

The Fate of Guzmania monostachia in Florida Rests with Humans


Abstract
:1. Introduction
1.1. Guzmania monostachia in Florida
1.2. History of Ecosystems in Modern-Day Florida and Threats to Bromeliad Habitats
1.3. Conserving Bromeliads in Florida
2. Materials and Methods
2.1. Occurrence Data Compilation
2.2. Ecological Variables Assembly
2.3. Ecological Niche Models
2.4. Identifying Areas of Suitable Habitat
3. Results
3.1. Occurrence Data
3.2. Ecological Niche Models
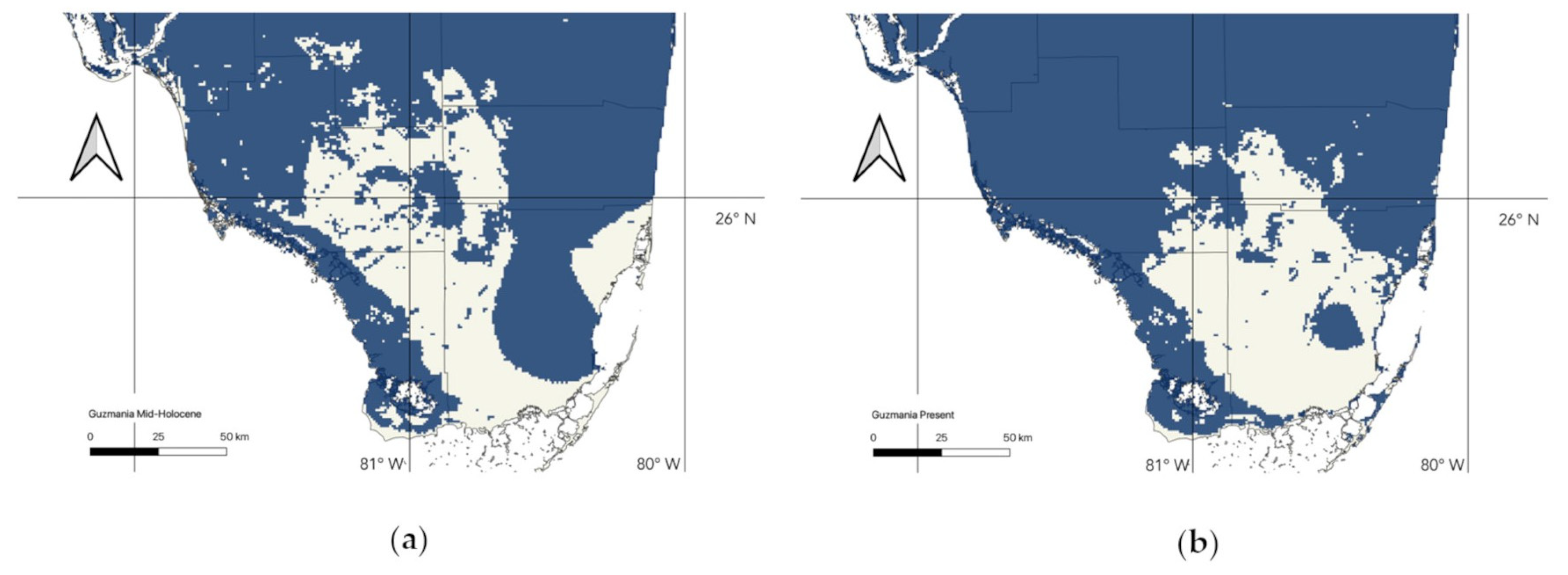
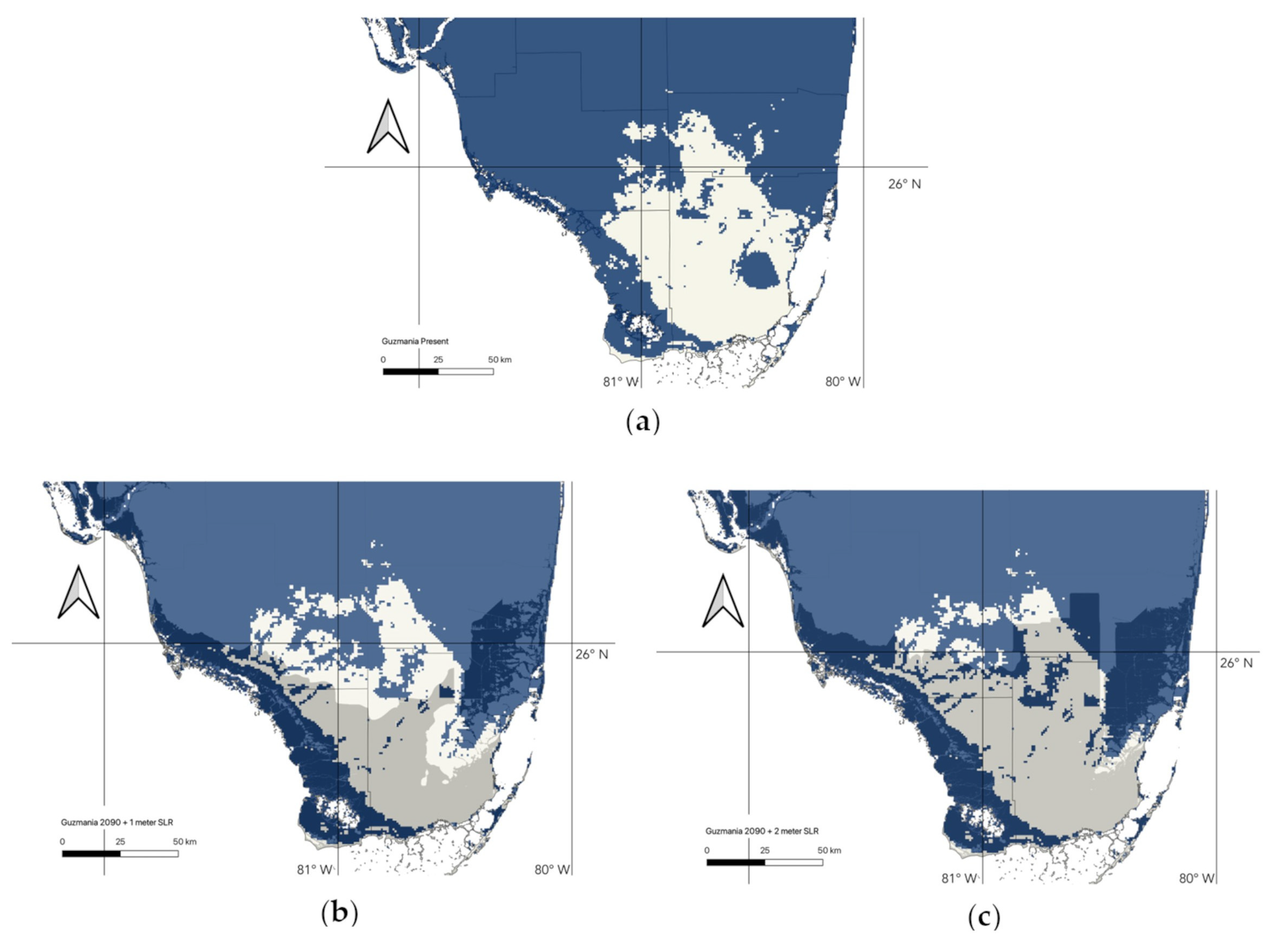
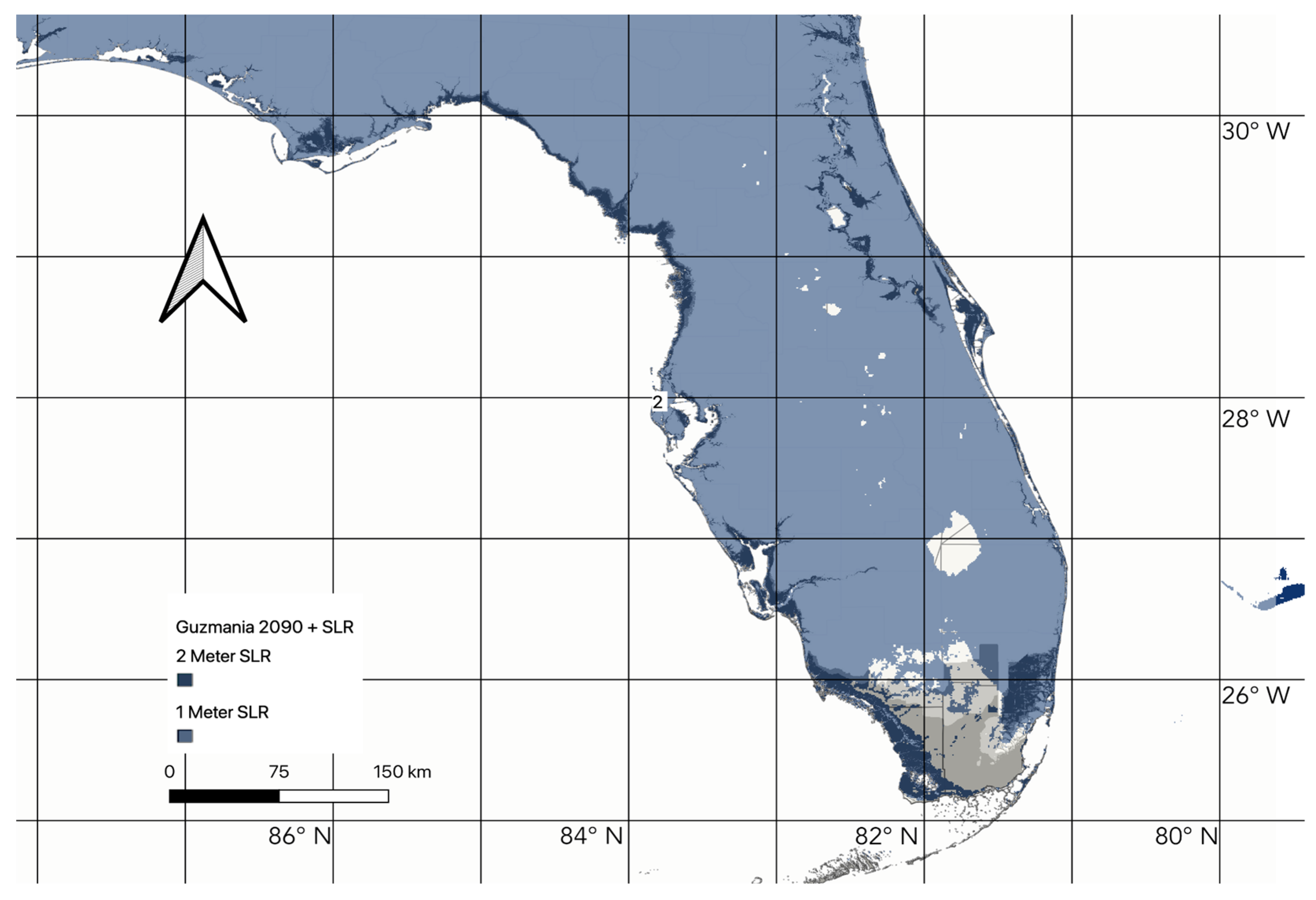
3.3. Changes in Suitable Habitat
4. Discussion
