

Forest Fragmentation and Developmental Stability of Wood Mice Apodemus sylvaticus: A Food-Mediated Effect?


Abstract
:1. Introduction
2. Material and Methods
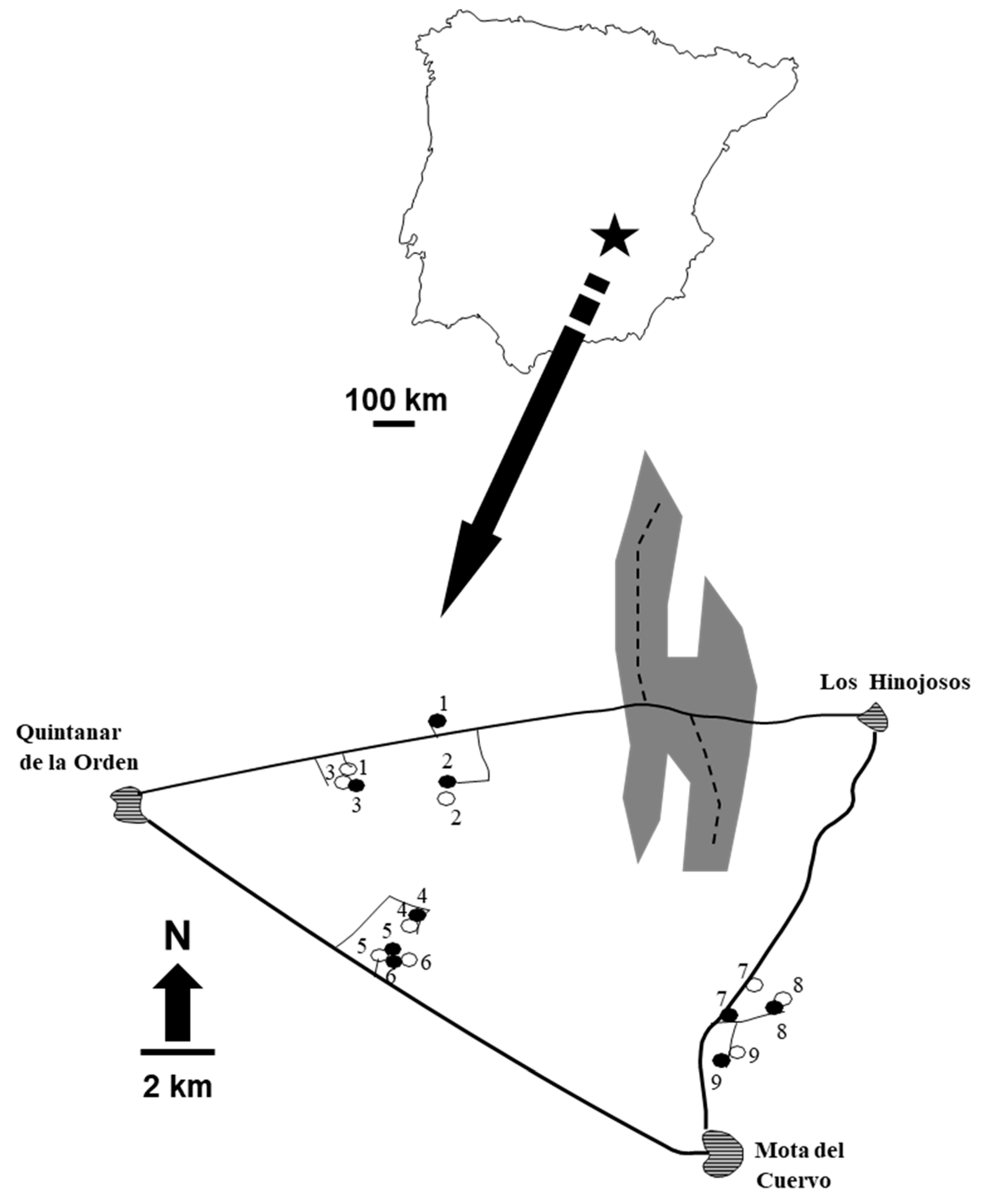
2.1. Study Area and Experimental Design
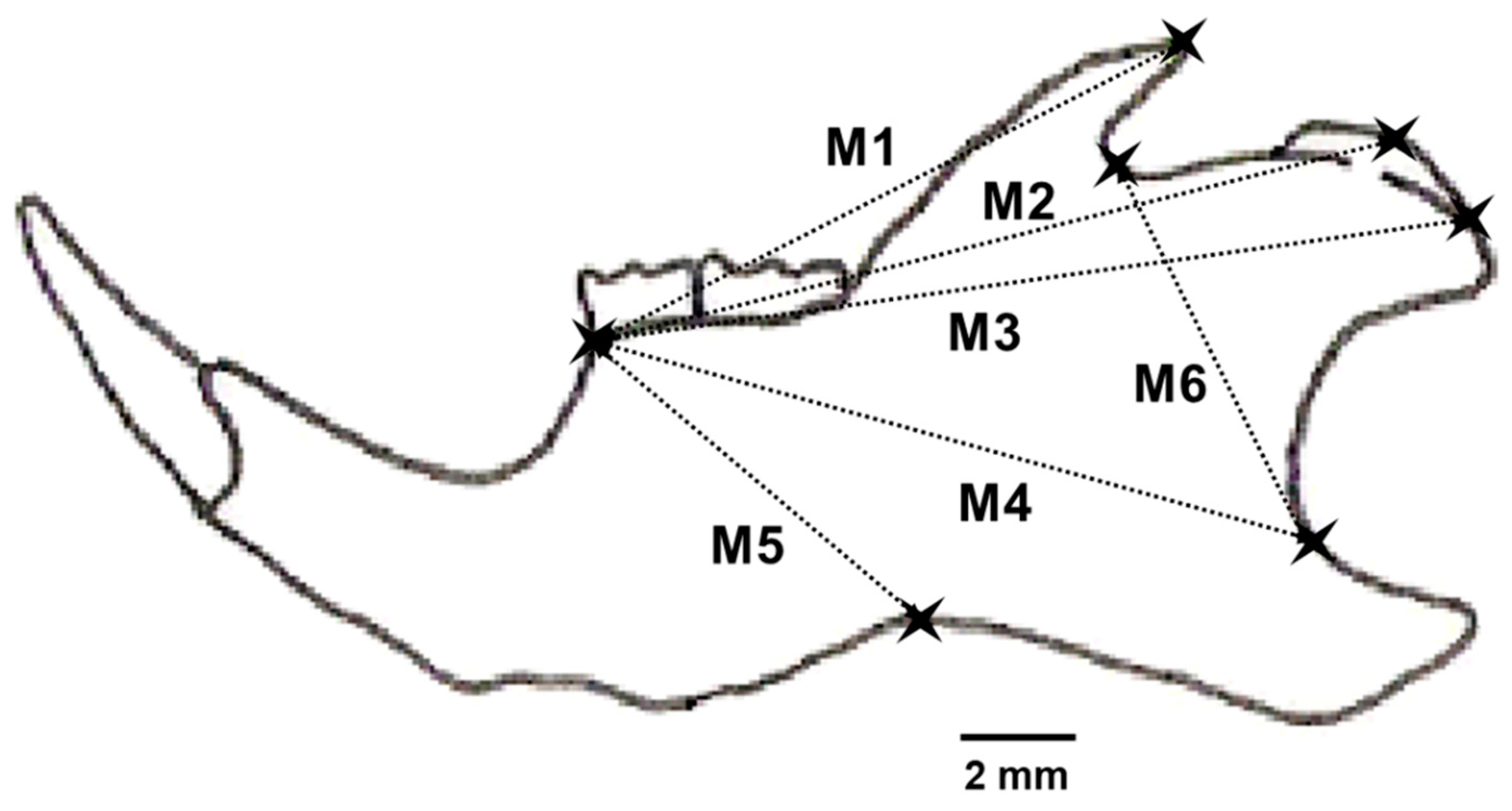
2.2. Fluctuating Asymmetry of Lower Mandibles
2.3. Statistical Analyses
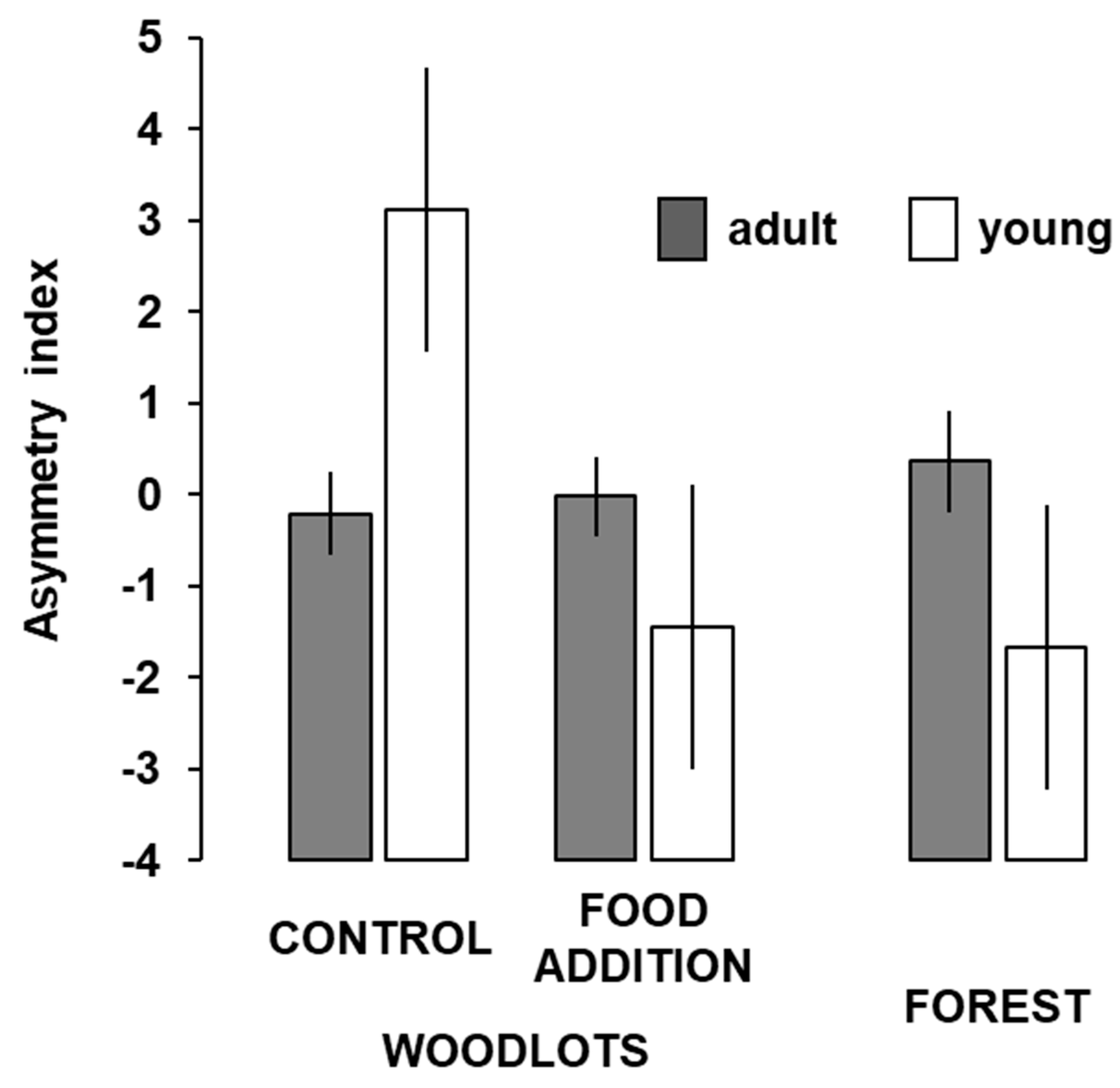
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Ann. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Butchart, S.H.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Gehring, T.M.; Swihart, R.K. Body size, niche breadth, and ecologically scaled responses to habitat fragmentation: Mammalian predators in an agricultural landscape. Biol. Conserv. 2003, 109, 283–295. [Google Scholar] [CrossRef]
- Lanchier, N.; Neuhauser, C. A spatially explicit model for competition among specialists and generalists in a heterogeneous environment. Ann. Appl. Prob. 2006, 16, 1385–1410. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Let. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Mattos, I.D.; Zimbres, B.; Marinho-Filho, J. Habitat specificity modulates the response of small mammals to habitat fragmentation, loss, and quality in a Neotropical savanna. Front. Ecol. Evol. 2021, 9, 751315. [Google Scholar] [CrossRef]
- Richmond, C.E.; Breitburg, D.L.; Rose, K.A. The role of environmental generalist species in ecosystem function. Ecol. Model. 2005, 188, 279–295. [Google Scholar] [CrossRef]
- Šálek, M.; Kreisinger, J.; Sedláček, F.; Albrecht, T. Do prey densities determine preferences of mammalian predators for habitat edges in an agricultural landscape? Landsc. Urban Plan. 2010, 98, 86–91. [Google Scholar] [CrossRef]
- Manson, R.H.; Ostfeld, R.S.; Canham, C.D. Long-term effects of rodent herbivores on tree invasion dynamics along forest-field edges. Ecology 2001, 82, 3320–3329. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhang, Z.; Wang, Y. Impacts of scatter-hoarding rodents on restoration of oil tea Camellia oleifera in a fragmented forest. For. Ecol. Manag. 2004, 196, 405–412. [Google Scholar] [CrossRef]
