
Fantastic Flatworms and Where to Find Them: Insights into Intertidal Polyclad Flatworm Distribution in Southeastern Australian Boulder Beaches

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
Statistical Analyses
3. Results
3.1. Species of Flatworms Identified
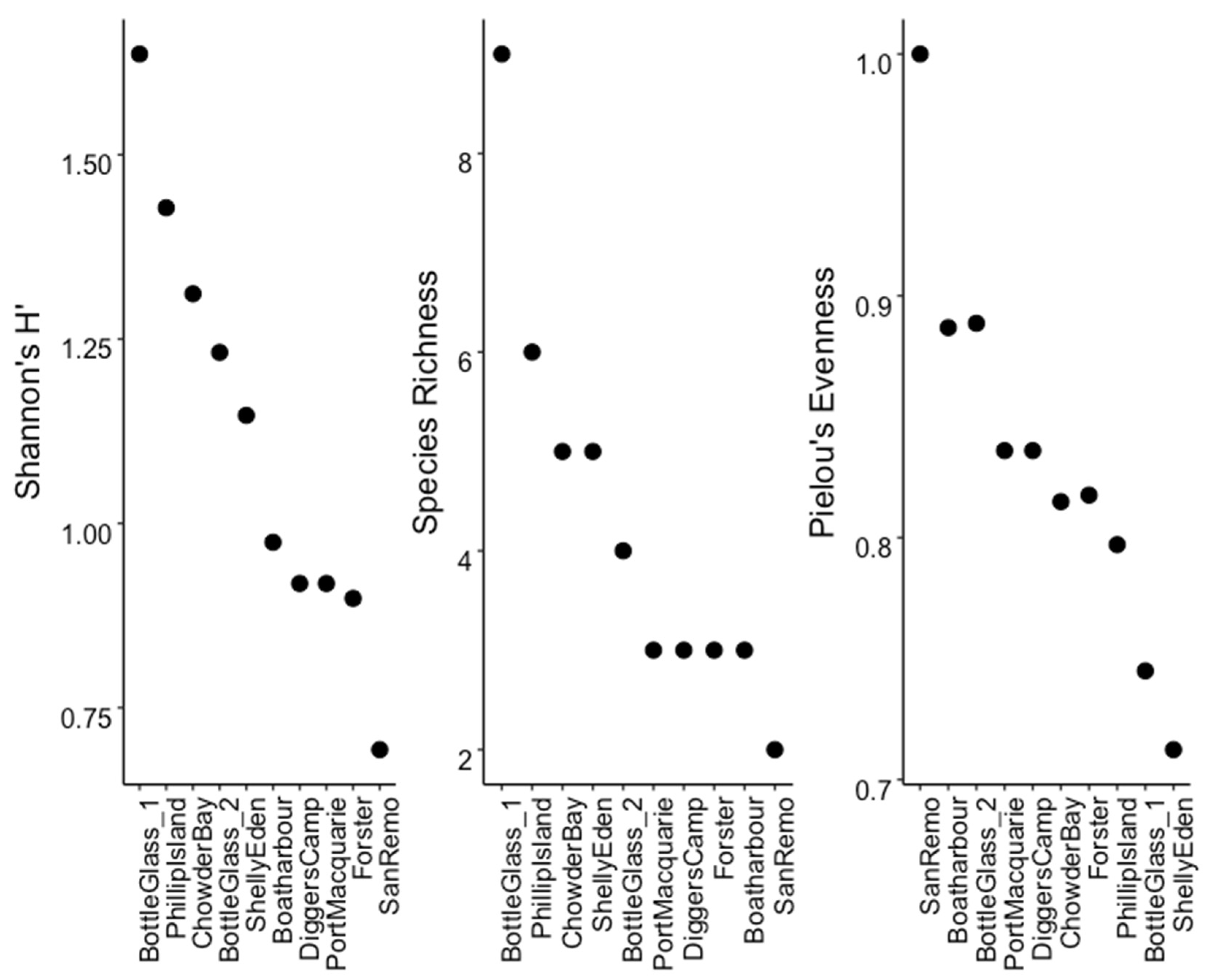
3.2. Hotspots of Abundance and Biodiversity
3.3. Influence of Boulder Size and Beach Exposure on Flatworm Diversity
3.4. Relationship between Rock Size and Flatworm Abundance
3.5. Influence of Beach Exposure on Preferred Boulder Size for Flatworms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sala, E.; Knowlton, N. Global marine biodiversity trends. Annu. Rev. Environ. Resour. 2006, 31, 93–122. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.J.; Rees, T.; Beesley, P.; Bax, N.J. Marine biodiversity in the Australian region. PLoS ONE 2010, 5, e11831. [Google Scholar] [CrossRef] [Green Version]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- McCauley, D.J.; Pinsky, M.L.; Palumbi, S.R.; Estes, J.A.; Joyce, F.H.; Warner, R.R. Marine defaunation: Animal loss in the global ocean. Science 2015, 347, 6219. [Google Scholar] [CrossRef] [Green Version]
- McClenachan, L.; Ferretti, F.; Baum, J.K. From archives to conservation: Why historical data are needed to set baselines for marine animals and ecosystems. Conserv. Lett. 2012, 5, 349–359. [Google Scholar] [CrossRef]
- Hutchings, P.A. Potential loss of biodiversity and the critical importance of taxonomy—An Australian perspective. Adv. Mar. Biol 2021, 88, 3–16. [Google Scholar]
- Sousa, W.P. Disturbance in marine intertidal boulder fields: The nonequilibrium maintenance of species diversity. Ecology 1979, 60, 1225–1239. [Google Scholar] [CrossRef] [Green Version]
- McGuinness, K.A. Disturbance and organisms on boulders. II. Causes of patterns in diversity and abundance. Oecologia 1987, 71, 420–430. [Google Scholar] [CrossRef]
- Chapman, M.G. Molluscs and echinoderms under boulders: Tests of generality of patterns of occurrence. J. Exp. Mar. Biol. Ecol. 2005, 325, 65–83. [Google Scholar] [CrossRef]
- Chapman, M.G. Intertidal boulder-fields: A much neglected, but ecologically important, intertidal habitat. Oceanogr. Mar. Biol. Ann. Rev. 2017, 55, 35–53. [Google Scholar]
- Chapman, M.G.; Underwood, A.J.; Browne, M.A. An assessment of the current usage of ecological engineering and reconciliation ecology in managing alterations to habitats in urban estuaries. Ecol. Eng. 2018, 120, 560–573. [Google Scholar] [CrossRef]
- Liversage, K.; Kotta, J. Unveiling commonalities in understudied habitats of boulder-reefs: Life-history traits of the widespread invertebrate and algal inhabitants. Mar. Biol. Res. 2018, 14, 655–671. [Google Scholar] [CrossRef]