4.1. Habitat Suitability in the Future under Climate Change
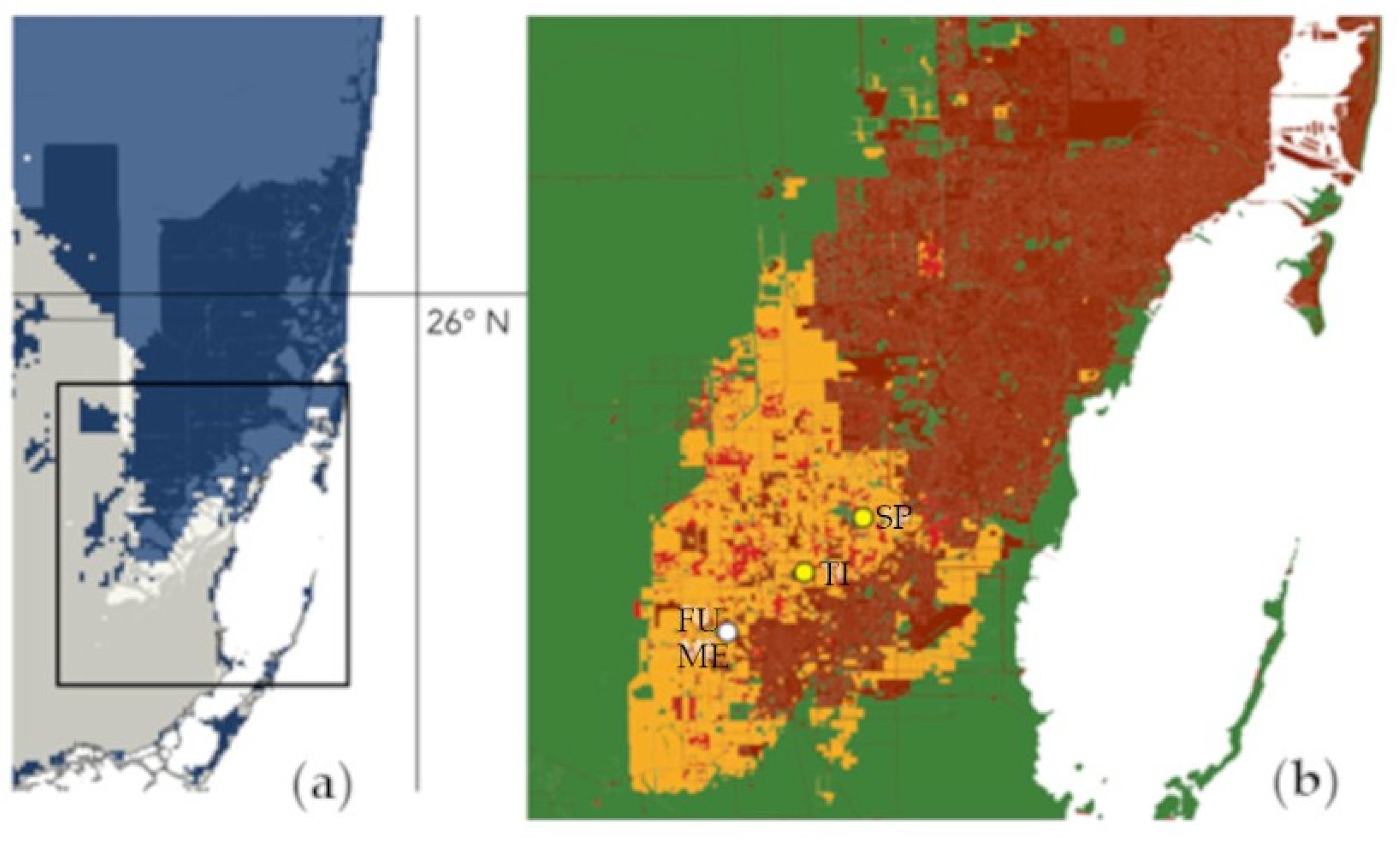
4.2. Implications for Conservation and Future Steps
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Barfuss, M.H.J.; Till, W.; Leme, E.M.C.; Pinzón, J.P.; Manzanares, J.M.; Halbritter, H.; Samuel, R.; Brown, G.K. Taxonomic revision of Bromeliaceae subfam. Tillandsioideae based on a multi-locus DNA sequence phylogeny and morphology. Phytotaxa 2016, 279, 1. [Google Scholar] [CrossRef]
- Givnish, T.; Millam, K.; Berry, P.; Sytsma, K. Phylogeny, Adaptive Radiation, and Historical Biogeography of Bromeliaceae Inferred from ndhF Sequence Data. Aliso 2007, 23, 3–26. [Google Scholar] [CrossRef]
