
Ancient DNA Contradicts the Presence of Social Voles (Genus Microtus, Subgenus Sumeriomys) in the Late Pleistocene of Western Europe

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Ancient Sample and Morphological Methods
2.2. DNA Extraction, Enrichment and Sequencing
2.3. Sequencing Data Processing
2.4. Phylogenetic Analyses
3. Results
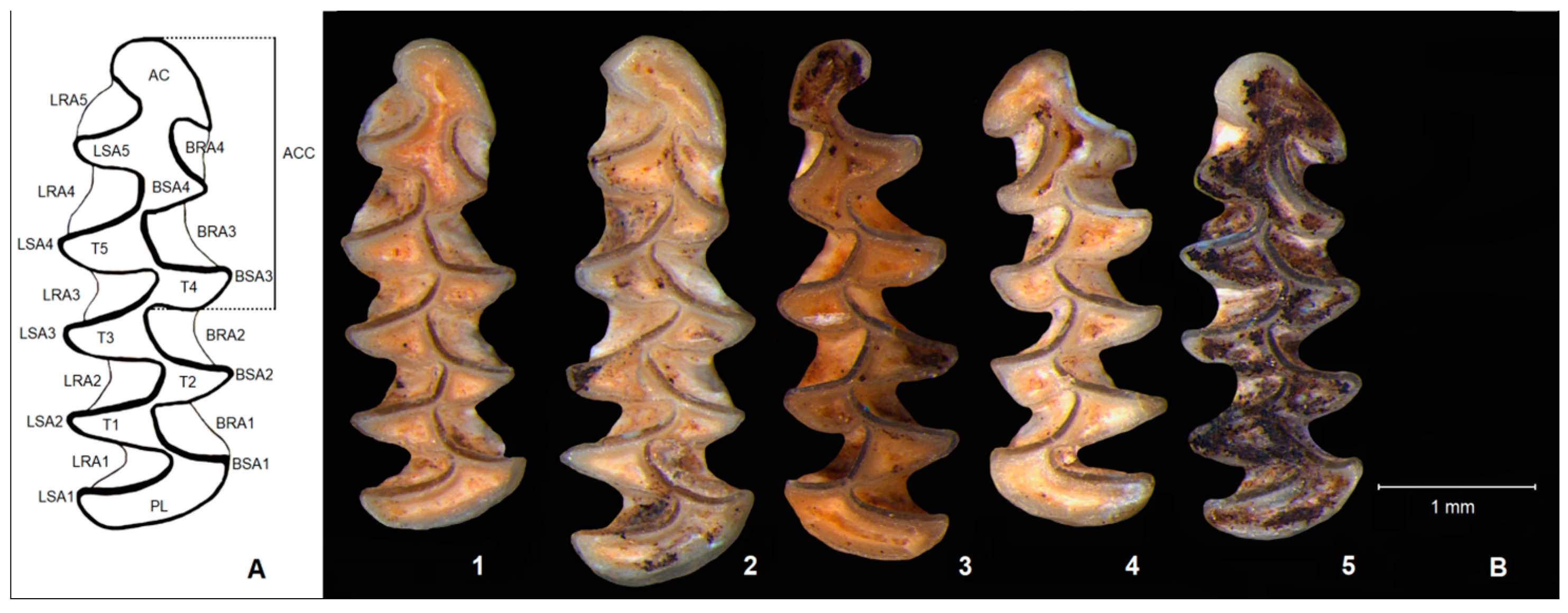
3.1. Morphological Characteristics of M. bifrons
- MI2868 (CVPG 18-371) (right m1, L = 3.04 mm): five triangles (T1–T5); T6 and T7 broadly confluent; BSA 4 less pronounced in comparison with LSA 5; and anterior cap (AC) wide with spur-shaped BSA 5 (Figure 1/1);
- MI2869 (CVPG 47-957) (right m1, L = 3.45 mm, paratype): seven dental triangles (T1–T7); T6 and T7 separated; BSA 4 and LSA 5 almost of the same width; and anterior cap (AC) wide, with spur-shaped BSA 5 (Figure 1/2);
- MI2870 (CVPG 59-1309, left m1, L = 3.22 mm): five dental triangles (T1–T5); T6 and T7 confluent; BSA 4 much less pronounced in comparison with LSA 5; and anterior cap (AC) separated from T6–T7 with small BSA 5 (Figure 1/3);
- MI2871 (CVPG 56-1213, left m1, L = 2.95 mm): five dental triangles (T1–T5); T6 and T7 broadly confluent; BSA 4 pronounced as much as LSA 5; and anterior cap (AC) not fully separated from T6–T7 with pronounced BSA 5 and very small LSA 6 (Figure 1/4);
- MI2872 (CVPG 51-1065, right m1, L = 2.98 mm): five dental triangles (T1–T5); T6 and T7 broadly confluent; BSA 4 pronounced as much as LSA 5; and anterior cap (AC) wide with spur-shaped BSA 5 (Figure 1/5).