- Gómez, J.M.; Schupp, E.W.; Jordano, P. Synzoochory: The ecological and evolutionary relevance of a dual interaction. Biol. Rev. 2019, 94, 874–902. [Google Scholar] [CrossRef]
- Torre, I.; Arrizabalaga, A.; Freixas, L.; Ribas, A.; Flaquer, C.; Díaz, M. Using scats of a generalist carnivore as a tool to monitor small mammal communities in Mediterranean habitats. Basic Appl. Ecol. 2013, 14, 155–164. [Google Scholar] [CrossRef]
- Torre, I.; Díaz, M. Small mammal abundance in Mediterranean post-fire habitats: a role for predators? Acta Oecol. 2004, 25, 137–142. [Google Scholar] [CrossRef]
- Oró, D.; Sanz-Aguilar, A.; Carbonell, F.; Grajera, J.; Torre, I. Multi-species prey dynamics influence local survival in resident and wintering generalist predators. Oecologia 2021, 197, 437–446. [Google Scholar] [CrossRef]
- Santos, T.; Tellería, J.L. Vertebrate predation on Holm Oak, Quercus ilex, acorns in a fragmented habitat: Effects on seedling recruitment. For. Ecol. Manag. 1997, 98, 181–187. [Google Scholar] [CrossRef]
- Morán-López, T.; Fernández, M.; Alonso, C.L.; Flores, D.; Valladares, F.; Díaz, M. Effects of forest fragmentation on the oak-rodent mutualism. Oikos 2015, 124, 1482–1491. [Google Scholar] [CrossRef]
- Morán-López, T.; Wiegand, T.; Morales, J.M.; Valladares, F.; Díaz, M. Predicting forest management effects on oak—Rodent mutualisms. Oikos 2016, 125, 1445–1457. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Montemayor, E.; Ferryman, M.; Watts, K.; Macgregor, N.A.; Hambly, N.; Brennan, S.; Coxon, R.; Langridge, H.; Park, K.J. Small mammal responses to long-term large-scale woodland creation: The influence of local and landscape-level attributes. Ecol. Appl. 2020, 30, e02028. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, D.W.; Tew, T.E.; Todd, I.A.; Garner, J.P.; Johnson, P.J. Arable habitat use by wood mice (Apodemus sylvaticus). 3. A farm-scale experiment on the effects of crop rotation. J. Zool. 2000, 250, 313–320. [Google Scholar] [CrossRef]