- Givnish, T.J.; Barfuss, M.H.J.; Van Ee, B.; Riina, R.; Schulte, K.; Horres, R.; Gonsiska, P.A.; Jabaily, R.S.; Crayn, D.M.; Smith, J.A.C.; et al. Phylogeny, adaptive radiation, and historical biogeography in Bromeliaceae: Insights from an eight-locus plastid phylogeny. Am. J. Bot. 2011, 98, 872–895. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.H.; Thomas, M.C. Metamasius callizona (CHEVROLAT) (COLEOPTERA: CURCULIONIDAE), AN IMMIGRANT PEST, DESTROYS BROMELIADS IN FLORIDA. Can. Entomol. 1994, 126, 673–682. [Google Scholar] [CrossRef]
- Frank, J.H.; Cave, R.D. Metamasius callizona is destroying Florida’s native bromeliads. In Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 2–16 September 2005. [Google Scholar]
- Frank, J.H. Exploration in Guatemala and Belize for more parasitoids to use against Metamasius callizona in Florida. J. Bromel. Soc. 2011, 61, 112–125. [Google Scholar]
- Sunquist-Blunden, C.; Montero-McAllister, N. Florida’s Endangered and Threatened Species; Florida Fish and Wildlife Conservation Commission: Tallahassee, FL, USA, 2022. [Google Scholar]
- Gann, G.D.; Stocking, G.C. Collaborators Floristic Inventory of South Florida Database Online. Available online: https://regionalconservation.org/ircs/database/database.asp (accessed on 22 September 2022).
