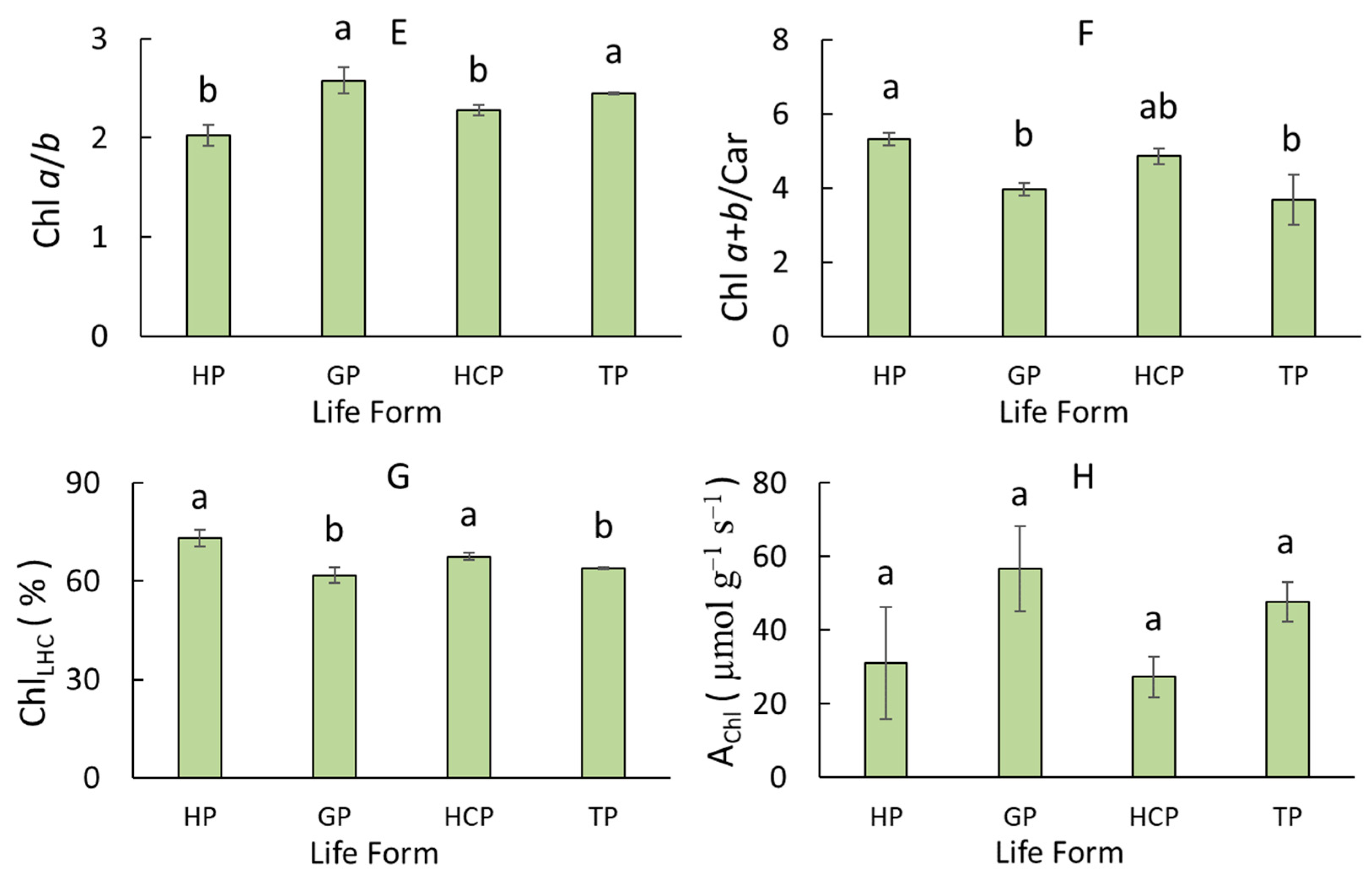
3.2. Variation of Leaf Pigment Complex Traits in Dicots of Different Life Forms
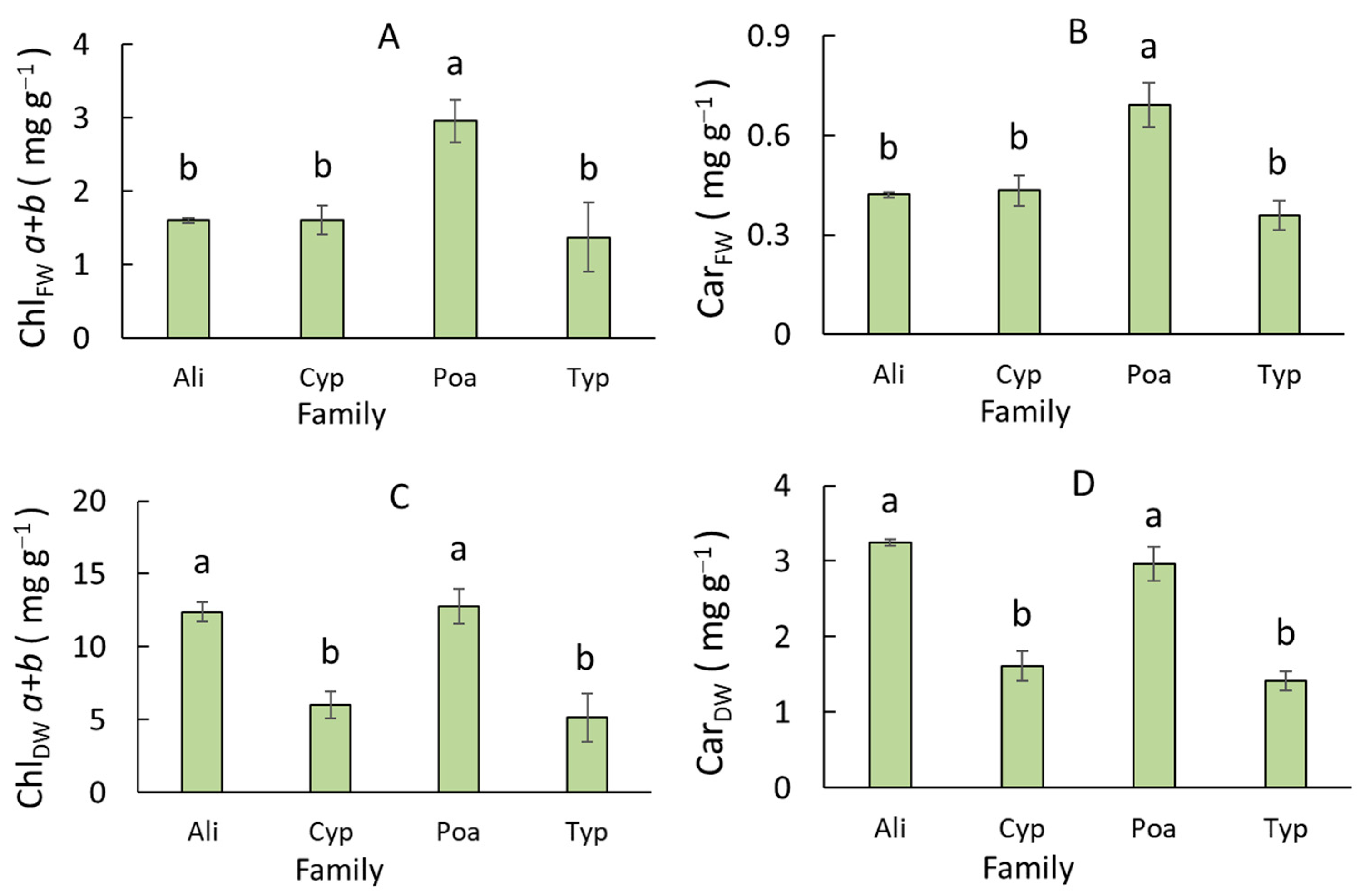
3.3. Variation of Leaf Pigment Complex Traits in Monocots of Different Families
3.4. Relationships between Leaf Pigment Complex Parameters, Leaf Traits, and Plant Productivity
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernandez-Marin, B.; Hernandez, A.; Garcia-Plazaola, J.I. Internal and external factors affecting photosynthetic pigment composition in plants: A meta-analytical approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytol. 2006, 172, 11–21. [Google Scholar] [CrossRef]
- Gao, J.; Wang, H.; Yuan, Q.; Feng, Y. Structure and Function of the Photosystem Supercomplexes. Front. Plant Sci. 2018, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Croce, R. Chlorophyll-binding proteins of higher plants and cyanobacteria. In Photosynthesis: Plastid Biology, Energy Conversion and Carbon Assimilation, Advances in Photosynthesis and Respiration; Eaton-Rye, J.J., Tripathy, B.C., Sharkey, T.D., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 127–149. [Google Scholar]
- Cazzaniga, S.; Li, Z.; Niyogi, K.K.; Bassi, R.; Dall’Osto, L. The Arabidopsis szl1 mutant reveals a critical role of b-carotene in photosystem I photoprotection. Plant Physiol. 2012, 159, 1745–1758. [Google Scholar] [CrossRef] [Green Version]
- Ballottari, M.; Dall’Osto, L.; Morosinotto, T.; Bassi, R. Contrasting behavior of higher plant photosystem I and II antenna systems during acclimation. J. Biol. Chem. 2007, 282, 8947–8958. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef]
- Rossatto, D.R.; de Araújo, P.E.; da Silva, B.H.P.; Franco, A.C. Photosynthetic responses of understory savanna plants: Implications for plant persistence in savannas under tree encroachment. Flora 2018, 240, 34–38. [Google Scholar] [CrossRef]
- Lippert, I.; Rolletschek, H.; Kohl, J.-G. Photosynthetic pigments and efficiencies of two Phragmites australis stands in different nitrogen availabilities. Aquat. Bot. 2001, 69, 359–365. [Google Scholar] [CrossRef]