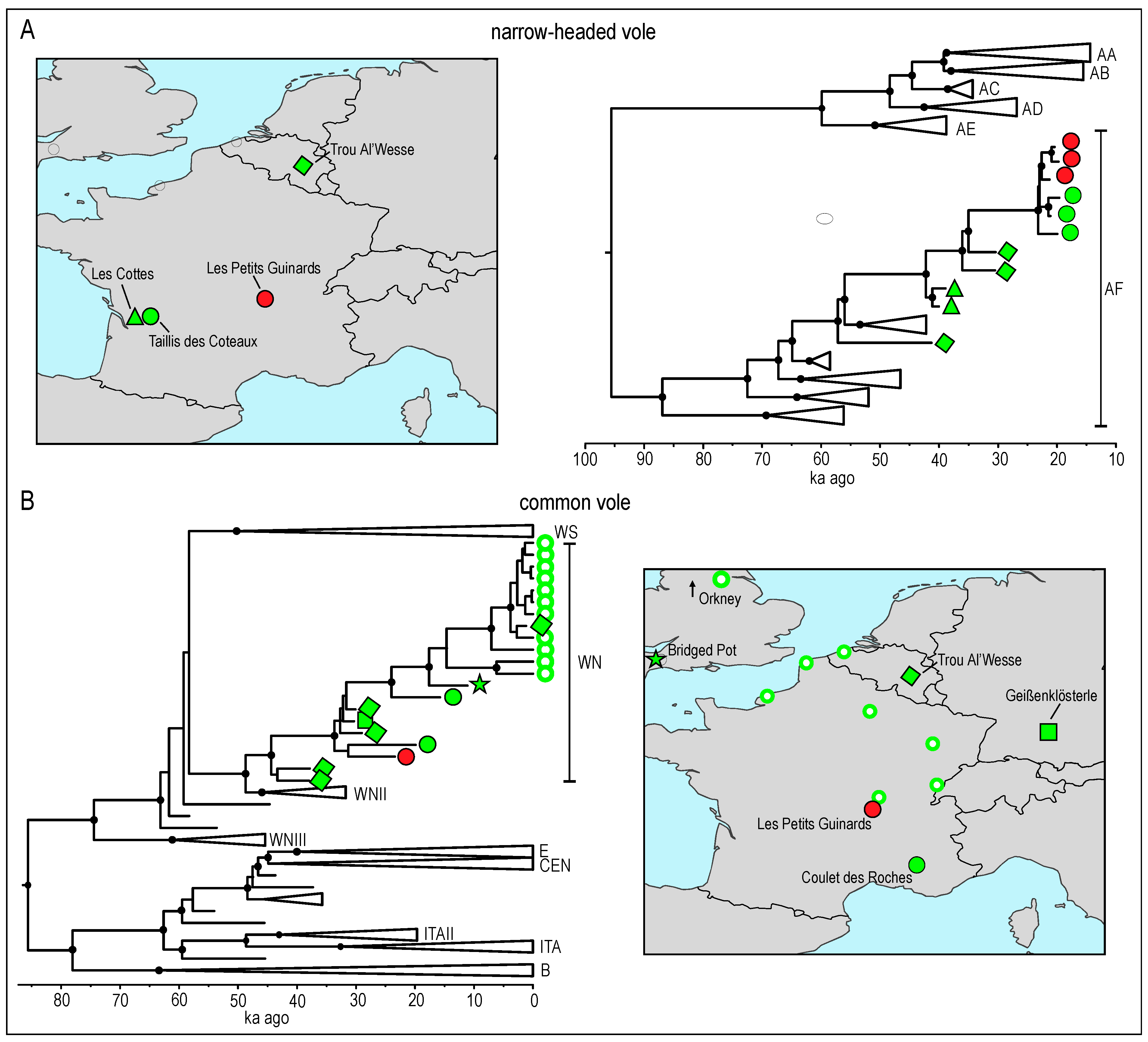
3.2. Phylogenetic Analysis and Molecular Dating
4. Discussion
4.1. Taxonomic Attribution Revealed from Morphological Characteristics and Genetic Research
4.2. Supernumerary Triangles
4.3. Wide and Spur-Shaped Anterior Cap
4.4. Reasons for Incorrect Species Determination Based on Morphological Criteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pardiñas, U.F.J.; Myers, P.; León-Paniagua, L.; Ordóñez Garza, N.; Cook, J.; Kryštufek, B.; Haslauer, R.; Bradley, R.; Shenbrot, G.; Patton, J. Family Cricetidae (true hamsters, voles, lemmings and new world rats and mice). In Handbook of the Mammals of the World, Rodents II; Wilson, D.E., Lacher, T.E., Mittermeier, R.A., Eds.; Lynx Edicions: Barcelona, Spain, 2017; Volume 7, pp. 204–279. [Google Scholar]
- Jaarola, M.; Martínková, N.; Gündüz, İ.; Brunhoff, C.; Zima, J.; Nadachowski, A.; Amori, G.; Bulatova, N.S.; Chondropoulos, B.; Fraguedakis-Tsolis, S.; et al. Molecular phylogeny of the speciose vole genus Microtus (Arvicolinae, Rodentia) inferred from mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2004, 33, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Kryštufek, B.; Shenbrot, G.I. Voles and Lemmings (Arvicolinae) of the Palaearctic Region; University of Maribor Press: Maribor, Slovenia, 2022; p. 436. [Google Scholar]
- Brunet-Lecomte, P.; Nadachowski, A.; Chaline, J. Microtus (Terricola) grafi nov. sp. du pléistocène supérieur de la grotte de Bacho Kiro (Bulgarie). Geobios 1992, 25, 505–509. [Google Scholar] [CrossRef]
- Jeannet, M.; Fontana, L. Microtus (Sumeriomys) bifrons nov. sp. (Rodentia, Mammalia), a new vole in the French Upper Pleistocene identified at the Petits Guinards site (Creuzier-le-Vieux, Allier, France). PALEO Rev. Archéol. Préhist. 2015, 26, 59–77. [Google Scholar] [CrossRef]
- Baca, M.; Popović, D.; Lemanik, A.; Baca, K.; Horáček, I.; Nadachowski, A. Highly divergent lineage of narrow-headed vole from the Late Pleistocene Europe. Sci. Rep. 2019, 9, 17799. [Google Scholar] [CrossRef] [Green Version]
- Baca, M.; Popović, D.; Agadzhanyan, A.K.; Baca, K.; Conard, N.J.; Fewlass, H.; Filek, T.; Golubiński, M.; Horáček, I.; Knul, M.V.; et al. Ancient DNA of narrow-headed voles reveals common features of the Late Pleistocene population dynamics in cold-adapted small mammals. Proc. R. Soc. B 2023, 290, 20222238. [Google Scholar] [CrossRef] [PubMed]
- Baca, M.; Popović, D.; Lemanik, A.; Fewlass, H.; Talamo, S.; Zima, J.; Ridush, B.; Popov, V.; Nadachowski, A. The Tien Shan vole (Microtus ilaeus; Rodentia: Cricetidae) as a new species in the Late Pleistocene of Europe. Ecol. Evol. 2021, 11, 16113–16125. [Google Scholar] [CrossRef]