- Smith, K.A.; Otway, N.M. Spatial and temporal patterns of abundance and the effects of disturbance on under-boulder chitons. Molluscan Res. 2013, 18, 43–57. [Google Scholar] [CrossRef]
- Liversage, K.; Cole, V.J.; McQuaid, C.D.; Coleman, R.A. Intercontinental tests of the effects of habitat patch type on the distribution of chitons within and among patches in intertidal boulder field landscapes. Mar. Biol. 2012, 159, 2777–2786. [Google Scholar] [CrossRef]
- Liversage, K.; Cole, V.; Coleman, R.; McQuaid, C. Availability of microhabitats explains a widespread pattern and informs theory on ecological engineering of boulder reefs. J. Exp. Mar. Biol. Ecol. 2017, 489, 36–42. [Google Scholar] [CrossRef]
- Hutchings, P.A.; Ahyong, S.T.; Ashcroft, M.B.; McGrouther, M.A.; Reid, A.L. Sydney Harbour: Its diverse biodiversity. Aust. Zool. 2013, 36, 257–320. [Google Scholar] [CrossRef]
- Liversage, K.; Benkendorff, K. A preliminary investigation of diversity, abundance, and distributional patterns of chitons in intertidal boulder fields of differing rock type in South Australia. Molluscan Res. 2013, 33, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Nimbs, M.; Willan, R.; Smith, S. Is Port Stephens, eastern Australia, a global hotspot for biodiversity of Aplysiidae (Gastropoda: Heterobranchia)? Molluscan Res. 2016, 37, 45–65. [Google Scholar] [CrossRef]
- Ahyong, S.T. Biogeography of Australian Marine Invertebrates. In Handbook of Australasian Biogeography; CRC Press and Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 81–99. [Google Scholar]
- Nimbs, M.J.; Davis, T.R.; Holmes, S.P.; Hill, L.; Wehmeyer, S.; Prior, A.; Williamson, J.E. The Taming of Smeagol? A New Population and an Assessment of the Known Population of the Critically Endangered Pulmonate Gastropod Smeagol hilaris (Heterobranchia, Otinidae). Diversity 2023, 15, 86. [Google Scholar] [CrossRef]
- Aldana, M.; Maturana, D.; Pulgar, J.; García-Huidobro, M.R. Predation and anthropogenic impact on community structure of boulder beaches. Sci. Mar. 2016, 80, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Merory, M.; Newman, L. A new stylochid flatworm (Platyhelminthes, Polycladida) from Victoria, Australia and observations on its biology. J. Nat. Hist. 2005, 39, 2581–2589. [Google Scholar] [CrossRef]
- Lee, K.-M.; Beal, M.A.; Johnston, E.L. A new predatory flatworm (Platyhelminthes, Polycladida) from Botany Bay, New South Wales, Australia. J. Nat. Hist. 2006, 39, 3987–3995. [Google Scholar] [CrossRef]
- Gammoudi, M.; Ahmed, R.B.; Bouriga, N.; Ben-Attia, M.; Harrath, A.H. Predation by the polyclad flatworm Imogine mediterranea on the cultivated mussel Mytilus galloprovincialis in Bizerta Lagoon (northern Tunisia). Aquac. Res. 2016, 1, 10. [Google Scholar] [CrossRef]
