


A New Genus and Species of †Cladocyclidae (Teleostei: †Ichthyodectiformes) from the Lower Cretaceous “Batateira Beds”, Barbalha Formation, Araripe Basin: The First Vertebrate Record in a Still Poorly Explored Fossil Site
 , , and
, , and 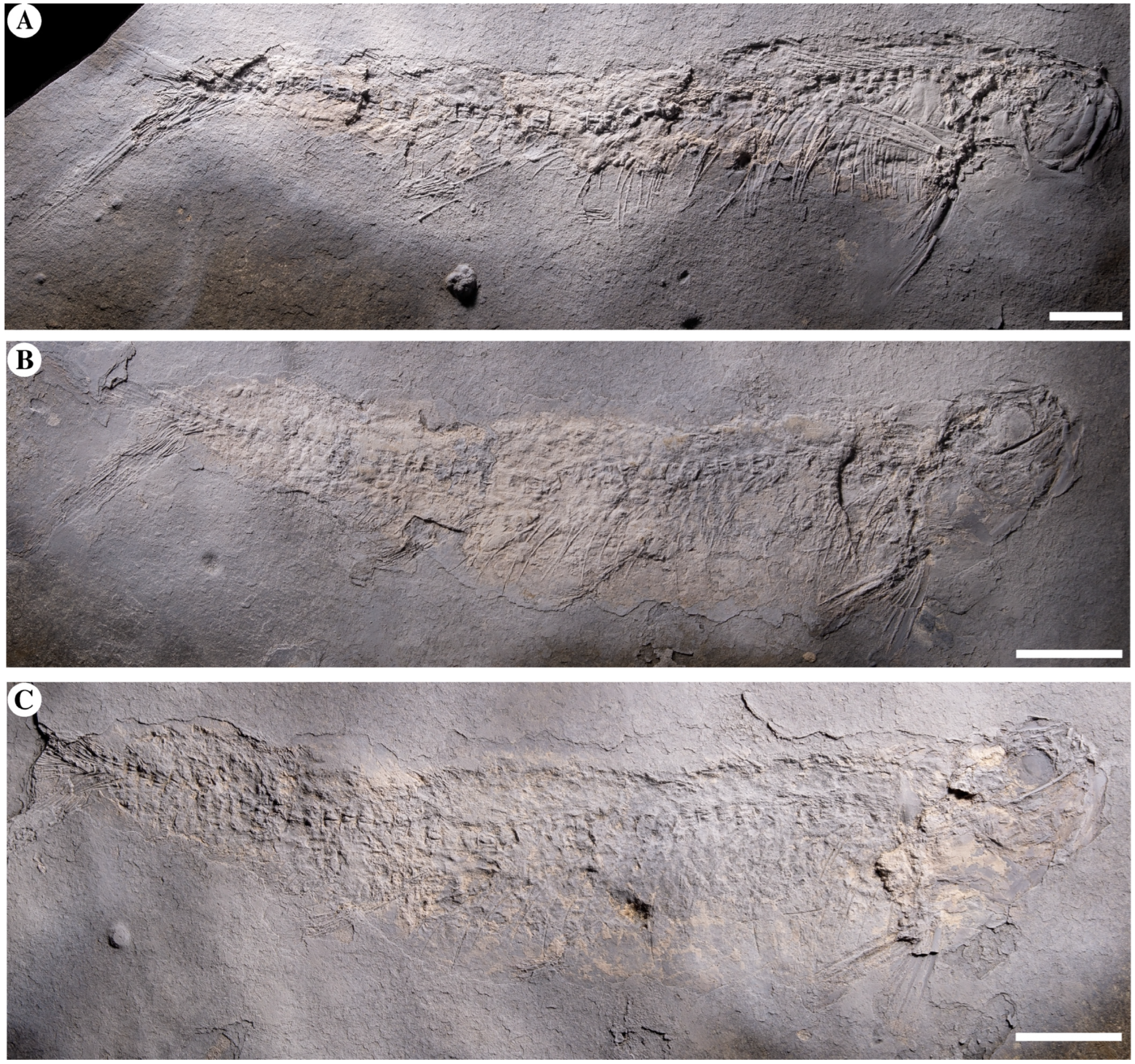{