- Fontana, L.; Lang, L.; Chauvière, F.-X.; Jeannet, M.; Mourer-Chauviré, C.; Magoga, L. Paléolithique supérieur récent du nord du Massif Central: Des données inattendues sur le site des Petits Guinards à Creuzier-le-Vieux (Allier, France). Bull. Préhist. Sud Quest. 2003, 10, 77–93. [Google Scholar]
- Chauvière, F.-X.; Fontana, L.; Land, L.; Bonani, G.; Hajdas, I. Une préhampe magdalénienne en bois du renne aux Petits Guinards (Allier, France). Comptes Rendus Palevol 2006, 5, 725–733. [Google Scholar] [CrossRef]
- Fontana, L.; Chauvière, F.-X. The total exploitation of reindeer at the site of Les Petits Guinards: What’s new about the annual cycle of Magdalenian groups in the French Massif Central? In Search of Total Animal Exploitation: Case Studies from the Upper Palaeolithic and Mesolithic; Fontana, L., Chauvière, F.-X., Bridault, A., Eds.; British Archaeological Reports, International Series 2040; John and Erica Hedges Ltd.: Oxford, UK, 2009; pp. 101–111. [Google Scholar]
- Nadachowski, A. Late Quaternary Rodents of Poland with Special Reference to Morphotype Dentition Analysis of Voles; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1982; p. 109. [Google Scholar]
- Hibbard, C.W. Mammals from the Rexroad Formation from Fox Canyon, Kansas; University of Michigan: Ann Arbor, MI, USA, 1950; Volume 8, pp. 113–192. [Google Scholar]
- Van der Meulen, A.J. Middle Pleistocene smaller mammals from the Monte Peglia (Orveto, Italy) with special reference to the phylogeny of Microtus (Arvicolidae, Rodentia). Quaternaria 1973, 17, 1–144. [Google Scholar]
- Nadachowski, A. Biharian voles (Arvicolidae, Rodentia, Mammalia) from Kozi Grzbiet (Central Poland). Acta Zool. Crac. 1985, 29, 13–28. [Google Scholar]
- Nadachowski, A. Systematics, geographic variation, and evolution of snow voles (Chionomys) based on dental characters. Acta Theriol. 1991, 36, 1–45. [Google Scholar] [CrossRef] [Green Version]
- Rohland, N.; Glocke, I.; Aximu-Petri, A.; Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc. 2018, 13, 2447–2461. [Google Scholar] [CrossRef]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 5, 5448. [Google Scholar] [CrossRef]
- Gansauge, M.T.; Aximu-Petri, A.; Nagel, S.; Meyer, M. Manual and automated preparation of single-stranded DNA libraries for the sequencing of DNA from ancient biological remains and other sources of highly degraded DNA. Nat. Protoc. 2020, 15, 2279–2300. [Google Scholar] [CrossRef] [PubMed]
- Horn, S. Case study: Enrichment of ancient mitochondrial DNA by hybridization capture. Methods Mol. Biol. 2012, 840, 189–195. [Google Scholar] [PubMed]
- Lord, E.; Marangoni, A.; Baca, M.; Popović, D.; Goropashnaya, A.V.; Stewart, J.R.; Knul, M.V.; Noiret, P.; Germonpré, M.; Jimenez, E.-L.; et al. Population dynamics and demographic history of Eurasian collared lemmings. BMC Ecol. Evol. 2022, 22, 126. [Google Scholar] [CrossRef]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid Adapter Trimming, identification, and read merging. BMC Res Notes 2016, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997v2. [Google Scholar]