- Bolton, D.K.; Clark, G.F.; Johnston, E.L. Novel in situ predator exclusion method reveals the relative effects of macro and mesopredators on sessile invertebrates in the field. J. Exp. Mar. Biol. Ecol. 2019, 513, 13–20. [Google Scholar] [CrossRef]
- Rodríguez, J.; Hutchings, P.A.; Williamson, J.E. Biodiversity of intertidal marine flatworms (Polycladida, Platyhelminthes) in southeastern Australia. Zootaxa 2021, 5024, 1–63. [Google Scholar] [CrossRef]
- Newman, L.J.; Cannon, L.R. Marine Flatworms: The World of Polyclads; CSIRO Publishing: Collingwood, Australia, 2003; 97p. [Google Scholar]
- Holleman, J. Some New Zealand Polyclads (Platyhelminthes, Polycladida). Zootaxa 2007, 1560, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bahia, J.; Padula, V.; Delgado, M. Five new records and morphological data of polyclad species (Platyhelminthes: Turbellaria) from Rio Grande do Norte, Northeastern Brazil. Zootaxa 2012, 3170, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Sivaperuman, C.; Raghunathan, C. Three new records of polyclad flatworms from India. Mar. Biodivers. Rec. 2015, 8, E29. [Google Scholar] [CrossRef]
- Noreña, C.; Rodríguez, J.; Pérez, J.; Almon, B. New Acotylea (Polycladida, Platyhelminthes) from the east coast of the North Atlantic Ocean with special mention of the Iberian littoral. Zootaxa 2015, 4039, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Oya, Y.; Kajihara, H.A. New Species of Phaenoplana (Platyhelminthes: Polycladida) from the Ogasawara Islands. Species Divers 2019, 24, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Oak, H.L. The Boulder Beach: A Fundamentally Distinct Sedimentary Assemblage. Ann. Am. Assoc. Geogr. 1984, 74, 71–82. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J Geol. 1922, 30, 377. [Google Scholar] [CrossRef]
- Carter, R.W.G.; Orford, J.D. The morphodynamics of coarse clastic beaches and barriers: A short-and long-term perspective. J. Coast. Res. 1993, 15, 158–179. [Google Scholar]
- McKenna, J. Boulder Beaches. In Encyclopedia of Coastal Science; Encyclopedia of Earth Science Series; Schwartz, M.L., Ed.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar] [CrossRef]
- Baardseth, E.M. Square-scanning, two-stage sampling method of estimating seaweed quantities. Rep. Norw. Inst Seaweed Res. 1970, 33, 1–41. [Google Scholar]
- Wernberg, T.; Connell, S.D. Physical disturbance and subtidal habitat structure on open rocky coasts: Effects of wave exposure, extent and intensity. J. Sea Res. 2008, 59, 237–248. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. RStudio; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 10 January 2023).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 January 2023).