- Krupar, S.L. (Department of Biology, University of Florida, Gainesville, FL, USA) Personal communication, 2022.
- Benzing, D.H. Bromeliaceae: Profile of An Adaptive Radiation; Cambridge University Press: Cambridge, UK, 2000; ISBN 0521430313. [Google Scholar]
- Cascante-Marín, A.; de Jong, M.; Borg, E.D.; Oostermeijer, J.G.B.; Wolf, J.H.D.; den Nijs, J.C.M. Reproductive strategies and colonizing ability of two sympatric epiphytic bromeliads in a tropical premontane area. Int. J. Plant Sci. 2006, 167, 1187–1195. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Peñuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef]
- Hine, A.C. Geologic History of Florida, 2nd ed.; University Press of Florida: Gainesville, FL, USA, 2013. [Google Scholar]
- Gardulski, A.F.; Gowen, M.H.; Milsark, A.; Weiterman, S.D.; Wise, S.W.; Mullins, H.T. Evolution of a deep-water carbonate platform: Upper Cretaceous to Pleistocene sedimentary environments on the west Florida margin. Mar. Geol. 1991, 101, 163–179. [Google Scholar] [CrossRef]
- Missimer, T.M.; Maliva, R.G. Late Miocene fluvial sediment transport from the southern Appalachian Mountains to southern Florida: An example of an old mountain belt sediment production surge. Sedimentology 2017, 64, 1846–1870. [Google Scholar] [CrossRef] [Green Version]
- Dahl, T.E. Florida’s Wetlands: An Update on Status and Trends 1985 to 1996; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 2005. [Google Scholar]
- McLaughlin, D.L.; Cohen, M.J. Realizing ecosystem services: Wetland hydrologic function along a gradient of ecosystem condition. Ecol. Appl. 2013, 23, 1619–1631. [Google Scholar] [CrossRef]
- Dahl, T.E. Wetlands Losses in the United States 1780’s to 1980’s; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1990. [Google Scholar]
- Todd, M.J.; Muneepeerakul, R.; Pumo, D.; Azaele, S.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydrological drivers of wetland vegetation community distribution within Everglades National Park, Florida. Adv. Water Resour. 2010, 33, 1279–1289. [Google Scholar] [CrossRef]
- Kushlan, J.A. External threats and internal management: The hydrologic regulation of the Everglades, Florida, USA. Environ. Manag. 1987, 11, 109–119. [Google Scholar] [CrossRef]
- Todd, M.J.; Muneepeerakul, R.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Possible climate change impacts on the hydrological and vegetative character of Everglades National Park, Florida. Ecohydrology 2012, 5, 326–336. [Google Scholar] [CrossRef]
- Comer, P.; Faber-Langendoen, D.; Evans, R.; Gawler, S.; Josse, C.; Kittel, G.; Menard, S.; Pyne, M.; Reid, M.; Schulz, K.; et al. Ecological Systems of the United States: A Working Classification of U.S. Terrestrial Systems; NatureServe: Arlington, VA, USA, 2003. [Google Scholar]
- Meeder, J.F.; Harlem, P.W. Origin and development of true karst valleys in response to late Holocene sea-level change, the Transverse Glades of southeast Florida, USA. Depos. Rec. 2019, 5, 558–577. [Google Scholar] [CrossRef]
- Brooks, W.R.; Lockwood, J.L.; Jordan, R.C. Tropical paradox: A multi-scale analysis of the invasion paradox within Miami Rock Ridge tropical hardwood hammocks. Biol. Invasions 2013, 15, 921–930. [Google Scholar] [CrossRef]
- Owen, M. (Park Biologist, Fakahatchee Strand Preserve State Park, Copeland, FL, USA) Personal communication, 2021.