- Feuerborn, T.R.; Palkopoulou, E.; van der Valk, T.; von Seth, J.; Munters, A.R.; Pečnerová, P.; Dehasque, M.; Ureña, I.; Ersmark, E.; Lagerholm, V.K.; et al. Competitive mapping allows for the identification and exclusion of human DNA contamination in ancient faunal genomic datasets. BMC Genom. 2020, 21, 844. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Gangavarapu, K.; Quick, J.; Matteson, N.L.; de Jesus, J.G.; Main, B.J.; Tan, A.L.; Paul, L.M.; Brackney, D.E.; Grewal, S.; et al. An amplicon-based sequencing framework for accurately measuring intrahost virus diversity using PrimalSeq and IVar. Genome Biol. 2019, 20, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using tablet for visual exploration of second-generation sequencing data. Brief Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Jónsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.F.; Orlando, L. MapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef] [PubMed]
- Baca, M.; Popović, D.; Lemanik, A.; Bañuls-Cardona, S.; Conard, N.J.; Cuenca-Bescós, G.; Desclaux, E.; Fewlass, H.; Garcia, J.T.; Hadravova, T.; et al. Ancient DNA reveals interstadials as a driver of common vole population dynamics during the last glacial period. J. Biogeogr. 2023, 50, 183–196. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, B.; Ho, S.Y.W.; Drummond, A.J.; Suchard, M.A.; Pybus, O.G.; Rambaut, A.A. Bayesian Phylogenetic Method to estimate unknown sequence ages. Mol. Biol. Evol. 2011, 28, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Abramson, N.I.; Bodrov, S.Y.; Bondareva, O.V.; Genelt-Yanovskiy, E.A.; Petrova, T.V. A mitochondrial genome phylogeny of voles and lemmings (Rodentia: Arvolinae): Evolutionary and taxonomic implications. PLoS ONE 2021, 16, e0248198. [Google Scholar] [CrossRef]
- Ognev, S.I. Mammals of the USSR and Adjacent Countries; Izd. AN SSSR, Moskva: Leningrad, Russia, 1950; Volume 7, p. 706. (In Russian) [Google Scholar]
- Golenishchev, F.N.; Sablina, O.V.; Borodin, P.M.; Gerasimov, S. Taxonomy of voles of the subgenus Sumeriomys Argyropulo, 1933 (Rodentia, Arvicolinae, Microtus). Russ. J. Theriol. 2002, 1, 43–55. [Google Scholar] [CrossRef]
- Tchernov, E. Succession of Rodents Fauna during the Upper Pleistocene of Israel; Verlag Paul Parey: Hamburg, Germany; Berlin, Germany, 1968; p. 152. [Google Scholar]
- Aşan Baymedir, N.; Duman, L. Molar pattern in Microtus guentheri (Danford and Alston, 1880) (Mammalia: Rodentia) from Kirikkale Province. J. Appl. Biol. Sci. 2009, 3, 47–53. [Google Scholar]
- Chaline, J. Les Rongeurs du Pléistocène Moyen et Supérieur de France: (Systématique, Biostratigraphie, Paléoclimatologie); Cahiers Paléontologie: Paris, France, 1972; p. 410. [Google Scholar]