- Ronzhina, D.A.; Nekrasova, G.F.; P’yankov, V.I. Comparative characterization of the pigment complex in emergent, floating, and submerged leaves of hydrophytes. Rus. J. Plant Physiol. 2004, 51, 21–27. [Google Scholar] [CrossRef]
- Slemnev, N.N.; Sheremet’ev, S.N.; Maslova, T.G.; Tsoozh, S.; Altantsoozh, A. Variety of photosynthetic plant apparatus of Mongolia: Analysis of biological, ecological and evolutionary series. Bot. Zhurnal 2012, 97, 1377–1396. (In Russian) [Google Scholar]
- Houborg, R.; Cescatti, A.; Migliavacca, M.; Kustas, W. Satellite retrievals of leaf chlorophyll and photosynthetic capacity for improved modeling of GPP. Agric. For. Meteorol. 2013, 177, 10–23. [Google Scholar] [CrossRef]
- Dymova, O.; Dalke, I.; Golovko, T. Pigment characteristics of the plants of northern ecosystems and their correlation with photosynthetic activity. In Photosynthetic Pigments—Chemical Structure, Biological Function and Ecology; Golovko, T.K., Gruszeski, W.I., Prasad, M.N.V., Strzalka, K., Eds.; Komi Scientific Centre of the Ural Branch of the Russian Academy of Sciences: Syktyvkar, Russia, 2014; pp. 221–236. [Google Scholar]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Grubb, P.J.; Marañón, T. The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain. Plant Ecol. 2003, 168, 139–163. [Google Scholar] [CrossRef]
- Ivanov, L.A.; Ronzhina, D.A.; Ivanova, L.A. Changes in leaf characteristics as indicator of the alteration of functional types of steppe plants along the aridity gradient. Russ. J. Plant Physiol. 2008, 55, 301–307. [Google Scholar] [CrossRef]
- Zunzunegui, M.; Diaz-Barradas, M.C.; Jauregui, J.; Rodriguez, H.; Alvarez-Cansino, L. Season-dependent and independent responses of Mediterranean scrub to light conditions. Plant Physiol. Biochem. 2016, 102, 80–91. [Google Scholar] [CrossRef]