- Russell, V.; Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.7.2. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 10 January 2023).
- Johnston, E.L.; Mayer-Pinto, M.; Hutchings, P.A.; Marzinelli, E.M.; Ahyong, S.T.; Birch, G.; Booth, D.J.; Creese, R.G.; Doblin, M.A.; Figueira, W.; et al. Sydney Harbour: What we do and do not know about a highly diverse estuary. Mar. Freshw. Res. 2015, 66, 1073–1087. [Google Scholar] [CrossRef]
- Barton, J.; Humphrey, C.; Bourne, D.G.; Hutson, K.S. Biological controls to manage Acropora eating flatworms in coral aquaculture. Aquac. Environ. Interact. 2020, 12, 61–66. [Google Scholar] [CrossRef]
- Itoi, S.; Sato, T.; Takei, M.; Yamada, R.; Ogata, R.; Oyama, H.; Teranishi, S.; Kishiki, A.; Wada, T.; Noguchi, K.; et al. The planocerid flatworm is a main supplier of toxin to tetrodotoxin-bearing fish juveniles. Chemosphere 2020, 249, 126217. [Google Scholar] [CrossRef]
- Herrera, A.; Bustamante, R.H.; Shepherd, S. The fishery for endemic chitons in the Galapagos Islands. Not. Galápagos 2003, 62, 24–28. [Google Scholar]
- Rogers, C.N.; Williamson, J.E.; Carson, D.G.; Steinberg, P.D. Diel vertical movement by mesograzers on seaweeds. Mar. Ecol. Prog. Ser. 1998, 166, 301–306. [Google Scholar] [CrossRef]
- Shepherd, S.; Toral-Granda, V.; Edgar, G.J. Estimating the abundance of clustered and cryptic marine macro-invertebrates in the Galápagos with particular reference to sea cucumbers. Not. Galápagos 2003, 62, 36–39. [Google Scholar]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Dial, R.; Roughgarden, J. Theory of marine communities: The intermediate disturbance hypothesis. Ecology 1988, 79, 1412–1424. [Google Scholar] [CrossRef]
- Benkendorff, K.; Davis, A.R. Identifying hotspots of molluscan species richness on rocky intertidal reefs. Biodivers. Conserv. 2002, 11, 1959–1973. [Google Scholar] [CrossRef]
- Underwood, A.J.; Chapman, M.G. Spatial analyses of intertidal assemblages on sheltered rocky shores. Aust. J. Ecol. 1998, 23, 138–157. [Google Scholar] [CrossRef]