- Rekovets, L.I. New subspecies of narrow-headed vole (Microtus gregalis) from the Upper Pleistocene deposits of Ukraine. Dokl. AN USSR. Ser. Biol. 1978, 6, 559–563. (In Ukrainian) [Google Scholar]
- Bolshakov, V.N.; Vasilyeva, I.A.; Maleeva, A.G. Morphotypical Variability of Voles’ Dentition; Nauka Publisher: Moscow, Russia, 1980; p. 140. (In Russian) [Google Scholar]
- Kochev, V.A. Species criteria for M1 molars of Microtus agrestis, M. arvalis, M. oeconomus, M. gregalis, M. middendorfi and M. hyperboreus. Vestn. Zool. 1986, 3, 40–45. (In Russian) [Google Scholar]
- Smirnov, N.G.; Bolshakov, V.N.; Borodin, A.V. Pleistocene Rodents of Northern Western Siberia; Nauka Publishers: Moscow, Russia, 1986; p. 144. (In Russian) [Google Scholar]
- Rekovets, L.I.; Nadachowski, A. Pleistocene voles (Arvicolidae) of the Ukraine. Paleontol. Evol. 1995, 28, 145–245. [Google Scholar]
- Válóczi, T. A Vaskapu-barlang (Bükk hegység) felső-pleisztocén faunájának vizsgálata. Folia Hist. Nat. Musei Matra. 1998, 23, 79–96. (In Hungarian) [Google Scholar]
- Dupal, T.A. Geografic variability and sytematics of the subspecies in the narrow-skulled vole Microtus (Stenocranius) gregalis (Rodentia, Cricetidae). Zool. Zhurnal 2000, 79, 851–858. (In Russian) [Google Scholar]
- Borodin, A.W. Identification Guide for Vole’s Teeth of the Urals and Western Siberia (Late Pleistocene—Recent); Ural Branch of RAS: Yekaterinburg, Russia, 2009; p. 100. (In Russian) [Google Scholar]
- Dupal, T.A.; Abramov, S.A. Intrapopulation morphological varation of the narrow-skulled vole (Microtus gregalis, Rodentia, Arvicolinae). Zool. Zhurnal 2010, 80, 850–861. (In Russian) [Google Scholar]
- Markova, E.A.; Smirnov, N.G.; Kourova, T.P.; Kropacheva, Y.E. Ontogenetic variation in occlusal shape of evergrowing molars in voles: An intravital study in Microtus gregalis (Arvicolidae, Rodentia). Mamm. Biol. 2013, 78, 251–257. [Google Scholar] [CrossRef]
- Ponomarev, D.; Puzachenko, A. Changes in the morphology and morphological diversity of the first lower molar of narrow-headed voles (Microtus gregalis, Arvicolinae, Rodentia) from northeastern European Russia since the Late Pleistocene. Quat. Int. 2017, 436, 239–252. [Google Scholar] [CrossRef]
- Fadeeva, T.V.; Kosintsev, P.A.; Gimranov, D.O.; Yakovlev, A.G. The finding of molars of the archaic vole Lasiopodomys (Stenocranius) gregaloides (Hinton, 1923) (Mammalia, Rodentia, Cricetidae) in the Late Pleistocene of the Southrn Urals. Dokl. Biol. Sci. 2022, 505, 105–108. [Google Scholar] [CrossRef]
- Klimowicz, M.; Nadachowski, A.; Lemanik, A.; Socha, P. Is enamel differentiation quotient (SDQ) of the narrow-headed vole (Microtus gregalis) useful for the Pleistocene biostratigraphy? Quat. Inter. 2016, 420, 348–356. [Google Scholar] [CrossRef]