- Alcántara, M.; Tellería, J.L. Habitat selection of the Wood mouse (Apodemus sylvaticus) in cereal steppes of Central Spain. Z. Säugetierkunde 1991, 56, 347–351. [Google Scholar]
- Fitzgibbon, C.D. Small mammals in farm woodlands: The effects of habitat, isolation and surrounding land-use patterns. J. Appl. Ecol. 1997, 34, 530–539. [Google Scholar] [CrossRef]
- Ouin, A.; Paillat, G.; Butet, A.; Burel, F. Spatial dynamics of wood mouse (Apodemus sylvaticus) in an agricultural landscape under intensive use in the Mont Saint Michel Bay (France). Agric. Ecosyst. Environ. 2000, 78, 159–165. [Google Scholar] [CrossRef]
- Todd, I.A.; Tew, T.E.; Macdonald, D.W. Habitat use of the arable ecosystem by wood mice, Apodemus sylvaticus. 1. Macrohabitat. J. Zool. 2000, 250, 299–303. [Google Scholar] [CrossRef]
- Díaz, M.; Santos, T.; Tellería, J.L. Effects of forest fragmentation on the winter body condition and population parameters of an habitat generalist, the wood mouse Apodemus sylvaticus: A test of hypotheses. Acta Oecol. 1999, 20, 39–49. [Google Scholar] [CrossRef]
- Morán-López, T.; Forner, A.; Flores-Rentería, D.; Díaz, M.; Valladares, F. Some positive effects of the fragmentation of holm oak forests: Attenuation of water stress and enhancement of acorn production. For. Ecol. Manag. 2016, 370, 22–30. [Google Scholar] [CrossRef] [Green Version]
- García, F.J.; Díaz, M.; De Alba, J.M.; Alonso, C.L.; Carbonell, R.; De Carrión, M.L.; Monedero, C.; Santos, T. Edge effects and patterns of winter abundance of wood mice Apodemus sylvaticus in Spanish fragmented forests. Acta Theriol. 1998, 43, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.; Torre, I.; Arrizabalaga, A. Relative roles of density and rainfall on the short-term regulation of Mediterranean wood mouse Apodemus sylvaticus populations. Acta Theriol. 2010, 55, 251–260. [Google Scholar] [CrossRef]
- Rosário, I.T.; Mathias, M.L. Annual weight variation and reproductive cycle of the wood mouse (Apodemus sylvaticus) in a Mediterranean environment. Mammalia 2004, 68, 133–140. [Google Scholar] [CrossRef]