- Frank, J.H. Bromeliad-Eating Weevils. Selbyana 1999, 20, 40–48. [Google Scholar]
- Rifat, S.A.A.; Liu, W. Quantifying spatiotemporal patterns and major explanatory factors of urban expansion in miami metropolitan area during 1992–2016. Remote Sens. 2019, 11, 2493. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.; Solecki, W. Theorizing Land-Cover and Land-Use Change: The Case of the Florida Everglades and Its Degradation. Ann. Assoc. Am. Geogr. 2004, 94, 311–328. [Google Scholar] [CrossRef]
- Cooper, T.M.; Frank, J.H.; Cave, R.D. Loss of phytotelmata due to an invasive bromeliad-eating weevil and its potential effects on faunal diversity and biogeochemical cycles. Acta Oecol. 2014, 54, 51–56. [Google Scholar] [CrossRef]
- Van Rossum, F.; Vekemans, X.; Gratia, E.; Meerts, P. A comparative study of allozyme variation of peripheral and central populations of Silene nutans L. (Caryophyllaceae) from Western Europe: Implications for conservation. Plant Syst. Evol. 2003, 242, 49–61. [Google Scholar] [CrossRef]
- Hamilton, J.A.; Eckert, C.G. Population genetic consequences of geographic disjunction: A prairie plant isolated on Great Lakes alvars. Mol. Ecol. 2007, 16, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.Y.; Van Der Marel, R.C.; Starzomski, B.M. Climate Change and Conservation of Leading-Edge Peripheral Populations. Conserv. Biol. 2009, 23, 1369–1373. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting species’ geographic distributions based on ecological niche modeling. Condor 2001, 103, 599–605. [Google Scholar] [CrossRef]
- Kolanowska, M.; Rewicz, A.; Baranow, P. Ecological niche modeling of the pantropical orchid Polystachya concreta (Orchidaceae) and its response to climate change. Sci. Rep. 2020, 10, 14801. [Google Scholar] [CrossRef]
- Naranjo, A.A.; Melton, A.E.; Soltis, D.E.; Soltis, P.S. Endemism, projected climate change, and identifying species of critical concern in the Scrub Mint clade (Lamiaceae). Conserv. Sci. Pract. 2022, 4, e621. [Google Scholar] [CrossRef]
- Martínez-Meyer, E.; Peterson, A.T. Conservatism of ecological niche characteristics in North American plant species over the Pleistocene-to-Recent transition. J. Biogeogr. 2006, 33, 1779–1789. [Google Scholar] [CrossRef]
- Rios, N.E.; Bart, H.L. GEOLocate; Tulane University Museum of Natural History: Belle Chasse, LA, USA, 2010. [Google Scholar]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Package “dismo”. Circles 2017, 9, 1–68. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Batjes, N.H.; Ribeiro, E.; van Oostrum, A. Standardised soil profile data to support global mapping and modelling (WoSIS snapshot 2019). Earth Syst. Sci. Data 2020, 12, 299–320. [Google Scholar] [CrossRef] [Green Version]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PloS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, H.T.; Lovett-Doust, J. Accounting for regional niche variation in habitat suitability models. Oikos 2007, 116, 99–110. [Google Scholar] [CrossRef]
- Quesada, C.A.; Phillips, O.L.; Schwarz, M.; Czimczik, C.I.; Baker, T.R.; Patiño, S.; Fyllas, N.M.; Hodnett, M.G.; Herrera, R.; Almeida, S.; et al. Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate. Biogeosciences 2012, 9, 2203–2246. [Google Scholar] [CrossRef] [Green Version]
- Zuquim, G.; Costa, F.R.C.; Tuomisto, H.; Moulatlet, G.M.; Figueiredo, F.O.G. The importance of soils in predicting the future of plant habitat suitability in a tropical forest. Plant Soil 2020, 450, 151–170. [Google Scholar] [CrossRef] [Green Version]
- Ngoh, M.L. Evaluating Current and Future Potential Distribution of Epiphytic Orchids in the Congo Basin with Ecological Niche Models. Master’s Thesis, University of Tennessee, Knoxville, Tennessee, 2022. [Google Scholar]
- Bennett, B.C. Spatial distribution of catopsis and guzmania (bromeliaceae) in southern florida. Bull. Torrey Bot. Club 1987, 114, 265. [Google Scholar] [CrossRef]
- Hausfather, Z.; Peters, G.P. Emissions—The “business as usual” story is misleading. Nature 2020, 577, 618–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweet, W.V.; Kopp, R.; Weaver, C.P.; Obeysekera, J.; Horton, R.M.; Thieler, E.R.; Zervas, C.E. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Ocean Service, Center for Operational Oceanographic Products and Services. In Global and Regional Sea Level Rise Scenarios for the United States; NOAA: Washington, DC, USA, 2017. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers (USACE). Sea-Level Change Curve Calculator. Available online: https://cwbi-app.sec.usace.army.mil/rccslc/slcc_calc.html (accessed on 26 January 2023).