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Institutional Abbreviations
2.2. Anatomical Abbreviations
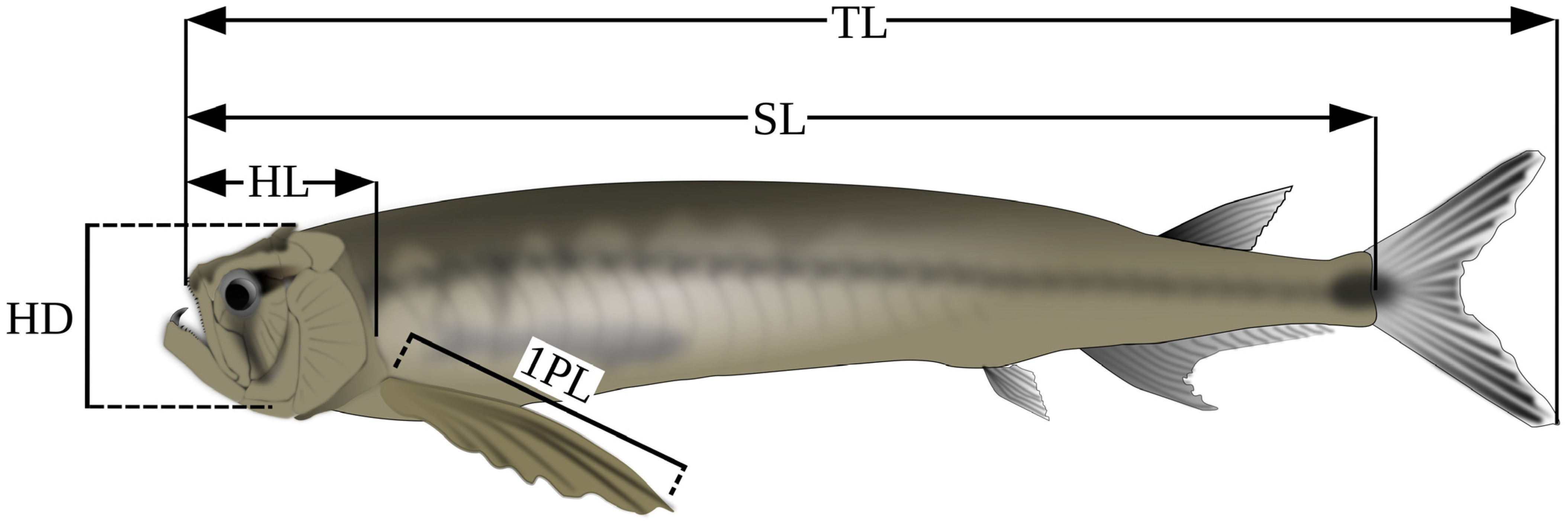
2.3. Measurements Abbreviations
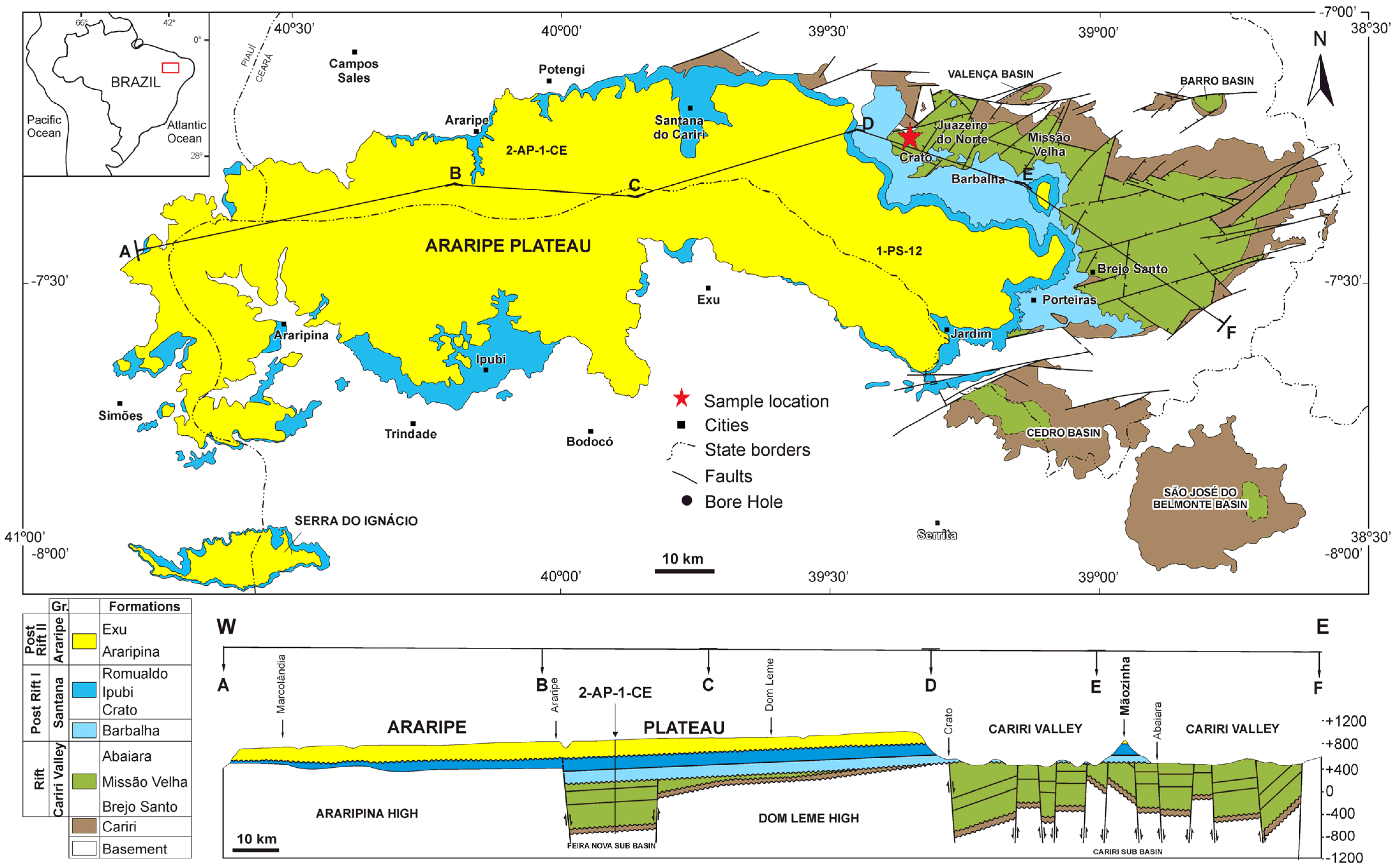
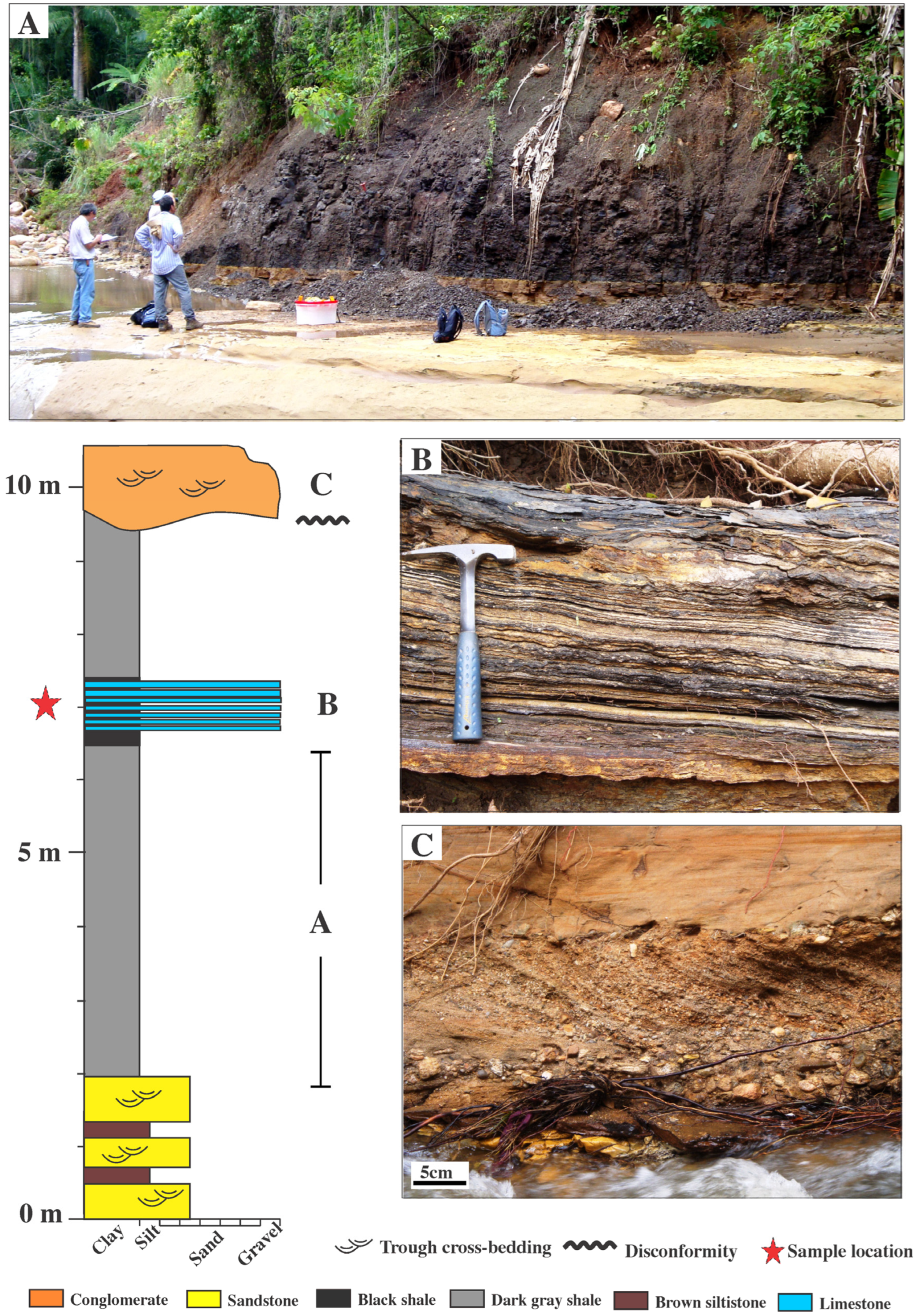
3. Geological Settings
4. Results
4.1. Systematic Palaeontology
- Teleostei Müller, 1846 [49]
- Order †Ichythyodectiformes Bardack and Sprinkle, 1969 [1]
- Suborder †Ichthyodectoidei Romer, 1966 [19]
- Family †Cladocyclidae Maisey, 1991 [5]
- †Cladocynodon gen. nov.
- Type species †Cladocynodon araripensis sp. nov.