- Li, Y.; Liu, C.; Zhang, J.; Yang, H.; Xu, L.; Wang, Q.; Sack, L.; Wu, X.; Hou, J.; He, N. Variation in leaf chlorophyll concentration from tropical to cold-temperate forests: Association with gross primary productivity. Ecol. Ind. 2018, 85, 383–389. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Zhang, J.; Baird, A.S.; He, N. Optimal community assembly related to leaf economic- hydraulic-anatomical traits. Front. Plant Sci. 2020, 11, 341. [Google Scholar] [CrossRef]
- Golovko, T.; Dymova, O.; Yatsco, Y.; Tabalenkova, G. Photosynthetic pigments apparatus in the northern plants. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, M., Ed.; Marcel Dekker Inc.: New York, NY, USA, 2011; pp. 391–405. [Google Scholar]
- Dymova, O.; Golovko, T. Photosynthetic pigments in native plants of the taiga zone at the European Northeast Russia. Russ. J. Plant Physiol. 2019, 66, 384–392. [Google Scholar] [CrossRef]
- Maslova, T.G.; Popova, I.A. Adaptive properties of the pigment systems. Photosynthetica 1993, 29, 195–203. [Google Scholar]
- Voronin, P.Y.; Ivanova, L.A.; Ronzhina, D.A.; Ivanov, L.A.; Anenkhonov, O.A.; Black, C.C.; Gunin, P.D.; P’yankov, V.I. Structural and functional changes in the leaves of plants from steppe communities as affected by aridization of the Eurasian climate. Russ. J. Plant Physiol. 2003, 50, 604–611. [Google Scholar] [CrossRef]
- Ivanova, L.A.; Yudina, P.K.; Ronzhina, D.A.; Ivanov, L.A.; Hölzel, N. Quantitative mesophyll parameters rather than whole-leaf traits predict response of C3 steppe plants to aridity. New Phytol. 2018, 217, 558–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, L.A.; Ivanova, L.A.; Ronzhina, D.A.; Yudina, P.K. Changes in the chlorophyll and carotenoid contents in the leaves of steppe plants along latitudinal gradient in South Ural. Russ. J. Plant Physiol. 2013, 60, 812–820. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Dymova, O.V.; Dalke, I.V. Photosynthetic pigments and CO2-exchange of macrophytes in the middle taiga subzone. Proc. Komi Sci. Centre Ural Branch Russ. Acad. Sci. 2016, 25, 37–44. (In Russian) [Google Scholar]