- Pepler, A.; Coutts-Smith, A.; Timbal, B. The role of East Coast Lows on rainfall patterns and inter-annual variability across the East Coast of Australia. Int. J. Climatol. 2014, 34, 1011–1021. [Google Scholar] [CrossRef]
- Corte, G.N.; Schlacher, T.A.; Checon, H.H.; Barboza, C.A.; Siegle, E.; Coleman, R.A.; Amaral, A.C.Z. Storm effects on intertidal invertebrates: Increased beta diversity of few individuals and species. PeerJ 2017, 5, e3360. [Google Scholar] [CrossRef] [Green Version]
- Mieszkowska, N.; Burrows, M.T.; Hawkins, S.J.; Sugden, H. Impacts of pervasive climate change and extreme events on rocky intertidal communities: Evidence from long-term data. Front. Mar. Sci. 2021, 8, 642764. [Google Scholar] [CrossRef]
- Lawrence, E.R.; Fraser, D.J. Latitudinal biodiversity gradients at three levels: Linking species richness, population richness and genetic diversity. Glob. Ecol. Biogeogr. 2020, 29, 770–788. [Google Scholar] [CrossRef] [Green Version]
- Galleni, L.; Tongiorgi, P.; Ferrero, E.; Salghetti, U. Stylochus mediterraneus (Turbellaria: Polycladida), predator on the mussel Mytilus galloprovincialis. Mar. Biol. 1980, 55, 317–326. [Google Scholar]
- Rawlinson, K.A.; Gillis, J.A.; Billings, R.E.; Borneman, E.H. Taxonomy and life history of the Acropora-eating polyclad flatworm: Amakusaplana acroporae nov. sp. (Polycladida, Prosthiostomidae). Coral Reefs 2011, 30, 693–705. [Google Scholar] [CrossRef]
- Prudhoe, S. Polyclad turbellarians from the southern coasts of Australia. Rec. Aust. Mus. 1982, 18, 361–384. [Google Scholar]
- Bennett, I.; Pope, E.C. Intertidal zonation of the exposed rocky shores of Victoria, together with a rearrangement of the biogeographical provinces of temperate Australian shores. Aust. J. Mar. Freshw. Res. 1960, 4, 105–159. [Google Scholar] [CrossRef]
- Rawlinson, K.A. The diversity, development and evolution of polyclad flatworm larvae. EvoDevo 2014, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- McNab, J.M.; Rodríguez, J.; Karuso, P.; Williamson, J.E. Natural products in polyclad flatworms. Mar. Drugs 2021, 19, 47. [Google Scholar] [CrossRef]