- Zoobank LSID. urn:lsid:zoobank.org:act:3D7833AD-294A-4897-9496-F0220BCAAB1E
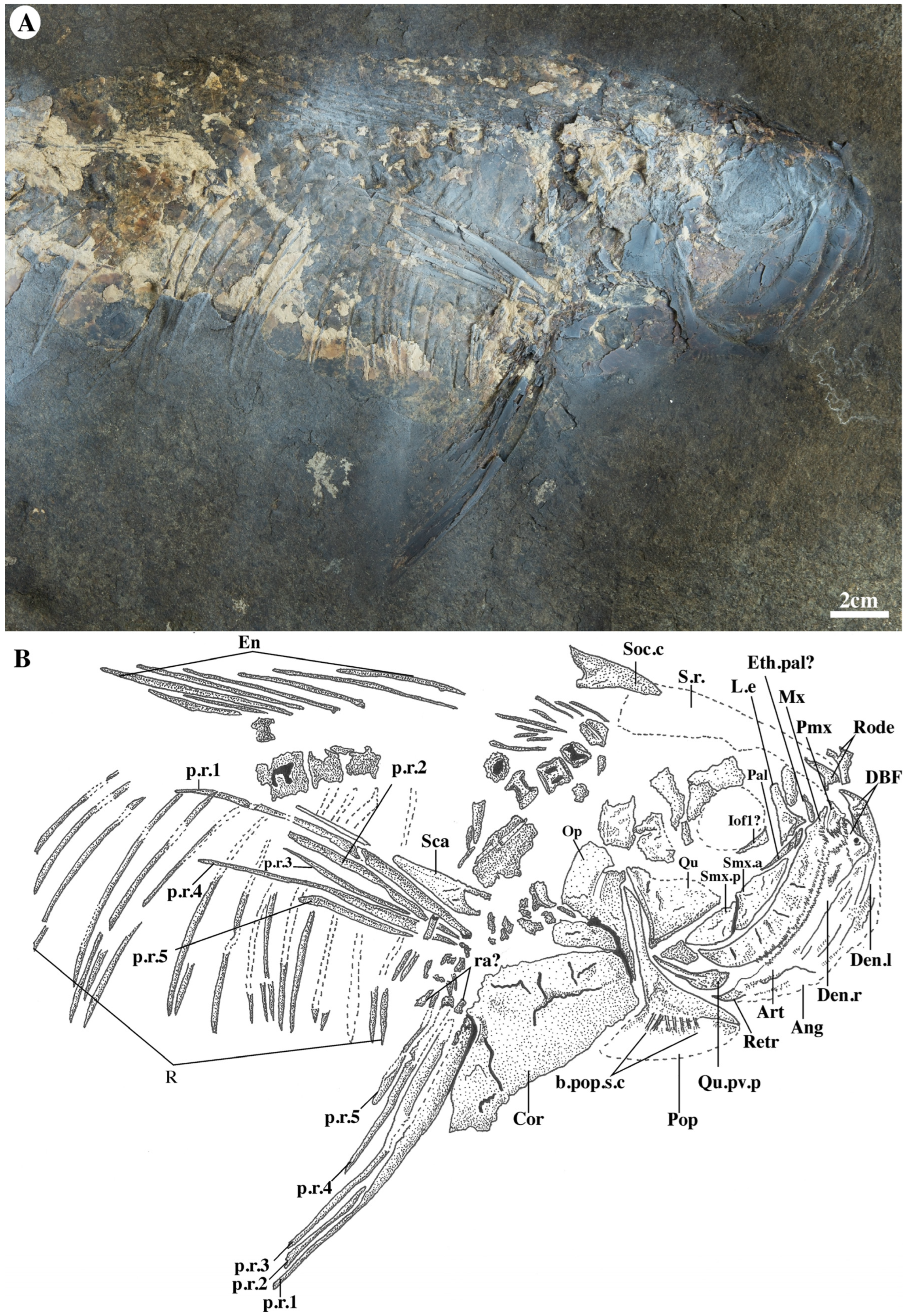
4.2. Anatomical Description
4.2.1. General Aspect, Morphometrics, and Meristics
4.2.2. Neurocranium
4.2.3. Hyopalatine and Circumorbital Series
4.2.4. Jaws
4.2.5. Opercular Series
4.2.6. Pectoral Fin and Girdle
4.2.7. Vertebral Column
4.2.8. Pelvic, Anal, and Dorsal Fins
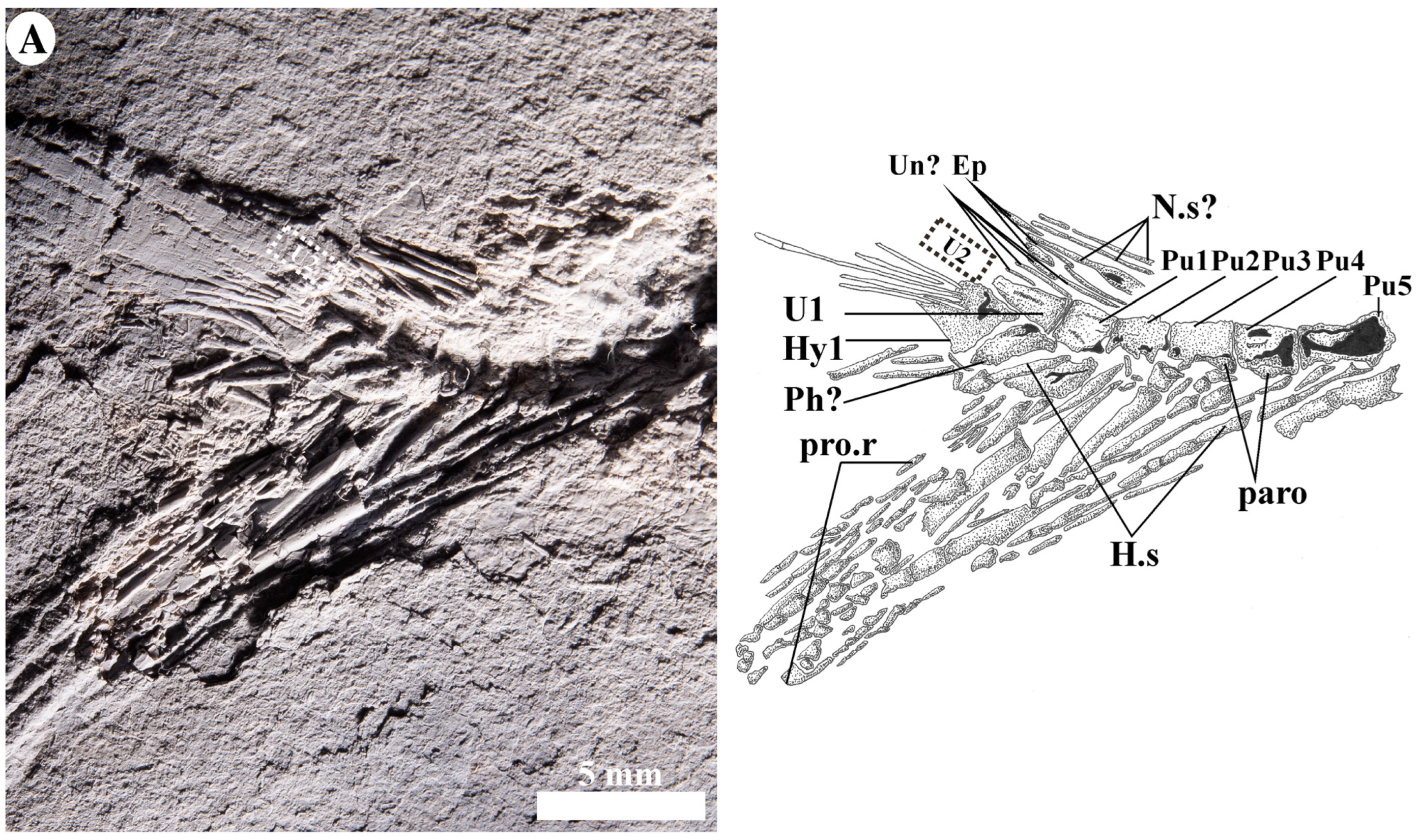
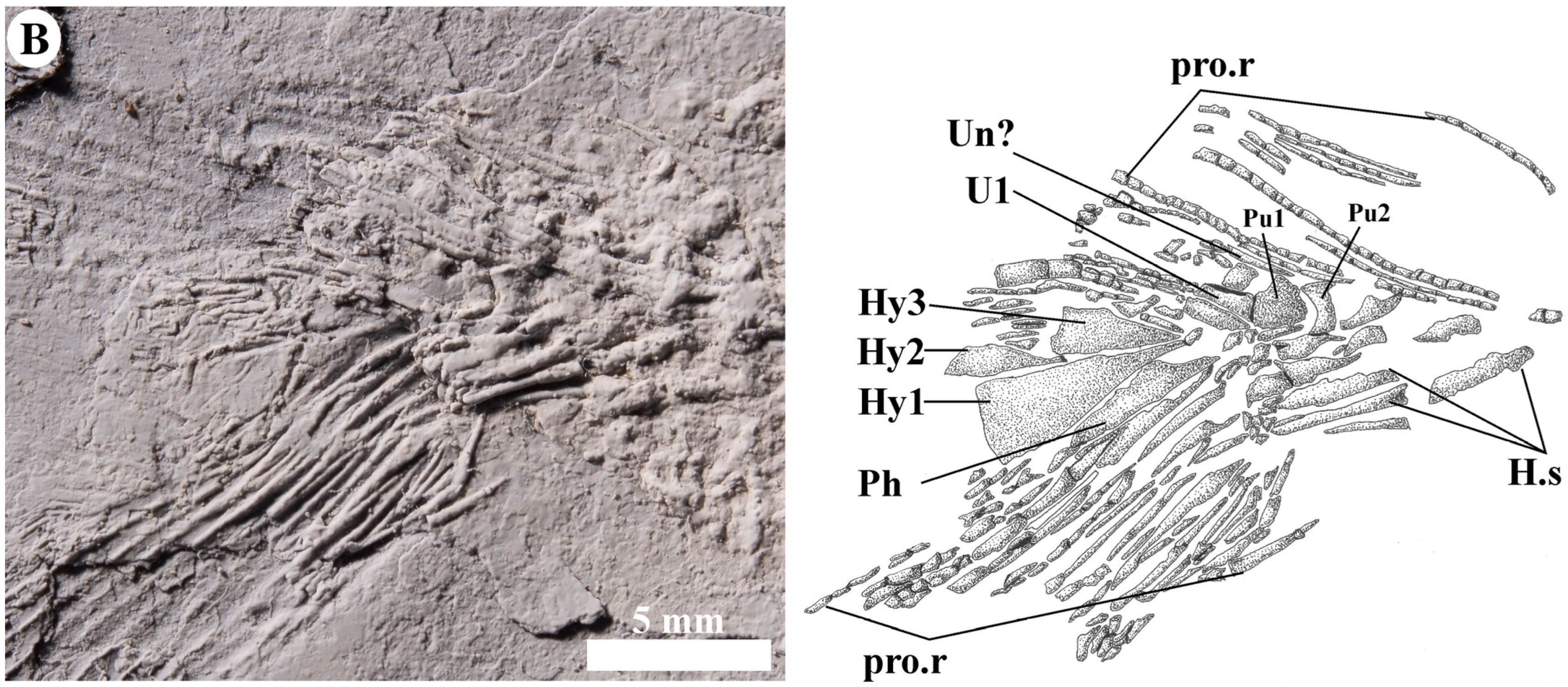
4.2.9. Caudal Skeleton
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardack, D.; Sprinkle, G. Morphology and relationships of saurocephalid fishes. Fieldiana Geol. 1969, 16, 297–340. [Google Scholar]
- Bardack, D. Anatomy and Evolution of Chirocentrid Fishes. In The University of Kansas Paleontological Contributions; University of Kansas Paleontological Institute: Lawrence, KS, USA, 1965; Volume 10, pp. 1–88. [Google Scholar]
- Taverne, L. Ostéologie et position systématique de Chirocentrites vexillifer de la Mésogée Eurafricaine. Considérations sur la phylogénie des Ichthyodectiformes, poisons Téléostéens du Jurassique et du Crétacé. Ann. Soc. R. Zool. Belg. 1986, 116, 33–54. [Google Scholar]
- Taverne, L. Les Ichthydectidae (Teleostei, Ichthyodectiformes) des schistes bitumineux de l’Aptien (Crétacé inférieur) de la Guinée Équatorial et du Gabon. Bull. Inst. R. Sci. Nat. Belg. Sci. Terre 2010, 80, 115–143. [Google Scholar]
- Maisey, J.G. (Ed.) Cladocyclus Agassiz, 1841. In Santana Fossils: An Illustrated Atlas; T.F.H. Publications Inc.: Neptune, NJ, USA, 1991; pp. 190–207. [Google Scholar]
- Arratia, G.; Scasso, R.A.; Kiessling, W. Late Jurassic Fishes from Longing Gap, Antarctic Peninsula. J. Vertebr. Paleontol. 2004, 24, 41–55. [Google Scholar] [CrossRef]
- Leal, M.E.C.; Brito, P.M. The ichthyodectiform Cladocyclus gardneri (Actinopterygii: Teleostei) from the Crato and Santana Formations, Lower Cretaceous of Araripe Basin, North-Eastern Brazil. Ann. Paléontol. 2004, 90, 103–113. [Google Scholar] [CrossRef]
- Alvarado-Ortega, J. Sistemática de los peces Ichthyodectiformes de la Cantera Tlayúa, Puebla, México. Ph.D. Thesis, Universidad Nacional Autónoma de México, Instituto de Geología, Mexico City, Mexico, 2005. [Google Scholar]
- Forey, P.L.; Cavin, L. A new species of Cladocyclus (Teleostei: Ichthyodectiformes) from the Cenomanian of Morocco. Palaeontol. Electron. 2007, 10, 1–10. [Google Scholar]
- Alvarado-Ortega, J.; Brito, P.M. A new ichthyodectiform (Actinopterygii, Teleostei) from the Lower Cretaceous of Marizal Formation, north-east Brazil. Palaeontology 2010, 53, 297–306. [Google Scholar] [CrossRef]