- Díaz, M.; Alonso, C.L. Wood mouse Apodemus sylvaticus winter food supply: Density, condition, breeding, and parasites. Ecology 2003, 84, 2680–2691. [Google Scholar] [CrossRef]
- Møller, A.P.; Swaddle, J.P. Asymmetry, Developmental Stability, and Evolution; Oxford University Press: Oxford, UK, 1997; ISBN 0-19-854895-8. [Google Scholar]
- Wayne, R.K.; Modi, W.S.; O’Brien, S.J. Morphological variability and asymmetry in the cheetah (Acinonyx jubatus), a genetically uniform species. Evolution 1986, 40, 78–85. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Shadrina, E.; Vol’pert, Y. Fluctuating asymmetry of craniological features of small mammals as a reflection of heterogeneity of natural populations. Symmetry 2016, 8, 142. [Google Scholar] [CrossRef] [Green Version]
- Floate, K.D.; Coghlin, P.C. No support for fluctuating asymmetry as a biomarker of chemical residues in livestock dung. Can. Entomol. 2010, 142, 354–368. [Google Scholar] [CrossRef]
- Vangestel, C.; Lens, L. Does fluctuating asymmetry constitute a sensitive biomarker of nutritional stress in house sparrows (Passer domesticus)? Ecol. Ind. 2011, 11, 389–394. [Google Scholar] [CrossRef]
- Leung, B.; Forbes, M.R.; Houle, D. Fluctuating asymmetry as a bioindicator of stress: Comparing efficacy of analyses involving multiple traits. Am. Nat. 2000, 155, 101–115. [Google Scholar] [CrossRef]
- Leamy, L. Heritability of directional and fluctuating asymmetry for mandibular characters in random-bred mice. J. Evol. Biol. 1999, 12, 146–155. [Google Scholar] [CrossRef]
- Yezerinac, S.M.; Lougheed, S.C.; Handford, P. Measurement error and morphometric studies: Statistical power and observer experience. Syst. Biol. 1992, 41, 471–482. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Witter, M.S.; Cuthill, I.C. The analysis of fluctuating asymmetry. Anim. Behav. 1994, 48, 986–989. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.; Pulido, F.J.; Møller, A.P. Herbivore effects on developmental instability and fecundity of holm oaks. Oecologia 2004, 139, 224–234. [Google Scholar] [CrossRef]