- Jánossy, D.; Schmidt, E. Extreme Varianten des M1 der Feldmaus (Microtus arvalis Pallas) in Ungarn. II. Z. Säugetierkd. 1975, 40, 34–36. [Google Scholar]
- Kapischke, H.-J.; Kraft, R.; Jentzsch, M.; Hiermeier, M. Variation and complexity of the enamel pattern in the first lower molar of the Field vole, Microtus agrestis (L., 1761) (Mammalia: Rodentia: Arvicolinae). Vertebr. Zool. 2009, 59, 191–195. [Google Scholar] [CrossRef]
- Dienske, H. Notes on differences between sime external and skull characters of Microtus arvalis (Pallas, 1778) and of Microtus agrestis (Linnaeus, 1761) from the Netherlands. Zool. Meded. 1969, 44, 83–108. [Google Scholar]
- Jentzsch, M. Zur Variabilität der Molarenmuster einer Population von Erdmäusen Microtus agrestis (L., 1761) aus dem Norden Sachsen-Anhalts (Mammalia: Rodentia: Arvicolidae). Zool. Abh. 2006, 55, 191–198. [Google Scholar]
- Nadachowski, A. Comments on variation, evolution and phylogeny of Chionomys (Arvicolidae). In Proceedings of the International Symposium on Evolution, Phylogeny and Biostratygraphy of Arvicolids (Rodentia, Mammalia), Rohanov, Czechoslovakia, May 1987; Fejfar, O., Heinrich, W.-D., Eds.; Geological Survey: Prague, Czech Republic, 1990; pp. 353–368. [Google Scholar]
- Popova, L.V.; Nezdolyii, Y.S.; Syniavska, I.; Rekovets, L.; Krokhmal, O.; Mironchuk, T.; Dzeverin, I. Spatial and temeporal patterns of species replacement in the Middle Pleistocene: A case study of Microtus nivaloides Major, 1902 and morphologically related species of the Northern Black Sea and Azov areas. J. Quat. Sci. 2022, 37, 1229–1245. [Google Scholar] [CrossRef]
- Markova, E.; Borodin, A. An advanced form of Microtus nivaloides Forsyth Major, 1902 (Arvicolinae, Rodentia) in the late Middle Pleistocene of West Siberia: Facts and hypotheses. Hist. Biol. 2022, 1–17. [Google Scholar] [CrossRef]
- Popov, V.V.; Marinska, M. An almost one million year long (Early to Late Pleistocene) small mammal succession from the archaeological layers of Kozarnika Cave in Northern Bulgaria. Cour. Forsch. Senckenberg 2007, 259, 79–92. [Google Scholar]
- Markova, A.K. Pleistocene Rodents of the Russian Plain (Their Paleogeographic and Stratigraphic Implications); Nauka Publisher: Moscow, Russia, 1982; p. 186. (In Russian) [Google Scholar]
- Hinton, M.A.C. Monograph of the Voles and Lemmings (Microtinae) Living and Extinct; Richard Clay & Sons: Bungay, UK, 1926; p. 488. [Google Scholar]
- Ivanov, M.; Vöröš, D. Middle Pleistocene voles and lemmings (Rodentia: Arvicolinae) from Za Hájovnou Cave (Javoříčko Karst). Acta Mus. Nat. Pragae Ser. B Hist. Nat. 2014, 70, 43–54. [Google Scholar] [CrossRef]
- Popova, L.V.; Nezdolyii, Y.S.; Krokhmal, O.I.; Rekovets, L.I. Appearance of Microtus agrestis in the territory of Ukraine in the Middle Pleistocene. Geo&Bio 2021, 20, 102–116. (In Ukrainian) [Google Scholar]