- Carrilo-Briceño, J.; Alvarado-Ortega, J.; Torres, C. Primer registro de Xiphactinus Leidy, 1870 (Teleostei, Ichthydectiformes) en el Cretácico Superior de América del Sur (Formación La Luna, Venezuela). Rev. Bras. Paleontol. 2012, 15, 327–335. [Google Scholar] [CrossRef]
- Cavin, L.; Forey, P.L.; Giersch, S. Osteology of Eubiodectes libanicus (Pictet & Humbert, 1866) and some other ichthyodectiformes (Teleostei): Phylogenetic implications. J. Syst. Palaeontol. 2013, 11, 115–177. [Google Scholar]
- Berrel, R.W.; Alvarado-Ortega, J.; Yabumoto, Y.; Salisbury, S.W. The first record of ichthydectiform fish Cladocyclus from eastern Gowdwana: A new species from the Lower Cretaceous of Queensland, Australia. Acta Paleontol. Pol. 2014, 59, 903–920. [Google Scholar]
- Kim, H.M.; Chang, M.M.; Wu, F.; Kim, Y.H. A new ichthyodectiform (Pisces, Teleostei) from the Lower Cretaceous of South Korea and its paleobiogeographic implications. Cretac. Res. 2014, 47, 117–130. [Google Scholar] [CrossRef]
- Yabumoto, Y.; Hirose, K.; Brito, P.M. A new ichthyodectiform fish, Amakusaichthys goshouraensis gen. et sp. nov. from the Upper Cretaceous (Santonian) Himenoura Group in Goshoura, Amakusa, Kumamoto, Japan. Hist. Biol. 2018, 32, 362–375. [Google Scholar] [CrossRef]
- Bahrami, A.; Yazdi, M.; Moghaddam, H.V.; Ghobadi, A.; Murray, A.M.; Brinkman, D.; Parvaneh-Nejad, M.S.; Ataabadi, M.M.; Kundrát, M. Early Cretaceous vertebrate and invertebrate fossils from Dariyan Formation, southern Iran. Hist. Biol. 2021, 33, 387–402. [Google Scholar] [CrossRef]
- Alvarado-Ortega, J.; Porras-Múzquiz, H.G. On the presence of Xiphactinus (Teleostei, Ichthyodectiformes) in the Coniacian-Campanian marine deposits of the Piedritas site, Coahuila, northern Mexico. Rev. Mex. De Cienc. Geol. 2022, 39, 142–150. [Google Scholar] [CrossRef]
- Patterson, C.; Rosen, D.E. Review of Ichthyodectiform and Other Mesozoic Teleost Fishes and the Theory and Practice of Classifying Fossils. Bull. Am. Mus. Nat. Hist. 1977, 158, 81–172. [Google Scholar] [CrossRef] [Green Version]
- Romer, A.S. Vertebrate Paleontology; University of Chicago Press: Chicago, IL, USA, 1966; pp. 1–486. [Google Scholar]
- Patterson, C. The contribution of paleontology to teleost phylogeny. In Major Patterns in Vertebrate Evolution; Hecht, M.K., Goody, P.C., Hecht, B.M., Eds.; Plenum Press: New York, NY, USA, 1977; pp. 579–643. [Google Scholar]
- Arratia, G. Basal teleosts and teleostean phylogeny. PalaeoIchthyologica 1997, 7, 1–168. [Google Scholar]
- Arratia, G. The monophyly of Teleostei and stem-group teleosts. In Mesozoic Fishes 2-Systematics and Fossil Record; Arratia, G., Schultze, H.-P., Eds.; Verlag Pfeil: München, Germany, 1999; pp. 265–334. [Google Scholar]
- Allis, E.F. The cranial anatomy of the mail checked fishes. Zoologica 1909, 22, 1–219. [Google Scholar]