- Nachman, G.; Heller, K.E. Fluctuating asymmetry as an index of fitness: Causality or statistical artifact? Oikos 1999, 86, 357–365. [Google Scholar] [CrossRef]
- Møller, A.P.; Gangestad, S.W.; Thornhill, R. Nonlinearity and the importance of fluctuating asymmetry as a predictor of fitness. Oikos 1999, 86, 366–368. [Google Scholar] [CrossRef]
- Klingenberg, C. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; Freeman: New York, NY, USA, 1981; ISBN 0-7167-2411-1. [Google Scholar]
- Leamy, L.J.; Meagher, S.; Taylor, S.; Carroll, L.; Potts, W.K. Size and fluctuating asymmetry of morphometric characters in mice: Their associations with inbreeding and t-haplotype. Evolution 2001, 55, 2333–2341. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Developmental stability and ideal despotic distribution of blackbirds in a patchy environment. Oikos 1995, 72, 228–234. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating asymmetry: An epigenetic measure of stress. Biol. Rev. Camb. Phil. Soc. 1990, 65, 131–145. [Google Scholar] [CrossRef]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef] [Green Version]
- Tew, T.E.; Macdonald, D.W. The effects of harvest on arable wood mice Apodemus sylvaticus. Biol. Conserv. 1993, 65, 279–283. [Google Scholar] [CrossRef]
- Tattersall, F.H.; Macdonald, D.W.; Hart, B.J.; Manley, W.J.; Feber, R.E. Habitat use by wood mice (Apodemus sylvaticus) in a changeable arable landscape. J. Zool. 2001, 255, 487–494. [Google Scholar] [CrossRef]
- Tellería, J.L.; Santos, T.; Alcántara, M. Abundance and food-searching intensity of wood mice (Apodemus sylvaticus) in fragmented forests. J. Mammal. 1991, 72, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.; Batzli, G.O. Effects of forest edge on populations of white-footed mice Peromyscus leucopus. Ecography 2002, 25, 193–199. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Hines, J.E.; Knutson, M.G.; Franklin, A.B. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 2003, 84, 2200–2207. [Google Scholar] [CrossRef] [Green Version]