- Gorchakovskii, P.L.; Shurova, E.A.; Knyazev, M.S. (Eds.) Opredelitel’ Sosudistykh Rastenii Srednego Urala (Identifier of Vascular Plants in the Middle Urals); Nauka: Moscow, Russia, 1994; 525p. (In Russian) [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford at the Clarendon Press: Oxford, UK, 1934; 632p. [Google Scholar]
- Kulikov, P.V. Opredelitel’ Sosudistykh Rrastenii Chelyabinskoi Oblasti (Synopsis of the Flora of the Chelyabinsk Region (Vascular Plants); Geotour: Yekaterinburg–Miass, Russia, 2005; 537p. (In Russian) [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–317. [Google Scholar] [CrossRef]
- Sofronova, V.E.; Chepalov, V.A.; Dymova, O.V.; Golovko, T.K. The role of pigment system of an evergreen dwarf shrub Ephedra monosperma in adaptation to the climate of Central Yakutia. Russ. J. Plant Physiol. 2014, 61, 246–254. [Google Scholar] [CrossRef]
- Dymova, O.V.; Golovko, T.K. Pigment apparatus in Ajuga reptans plants as affected by adaptation to light growth conditions. Russ. J. Plant Physiol. 2007, 54, 39–45. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Chen, F.; Zhao, G.; Shen, Y.; Li, Z.; Tan, B.; Zhu, H.; Wang, Q.; Fu, X. Shading gradients drive differential responses of leaf traits in an early community germinated by forest topsoil. Diversity 2022, 14, 600. [Google Scholar] [CrossRef]
- Ivanov, L.A.; Ronzhina, D.A.; Yudina, P.K.; Kalashnikova, I.V.; Ivanova, L.A.; Zolotareva, N.V. Seasonal dynamics of the chlorophyll and carotenoid content in the leaves of steppe and forest plants on species and community level. Russ. J. Plant Physiol. 2020, 67, 453–462. [Google Scholar] [CrossRef]
- Valladares, F.; Martinez-Ferri, E.; Balaguer, L.; Perez-Corona, E.; Manrique, E. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource-use strategy. New Phytol. 2000, 148, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Ronzhina, D.A. Ecological differentiation between invasive and native species of the genus Epilobium in riparian ecosystems is associated with plant functional traits. Russ. J. Biol. Invas. 2020, 11, 132–142. [Google Scholar] [CrossRef]