- Starks, E.C. Bones of the ethmoid region of the fish skull. Stanford Univ. Publ. Biol. Sci. 1926, 4, 137–338. [Google Scholar]
- Harrington, R.W. The osteocranium of the american cyprinid fish, Notropis bifrenatus, with an annotated synonymy of teleost skull bones. Copeia 1955, 4, 267–290. [Google Scholar] [CrossRef]
- Dornesco, G.T.; Soresco, C. Sur le development et la valeur morphologique de région ethmoïdale de la Carpe. Anat. Anz. 1971, 129, 33–52. [Google Scholar]
- Patterson, C. The braincase of the pholidophorid and leptolepid fishes, with a review of the actinopterygian braincase. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1975, 269, 275–579. [Google Scholar] [PubMed]
- Fink, S.V.; Fink, W.L. Interrelationships of the ostariophysan fishes (Teleostei). Zool. J. Linn. Soc. 1981, 72, 297–353. [Google Scholar] [CrossRef]
- Howes, J.G. Cranial muscles of gonorynchiforms fishes, with comments on generic relationships. Bull. Br. Mus. Nat. Hist. Zool. 1985, 49, 273–303. [Google Scholar]
- Brito, P.M.; Yabumoto, Y. An updated review of the fish faunas from the Crato and Santana formations in Brazil, a close relationship to the Tethys fauna. Bull. Kitakushu Mus. Nat. Hum. Hist. 2011, 9, 107–136. [Google Scholar]
- Wenz, S.; Campos, D.A. Ensaio sobre a distribuição estrtaigráfica de Cladocyclus (Pisces, Ichthyodectiformes do Cretáceo Inferior do Nordeste brasileiro. In Proceedings of the Resumo de Comunicações do IX Congresso Brasileiro de Paleontologia, Fortaleza, Brasil, 1–7 September 1985; p. 10. [Google Scholar]
- Ponte, F.C.; Appi, C.J. Proposta de revisão estratigráfica da Bacia do Araripe. An. Acad. Bras. Cienc. 1990, 1, 211–226. [Google Scholar]
- Rios-Netto, A.M.; Paula-Freitas, A.B.L.; Carvalho, I.S.; Regali, M.S.P.; Borgui, L.; Freitas, F.I. Formalização estatrigráfica do Membro Fundão, Formação Rio da Batateira, Cretáceo Inferior da Bacia do Araripe, Nordeste do Brasil. Rev. Bras. Geociênc. 2012, 42, 281–292. [Google Scholar]
- Assine, M.L.; Perinotto, J.A.J.; Custódio, M.A.; Neumann, V.H.; Varejão, F.G.; Mescolotti, P.C. Sequências deposicionais do Andar Alagoas da Bacia do Araripe, Nordeste do Brasil. Bol. Geociênc. Petrobrás 2014, 22, 3–28. [Google Scholar]
- Carvalho, I.S.; Rios-Netto, A.M.; Borgui, L.; Freitas, A.P.; Leonardi, G.; Andrade, J.A.; Freitas, F.I. Dinosaur trampling from the Rio da Batateira Formation—Lower Cretaceous of Araripe Brasil, Brazil. In Proceedings of the Livro de Resumos of Paleo Fall Meeting, Évora, Portugal, 26–28 September 2019; p. 21. [Google Scholar]
- Toombs, H.A.; Rixon, A.E. The use of acids in the preparation of vertebrate fossils. Curator Mus. J. 1959, 2, 304–312. [Google Scholar] [CrossRef]