- McNab, J.M.; Briggs, M.T.; Williamson, J.E.; Hoffmann, P.; Rodriguez, J.; Karuso, P. Structural Characterization and Spatial Mapping of Tetrodotoxins in Australian Polyclads. Mar. Drugs 2022, 20, 788. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Boulder Size Range (cm) | Wentworth Category |
|---|---|
| 0.10–0.40 | Coarse sand |
| 0.41–0.80 | Fine gravel |
| 0.81–1.60 | Coarse gravel |
| 1.61–3.20 | Medium gravel |
| 3.21–6.40 | Cobble |
| 6.41–12.80 | Coarse cobble |
| 12.81–25.60 | Small boulder |
| 25.61–51.20 | Medium boulder |
| 51.21–102.40 | Large boulder |
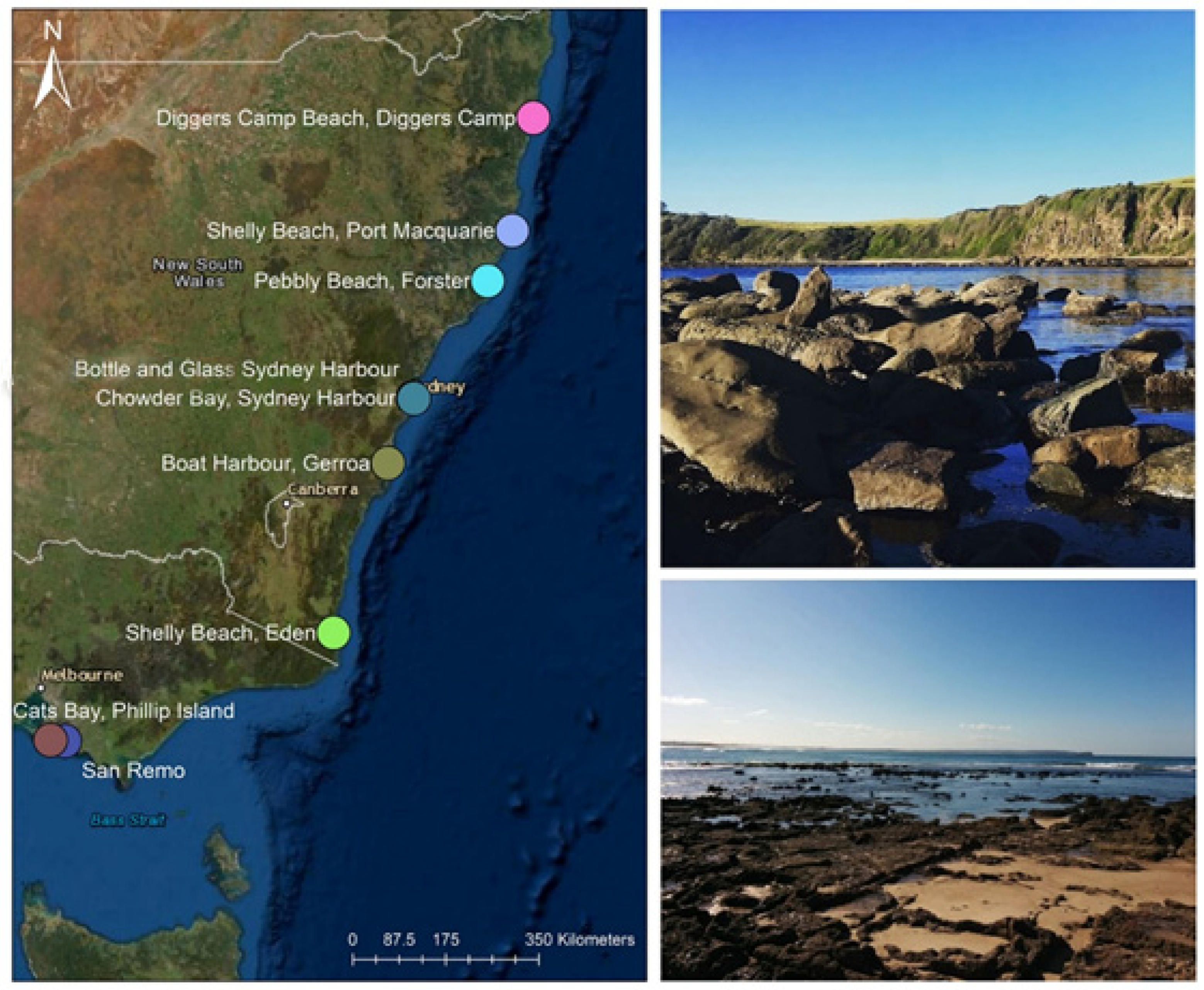
| State | Locality | Latitude | Longitude | Date | Sampling Time | Baardseth′s Wave Exposure Index |
|---|---|---|---|---|---|---|
| New South Wales | Diggers Camp Beach, Diggers Camp | 29°49′01.6″ S | 153°17′34.4″ E | 8 December 2019 | 12:00 p.m.–14:00 p.m. | 8 |
| Shelly Beach, Port Macquarie | 31°27′27.7″ S | 152°56′04.4″ E | 7 January 2020 | 12:30 p.m.–14:30 p.m. | 4 | |
| Pebbly Beach, Forster | 32°10′46.0″ S | 152°31′10.6″ E | 6 December 2019 | 09:00 a.m.–11:00 a.m. | 8 | |
| Chowder Bay, Sydney Harbour | 33°50′19.8″ S | 151°15′16.2″ E | 20 February 2020 | 13:30 p.m.–15:30 p.m. | 0 | |
| Bottle and Glass, Sydney Harbour | 33°50′54.0″ S | 151°16′13.1″ E | 25 October 2019; 21 February 2020 | 12:00 p.m.–14:00 p.m.; 14:30 p.m.–16:30 p.m. | 0 | |
| Boat Harbour, Gerroa | 34°45′02.0″ S | 150°49′56.5″ E | 7 July 2018 | 07:30 a.m.–8:30 a.m. * | 9 | |
| Shelly Beach, Eden | 37°04′22.0″ S | 149°54′45.6″ E | 10 July 2018 | 11:20 a.m.–12:20 p.m. * | 0 | |
| Victoria | San Remo | 38°31′11.9″ S | 145°22′02.2″ E | 13 July 2018 | 07:30 a.m.–8:30 a.m. * | 3 |
| Cats Bay, Phillip Island | 38°30′24.6″ S | 145°07′33.8″ E | 12 July 2018 | 16:00 p.m.–17:00 p.m. * | 3 |
| Family | Species | Locality | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diggers Camp | Port Macquarie | Forster | Chowder Bay | Bottle and Glass | Boat Harbour | Shelly Eden | San Remo | Phillip Island | ||
| Acotylea | ||||||||||
| Gnesiocerotidae | Echinoplana celerrima Haswell, 1907 | 6 | 10 | 7 | 4 | 9 | 4 | 4 | 4 | |