- Chemeris, E.V. Rastitel’nyi Pokrov Istokovykh Vetlandov Verkhnego Povolzh’ya (Vegetation Cover of the Upper Volga Region Source Wetlands); Rybinskii Dom Pechati: Rybinsk, Russia, 2004; 158p. (In Russian) [Google Scholar]
- Luo, X.; Croft, H.; Chen, J.M.; He, L.; Keenan, T.F. Improved estimates of global terrestrial photosynthesis using information on leaf chlorophyll content. Glob. Chang. Biol. 2019, 25, 2499–2514. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, N.; Yu, G. Opposing shifts in distributions of chlorophyll concentration and composition in grassland under warming. Sci. Rep. 2021, 11, 15736. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Mean | SE | Minimum | Maximum | CV | |
|---|---|---|---|---|---|---|
| ChlFW a+b, mg g−1 | Dicots | 1.95a | 0.10 | 1.33 | 2.66 | 0.23 |
| Monocots | 1.87a | 0.18 | 0.44 | 4.38 | 0.49 | |
| Total | 1.90 | 0.11 | 0.44 | 4.38 | 0.39 | |
| CarFW, mg g−1 | Dicots | 0.44a | 0.03 | 0.25 | 0.84 | 0.29 |
| Monocots | 0.47a | 0.04 | 0.22 | 0.99 | 0.42 | |
| Total | 0.45 | 0.03 | 0.22 | 0.99 | 0.37 | |
| ChlDW a+b, mg g−1 | Dicots | 11.79a | 1.13 | 5.23 | 21.91 | 0.42 |
| Monocots | 8.56b | 0.85 | 2.20 | 18.51 | 0.50 | |
| Total | 9.95 | 0.72 | 2.20 | 21.91 | 0.48 | |
| CarDW, mg g−1 | Dicots | 2.56a | 0.22 | 1.27 | 4.80 | 0.37 |
| Monocots | 2.14a | 0.19 | 0.79 | 3.82 | 0.44 | |
| Total | 2.32 | 0.14 | 0.79 | 4.80 | 0.41 | |
| ChlArea a+b, µg cm−2 | Dicots | 32.82a | 1.50 | 20.81 | 46.53 | 0.20 |
| Monocots | 40.25a | 3.42 | 11.67 | 81.77 | 0.42 | |
| Total | 37.04 | 2.10 | 11.67 | 81.77 | 0.38 | |
| CarArea, µg cm−2 | Dicots | 7.41b | 0.46 | 3.35 | 11.50 | 0.27 |
| Monocots | 10.05a | 0.62 | 5.18 | 17.37 | 0.31 | |
| Total | 8.91 | 0.45 | 3.35 | 17.37 | 0.33 | |
| Chl a/b | Dicots | 2.34b | 0.05 | 1.92 | 2.83 | 0.10 |
| Monocots | 2.53a | 0.03 | 2.22 | 2.99 | 0.07 | |
| Total | 2.44 | 0.03 | 1.92 | 2.99 | 0.09 | |
| Chl a+b/Car | Dicots | 4.58a | 0.18 | 3.01 | 6.35 | 0.17 |