- Martill, D.M. The preservation of fossil fishes in concretions from the Cretaceous of Brazil. Palaeontology 1988, 30, 1–18. [Google Scholar]
- Assine, M.L. Análise estratigráfica da Bacia do Araripe, Nordeste do Brasil. Rev. Bras. Geociênc. 1992, 22, 289–300. [Google Scholar] [CrossRef]
- Assine, M.L. Bacia do Araripe. Bol. Geociênc. Petrobrás 2007, 15, 371–389. [Google Scholar]
- Vila Nova, B.C.; Sayão, J.M.; Neumann, V.H.M.L.; Kellner, A.W.A. Redescription of Cearadactylus atrox (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation (Santana Group) of the Araripe Basin, Brazil. J. Vertebr. Paleontol. 2014, 34, 126–134. [Google Scholar] [CrossRef]
- Melo, R.B.; Guzmán, J.; Almeida-Lima, D.; Piovesan, E.K.; Neumann, V.H.M.L.; Souza, A.J. New Marine data and age accuracy of the Romualdo Formation, Araripe Basin, Brazil. Sci. Rep. 2020, 10, 15779. [Google Scholar] [CrossRef]
- Martill, D.M.; Brito, P.M.; Donovan, S.K. There are rudists in Brazil! Derived examples of cf. Amphitriscoelus Harris and Hodson, 1992, in the Araripe Basin, north-east Brazil: Implications for dating of the fossil lagerstätten of the Santana and Crato Formations. Cretac. Res. 2021, 120, 104718. [Google Scholar] [CrossRef]
- Matos, R.M.D. The Northeast Brazililian rift system. Tectonics 1992, 11, 766–791. [Google Scholar] [CrossRef]
- Neves, B.B.B.; Santos, E.J.; Van Schmus, W.R. Tectonic history of the Borborema Province, Northeastern Brazil. In Tectonic Evolution of South America, Proceedings 31st International Geological Congress, Rio de Janeiro, Brazil, 6–17 August, 2000; Cordanni, U.G., Milani, E.J., Thomaz-Filho, A., Campos, D.A., Eds.; The Brazilian Academy of Science: Rio de Janeiro, Brazil, 2000; pp. 151–182. [Google Scholar]
- Catto, B.; Jahnert, R.J.; Warren, L.V.; Varejão, F.G.; Assine, M.L. The microbial nature of laminated limestones: Lessons from the Upper Aptian, Araripe Basin, Brazil. Sediment. Geol. 2016, 341, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Souza, J.F.G.; Isozaki, Y.; Tsutsumi, Y.; Schmitt, R.S.; Medeiros, S.R.; Almeida, C.N.; Araújo, B.C.; Richetti, P.; Silva, E.A.; Rios-Netto, A.M. Provenance analysis of the Araripe intracontinental basin, northeast Brazil—Routes for proto-Atlantic marine incursions in northwest Gondwana. Sediment. Geol. 2022, 440, 106243. [Google Scholar] [CrossRef]
- Ponte, F.C.; Filho, F.C.P. Evolução tectônica e classificação da Bacia do Araripe. In Proceedings of the Bulletin of the 4th Symposium about the Cretaceous of Brazil, Águas de São Pedro, São Paulo, Brazil, 1996; August 18th-22Th; pp. 123–133. [Google Scholar]
- Martill, D.M.; Wilby, P.R. Stratigraphy. In Fossils of the Santana and Crato formations, Brazil. Field Guides to Fossils, 5th ed.; Martill, D.M., Ed.; The Palaeontological Association: London, UK, 1993; pp. 20–50. [Google Scholar]