- Swaddle, J.P.; Witter, M.S. On the ontogeny of developmental stability in a stabilized trait. Proc. R. Soc. Lond. B 1997, 264, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Polak, M.; Trivers, R. The science of symmetry in biology. Trends Ecol. Evol. 1994, 9, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; Møller, A.P.; Pulido, F.J. Fruit abortion, developmental selection and developmental stability in Quercus ilex. Oecologia 2003, 135, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Hallgrímsson, B. Fluctuating asymmetry in Macaca fascicularis: A study of the etiology of developmental noise. Int. J. Primatol. 1993, 14, 421–443. [Google Scholar] [CrossRef]
- Chippindale, A.K.; Palmer, A.R. Persistence of subtle departures from symmetry over multiple molts in individual brachyuran crabs: Relevance to developmental stability. Genetica 1993, 89, 185–199. [Google Scholar] [CrossRef]
- Gileva, E.A.; Ialkovskaia, L.E.; Borodin, A.V.; Zykov, S.V.; Kshniasev, I.A. Fluctuating asymmetry of craniometric characters in rodents (Mammalia: Rodentia): interspecific and interpopulational comparisons. Zhurnal Obs. Biol. 2007, 68, 221–230. [Google Scholar]
- Kellner, J.R.; Alford, R.A. The ontogeny of fluctuating asymmetry. Am. Nat. 2003, 161, 931–947. [Google Scholar] [CrossRef] [Green Version]
- Hallgrímsson, B.; Miyake, T.; Wilmore, K.; Hall, B.K. Embryological origins of developmental stability: Size, shape and fluctuating asymmetry in prenatal random bred mice. J. Exp. Zool. B 2003, 296, 40–57. [Google Scholar] [CrossRef]
- Knierim, U.; Van Dongen, S.; Forkman, B.; Tuyttens, F.A.M.; Špinka, M.; Campo, J.L.; Weissengruber, G.E. Fluctuating asymmetry as an animal welfare indicator—A review of methodology and validity. Physiol. Behav. 2007, 92, 398–421. [Google Scholar] [CrossRef] [PubMed]
- Torre, I.; Arrizabalaga, A.; Díaz, M. Ratón de campo Apodemus sylvaticus. Galemys 2002, 14, 1–26. [Google Scholar]
- Clarke, G.M. Relationships between developmental stability and fitness: Application for conservation biology. Conserv. Biol. 1995, 9, 18–24. [Google Scholar] [CrossRef]