| Monocots | 3.94b | 0.18 | 1.63 | 5.46 | 0.23 | |
| Total | 4.22 | 0.13 | 1.63 | 6.35 | 0.21 | |
| ChlLHC, % | Dicots | 66.35a | 1.07 | 57.42 | 75.52 | 0.07 |
| Monocots | 62.56b | 0.60 | 55.20 | 68.57 | 0.05 | |
| Total | 64.20 | 0.63 | 55.20 | 75.52 | 0.07 | |
| AChl, µmol CO2 g−1 Chl s−1 | Dicots | 36.67a | 4.74 | 11.05 | 84.13 | 0.56 |
| Monocots | 46.82a | 3.80 | 16.64 | 94.86 | 0.41 | |
| Total | 42.44 | 3.04 | 11.05 | 94.86 | 0.47 |
| Traits | Dicots | Monocots | ||
|---|---|---|---|---|
| Life Form n = 18 df = 3, 14 | Family n = 13 df = 5, 7 | Life Form n = 24 df = 2, 21 | Family n = 20 df = 3, 16 | |
| ChlFW a+b, mg g−1 | 0.88 ns | 1.20 ns | 0.57 ns | 6.80 ** |
| CarFW, mg g−1 | 2.60 ns | 0.68 ns | 0.60 ns | 6.09 ** |
| ChlDW a+b, mg g−1 | 6.17 ** | 0.75 ns | 0.55 ns | 9.99 ** |
| CarDW, mg g−1 | 7.15 ** | 0.83 ns | 0.09 ns | 13.35 *** |
| ChlArea a+b, µg cm−2 | 0.04 ns | 0.61 ns | 0.36 ns | 1.39 ns |
| CarArea, µg cm−2 | 1.67 ns | 0.85 ns | 0.18 ns | 1.86 ns |
| Chl a/b | 4.55 * | 2.24 ns | 1.00 ns | 2.34 ns |
| Chl a+b/Car | 4.15 * | 1.46 ns | 1.28 ns | 0.75 ns |
| ChlLHC, % | 4.63 * | 1.93 ns | 0.95 ns | 2.14 ns |
| AChl, µmol CO2 g−1 Chl s−1 | 2.66 ns | 2.97 ns | 0.24 ns | 0.66 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronzhina, D.A. Variation in Leaf Pigment Complex Traits of Wetland Plants Is Related to Taxonomy and Life Forms. Diversity 2023, 15, 372. https://doi.org/10.3390/d15030372
Ronzhina DA. Variation in Leaf Pigment Complex Traits of Wetland Plants Is Related to Taxonomy and Life Forms. Diversity. 2023; 15(3):372. https://doi.org/10.3390/d15030372
Chicago/Turabian StyleRonzhina, Dina A. 2023. "Variation in Leaf Pigment Complex Traits of Wetland Plants Is Related to Taxonomy and Life Forms" Diversity 15, no. 3: 372. https://doi.org/10.3390/d15030372
APA StyleRonzhina, D. A. (2023). Variation in Leaf Pigment Complex Traits of Wetland Plants Is Related to Taxonomy and Life Forms. Diversity, 15(3), 372. https://doi.org/10.3390/d15030372