- Bivand, R.; Rundel, C.; Pebesma, E.; Stuetz, R.; Hufthammer, K.O.; Bivand, M.R. Package “Rgeos”; The Comprehensive R Archive Network (CRAN): San Francisco, CA, USA, 2017. [Google Scholar]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Modell. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Romero-Alvarez, D.; Escobar, L.E.; Varela, S.; Larkin, D.J.; Phelps, N.B.D. Forecasting distributions of an aquatic invasive species (Nitellopsis obtusa) under future climate scenarios. PloS ONE 2017, 12, e0180930. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, R.L.; Soltis, D.E.; Cellinese, N. The future of cold-adapted plants in changing climates: Micranthes (Saxifragaceae) as a case study. Ecol. Evol. 2018, 8, 7164–7177. [Google Scholar] [CrossRef] [Green Version]
- van Proosdij, A.S.J.; Sosef, M.S.M.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- McPHERSON, J.M.; Jetz, W.; Rogers, D.J. The effects of species’ range sizes on the accuracy of distribution models: Ecological phenomenon or statistical artefact? J. Appl. Ecol 2004, 41, 811–823. [Google Scholar] [CrossRef]
- Leroy, B.; Meynard, C.N.; Bellard, C.; Courchamp, F. virtualspecies, an R package to generate virtual species distributions. Ecography 2016, 39, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. Ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Escalante, T.; Rodríguez-Tapia, G.; Linaje, M.; Illoldi-Rangel, P.; González-López, R. Identification of areas of endemism from species distribution models: Threshold selection and Nearctic mammals. TIP 2013, 16, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Van Etten, J.; Cheng, J.; Mattiuzzi, M.; Summer, M.; Greenburg, J.A.; Hijmans, M.R.J. Package “Raster”; R Package: Vienna, Austria, 2015. [Google Scholar]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Ecol. Modell. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Veloz, S.D. Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J. Biogeogr. 2009, 36, 2290–2299. [Google Scholar] [CrossRef]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberón, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E.; et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Modell. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Soberon, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodiv. Inf. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. S2), 19729–19736. [Google Scholar] [CrossRef] [Green Version]
- Wagner, K.; Zotz, G. Including dynamics in the equation: Tree growth rates and host specificity of vascular epiphytes. J. Ecol. 2020, 108, 761–773. [Google Scholar] [CrossRef]
- Vasconcelos, T.S.; Antonelli, C.P.; Napoli, M.F. Mutualism influences species distribution predictions for a bromeliad-breeding anuran under climate change. Austral Ecol. 2017, 42, 869–877. [Google Scholar] [CrossRef]
- Westwood, M.; Oldfield, S.; Jerome, D.; Romero-Severson, J. Fraxinus caroliniana. The IUCN Red List of Threatened Species 2017: e.T63004A96445289. Available online: https://www.iucnredlist.org/species/63004/96445289(accessed on 23 February 2023).
- Lippert, A.P.U.; Silva, V.L.; Mallmann, I.T.; Müller, A.; Droste, A.; Schmitt, J.L. Edge effect on vascular epiphytes in a subtropical Atlantic Forest fragment. J. Env. Anal. Progr. 2022, 7, 135–149. [Google Scholar] [CrossRef]
- The Florida Cooperative Land Cover Map; 3.5.; Florida Fish and Wildlife Conservation Commission and Florida Natural Areas Inventory. 2021. Available online: https://myfwc.com/research/gis/wildlife/cooperative-land-cover/(accessed on 31 October 2022).
- Possley, J. (South Florida Conservation Program Manager, Fairchild Tropical Botanic Garden, Miami, FL, USA) Personal communication, 2021.
- Wintergerst, S. Fairchild Tropical Botanic Garden’s Conservation Program on-site Frozen Seed Bankinventory. Available online: https://fairchildgarden.org/science-and-education/science/native-plant-conservation/ (accessed on 23 February 2023).