- Galeotti, P.; Vicario, V. Fluctuating asymmetry in body traits increases predation risks: Tawny owl selection against asymmetric woodmice. Evol. Ecol. 2005, 19, 405–418. [Google Scholar] [CrossRef]
- Tattersall, F.H.; Macdonald, D.W.; Hart, B.J.; Manley, W.J. Balanced dispersal or source–sink–do both models describe wood mice in farmed landscapes? Oikos 2004, 106, 536–550. [Google Scholar] [CrossRef]
- Rosalino, L.M.; Ferreira, D.; Leitão, I.; Santos-Reis, M. Usage patterns of Mediterranean agro-forest habitat components by wood mice Apodemus sylvaticus. Mamm. Biol. 2011, 76, 268–273. [Google Scholar] [CrossRef]
- Hale, R.; Swearer, S.E. Ecological traps: Current evidence and future directions. Proc. R. Soc. B 2016, 283, 20152647. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Trait | Measurement Error (%) | Repeatability | Mean (μm) | SD (μm) | Kolmogorov–Smirnov | p (Mean = 0) |
|---|---|---|---|---|---|---|
| L-R M1 | 2.359 | 0.976 | 171.3 | 392.5 | p > 0.20 | 0.082 |
| L-R M2 | 0.037 | 1.000 | 11.1 | 144.5 | p > 0.20 | 0.750 |
| L-R M3 | 13.679 | 0.863 | −48.9 | 127.3 | p > 0.20 | 0.122 |
| L-R M4 | 0.011 | 1.000 | 108.1 | 219.5 | p > 0.20 | 0.052 |
| L-R M5 | 16.363 | 0.836 | −4.3 | 192.1 | p > 0.20 | 0.926 |
| L-R M6 | 0.004 | 1.000 | −44.5 | 167.3 | p > 0.20 | 0.276 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz, M.; Morán-López, T. Forest Fragmentation and Developmental Stability of Wood Mice Apodemus sylvaticus: A Food-Mediated Effect? Diversity 2023, 15, 423. https://doi.org/10.3390/d15030423
Díaz M, Morán-López T. Forest Fragmentation and Developmental Stability of Wood Mice Apodemus sylvaticus: A Food-Mediated Effect? Diversity. 2023; 15(3):423. https://doi.org/10.3390/d15030423
Chicago/Turabian StyleDíaz, Mario, and Teresa Morán-López. 2023. "Forest Fragmentation and Developmental Stability of Wood Mice Apodemus sylvaticus: A Food-Mediated Effect?" Diversity 15, no. 3: 423. https://doi.org/10.3390/d15030423
APA StyleDíaz, M., & Morán-López, T. (2023). Forest Fragmentation and Developmental Stability of Wood Mice Apodemus sylvaticus: A Food-Mediated Effect? Diversity, 15(3), 423. https://doi.org/10.3390/d15030423