| Ceratoplana falconerae Rodriguez et al., 2021 | 1 | |||||||||
| Notocomplanidae | Notocomplana distincta (Prudhoe, 1982) | 1 | ||||||||
| Notoplanidae | Notoplana australis (Schmarda, 1859) | 17 | 3 | |||||||
| Notoplana felis (Rodriguez et al., 2021) | 1 | |||||||||
| Notoplana longiducta Hyman, 1959 | 1 | 7 | ||||||||
| Pseudostylochidae | Tripylocelis typica Haswell, 1907 | 2 | 2 | 2 | ||||||
| Planoceridae | Planocera edmondsi Prudhoe, 1982 | 1 | ||||||||
| Planocera sp. | 1 | |||||||||
| Stylochidae | Leptostylochus victoriensis Beveridge, 2017 | 2 | ||||||||
| Stylochus sp. | 2 | 4 | ||||||||
| Cotylea | ||||||||||
| Cestoplanidae | Cestoplana rubrocincta (Grube, 1840) | 2 | 2 | |||||||
| Euryleptididae | Eurylepta sp. | 1 | ||||||||
| Prosthiostomidae | Enchiridium sp. | 1 | ||||||||
| Pseudocerotidae | Pseudoceros sp. | 1 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tosetto, L.; McNab, J.M.; Hutchings, P.A.; Rodríguez, J.; Williamson, J.E. Fantastic Flatworms and Where to Find Them: Insights into Intertidal Polyclad Flatworm Distribution in Southeastern Australian Boulder Beaches. Diversity 2023, 15, 393. https://doi.org/10.3390/d15030393
Tosetto L, McNab JM, Hutchings PA, Rodríguez J, Williamson JE. Fantastic Flatworms and Where to Find Them: Insights into Intertidal Polyclad Flatworm Distribution in Southeastern Australian Boulder Beaches. Diversity. 2023; 15(3):393. https://doi.org/10.3390/d15030393
Chicago/Turabian StyleTosetto, Louise, Justin M. McNab, Pat A. Hutchings, Jorge Rodríguez, and Jane E. Williamson. 2023. "Fantastic Flatworms and Where to Find Them: Insights into Intertidal Polyclad Flatworm Distribution in Southeastern Australian Boulder Beaches" Diversity 15, no. 3: 393. https://doi.org/10.3390/d15030393
APA StyleTosetto, L., McNab, J. M., Hutchings, P. A., Rodríguez, J., & Williamson, J. E. (2023). Fantastic Flatworms and Where to Find Them: Insights into Intertidal Polyclad Flatworm Distribution in Southeastern Australian Boulder Beaches. Diversity, 15(3), 393. https://doi.org/10.3390/d15030393