- Müller, J. Über den Bau und die Grenzen der Ganoiden and über das natürliche System der Fische. Abh. Dtsch. Akad. Wiss. Berl. 1846, 145, 119–216. [Google Scholar]
- Stewart, J.D. A new genus of Saurodontidae (Teleostei: Ichthyodectiformes) from Upper Cretaceous rocks of the Western Interior North America. In Mesozoic Fishes 2-Systematics and Fossil Record; Arratia, G., Schultze, H.-P., Eds.; Verlag Pfeil: München, Germany, 1999; pp. 335–360. [Google Scholar]
- Taverne, L.; Capasso, L. Osteology and phylogenetic relationships of Furloichthys bonarellii gen. and sp. nov. (Teleostei, Ichthyodectidae), a tropical fish from the Upper Cretaceous of central Italy. Geo-Eco-Trop 2018, 42, 75–88. [Google Scholar]
- Mkhitaryan, T.D.; Averinov, A.O. New material and phylogenetic position of Aidachar paludalis Nesov, 1981 (Actinopterygii, Ichthyodectiformes) from the Late Cretaceous of Uzbekistan. Proc. Zool. Inst. RAS 2011, 315, 181–192. [Google Scholar] [CrossRef]
- Alvarado-Ortega, J. Description and relationships of a new Ichthyodectiformes fish from the Tlayúa Formation (Early Cretaceous: Albian), Puebla, Mexico. J. Vertbr. Paleontol. 2004, 24, 802–813. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mayrinck, D.; Ribeiro, A.C.; Assine, M.L.; Spigolon, A.L.D. A New Genus and Species of †Cladocyclidae (Teleostei: †Ichthyodectiformes) from the Lower Cretaceous “Batateira Beds”, Barbalha Formation, Araripe Basin: The First Vertebrate Record in a Still Poorly Explored Fossil Site. Diversity 2023, 15, 413. https://doi.org/10.3390/d15030413
de Mayrinck D, Ribeiro AC, Assine ML, Spigolon ALD. A New Genus and Species of †Cladocyclidae (Teleostei: †Ichthyodectiformes) from the Lower Cretaceous “Batateira Beds”, Barbalha Formation, Araripe Basin: The First Vertebrate Record in a Still Poorly Explored Fossil Site. Diversity. 2023; 15(3):413. https://doi.org/10.3390/d15030413
Chicago/Turabian Stylede Mayrinck, Diogo, Alexandre Cunha Ribeiro, Mario Luis Assine, and André Luis Durante Spigolon. 2023. "A New Genus and Species of †Cladocyclidae (Teleostei: †Ichthyodectiformes) from the Lower Cretaceous “Batateira Beds”, Barbalha Formation, Araripe Basin: The First Vertebrate Record in a Still Poorly Explored Fossil Site" Diversity 15, no. 3: 413. https://doi.org/10.3390/d15030413
APA Stylede Mayrinck, D., Ribeiro, A. C., Assine, M. L., & Spigolon, A. L. D. (2023). A New Genus and Species of †Cladocyclidae (Teleostei: †Ichthyodectiformes) from the Lower Cretaceous “Batateira Beds”, Barbalha Formation, Araripe Basin: The First Vertebrate Record in a Still Poorly Explored Fossil Site. Diversity, 15(3), 413. https://doi.org/10.3390/d15030413