- Negrelle, R.R.B.; Mitchell, D.; Anacleto, A. Bromeliad ornamental species: Conservation issues and challenges related to commercialization. Acta Sci. Biol. Sci. 2012, 34, 91–100. [Google Scholar] [CrossRef]
- Ismail, S.A.; Pouteau, R.; van Kleunen, M.; Maurel, N.; Kueffer, C. Horticultural plant use as a so-far neglected pillar of ex situ conservation. Conserv. Lett. 2021, 14, e12825. [Google Scholar] [CrossRef]
- Million Orchid Project—Fairchild Tropical Botanic Garden. Available online: https://fairchildgarden.org/science-and-education/science/million-orchid-project/ (accessed on 23 February 2023).
- Aubin, I.; Garbe, C.M.; Colombo, S.; Drever, C.R.; McKenney, D.W.; Messier, C.; Pedlar, J.; Saner, M.A.; Venier, L.; Wellstead, A.M.; et al. Why we disagree about assisted migration: Ethical implications of a key debate regarding the future of Canada’s forests. For. Chron. 2011, 87, 755–765. [Google Scholar] [CrossRef]
- Schlaepfer, M.A.; Helenbrook, W.D.; Searing, K.B.; Shoemaker, K.T. Assisted colonization: Evaluating contrasting management actions (and values) in the face of uncertainty. Trends Ecol. Evol. 2009, 24, 471–472. [Google Scholar] [CrossRef]
- Vitt, P.; Havens, K.; Kramer, A.T.; Sollenberger, D.; Yates, E. Assisted migration of plants: Changes in latitudes, changes in attitudes. Biol. Conserv. 2010, 143, 18–27. [Google Scholar] [CrossRef]
- Ricciardi, A.; Simberloff, D. Assisted colonization is not a viable conservation strategy. Trends Ecol. Evol. 2009, 24, 248–253. [Google Scholar] [CrossRef]
- Duarte, M.M.; Gandolfi, S. Diversifying growth forms in tropical forest restoration: Enrichment with vascular epiphytes. For. Ecol. Manag. 2017, 401, 89–98. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Hughes, L.; McIntyre, S.; Lindenmayer, D.B.; Parmesan, C.; Possingham, H.P.; Thomas, C.D. Ecology. Assisted colonization and rapid climate change. Science 2008, 321, 345–346. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, J.S.; Hellmann, J.J.; Schwartz, M.W. A Framework for Debate of Assisted Migration in an Era of Climate Change. Conserv. Biol. 2007, 21, 297–302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mid-Holocene Suitable Area (in km2) | Current Suitable Area (in km2) | Schoener’s D | Present Area/Past Area | |
|---|---|---|---|---|
| G. monostachia (90 and 95 pct) | 16,559.87 | 17,723.16 | 0.83 | 1.07 |
| G. monostachia (90 pct) | 2090 Suitable Area (in km2) | Current Suitable Area (in km2) | Schoener’s D | Future Area/Present Area |
| 0 m SLR | 17,206.83 | 17,723.16 | 0.93 | 0.97 |
| 1 m SLR | 13,180.92 | N/A | 0.93 | 0.74 |
| 2 m SLR | 10,861.82 | N/A | 0.95 | 0.61 |
| G. monostachia (95 pct) | 2090 Suitable Area (in km2) | Current Suitable Area (in km2) | Schoener’s D | Future Area/Present Area |
| 0 m SLR | 20,877.974 | 17,723.16 | 0.84 | 1.17 |
| 1 m SLR | 15,552.71 | N/A | 0.87 | 0.87 |
| 2 m SLR | 11,776.39 | N/A | 0.95 | 0.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupar, S.; Naranjo, A.A.; Godden, G.; Cellinese, N. The Fate of Guzmania monostachia in Florida Rests with Humans. Diversity 2023, 15, 525. https://doi.org/10.3390/d15040525
Krupar S, Naranjo AA, Godden G, Cellinese N. The Fate of Guzmania monostachia in Florida Rests with Humans. Diversity. 2023; 15(4):525. https://doi.org/10.3390/d15040525
Chicago/Turabian StyleKrupar, Shelby, Andre A. Naranjo, Grant Godden, and Nico Cellinese. 2023. "The Fate of Guzmania monostachia in Florida Rests with Humans" Diversity 15, no. 4: 525. https://doi.org/10.3390/d15040525
APA StyleKrupar, S., Naranjo, A. A., Godden, G., & Cellinese, N. (2023). The Fate of Guzmania monostachia in Florida Rests with Humans. Diversity, 15(4), 525. https://doi.org/10.3390/d15040525