- Luzi, E.; López-García, J.M.; Blasco, R.; Rivals, F. Variation in Microtus arvalis and Microtus agrestis (Arvicolinae, Rodentia) dental morphologies in an archaeological context: The case of Teixoneres Cave (Late Pleistocene, North-Eastern Iberia). J. Mamm. Evol. 2017, 24, 495–503. [Google Scholar] [CrossRef]
- Luzi, E.; Pazonyi, P.; López-García, J.M. The influence of climate on morphometric traits of fossil populations of Microtus arvalis and M. agrestis from the Carpathian Basin, northern Hungary. Lethaia 2019, 52, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Bogićević, K.; Nenadić, D.; Mihailović, D. Late Pleistocene voles (Arvicolinae, Rodentia) from the Baranica Cave (Serbia). Geol. Carpathica 2012, 63, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Rörig, G.; Börner, C. Studien über das Gebiss mitteleuropäischer Mäuse. Arb. Kaiserl. Biol. Anst. Land-Forst-Wirtsch. Berl. 1905, 5, 33–89. [Google Scholar]
- Uhlíková, J. Epigenetic and dental variation of the common vole, Microtus arvalis (Mammalia: Rodentia) in the Czech Republic. Folia Zool. 2004, 53, 157–170. [Google Scholar]
- Popov, V.V. Middle Pleistocene small mammals (Insectivora, Lagomorpha, Rodentia) from Morovitsa Cave (North Bulgaria). Acta Zool. Crac. 1989, 32, 561–588. [Google Scholar]
- Nadachowski, A. Morphomertic variability of dentition of the Late Pleistocene voles (Arvicolidae, Rodentia) from Bacho Kiro Cave (Bulgaria). Acta Zool. Crac. 1984, 27, 149–176. [Google Scholar]
- Popov, V.V. The small mammals (Mammalia: Insectivora, Chiroptera, Lagomorpha, Rodentia) from Cave 16 and the paleoenvironmental changes during the Late Pleistocene. In Temnata Cave: Excavations in Karlukovo Karst Area, Bulgaria; Ginter, B., Kozłowski, J.K., Laville, K., Eds.; Jagiellonian University Press: Cracow, Poland, 2000; Volume 2, pp. 159–240. [Google Scholar]
- Golenishchev, F.N.; Zorenko, T.A.; Petrova, T.V.; Voyta, L.L.; Kryuchkova, L.Y.; Atanasov, N. Evaluation of the “bottleneck” effect in an isolated population of Microtus hartingi (Rodentia, Arvicolinae) from the Eastern Rhodopes (Bulgaria) by methods of integrative analysis. Diversity 2022, 14, 709. [Google Scholar] [CrossRef]
- Guthrie, R.D. Factors regulating the evolution of Microtine tooth complexity. Z. Sägetierk. 1971, 36, 37–54. [Google Scholar]
- Markova, E.A. Assessment of tooth complexity in Arvicolines (Rodentia): A morphotype ranking approach. Zool. Zhurnal 2013, 92, 968–980. (In Russian) [Google Scholar] [CrossRef]
- Markova, E.A.; Sibiryakov, P.A.; Kartavtseva, I.V.; Lapin, A.S.; Morozkina, A.V.; Petukhov, V.A.; Tiunov, M.P.; Starikov, V.P. What can an invasive species tell us about evolution? A study of dental variation in disjunctive populations of Microtus rossiaemeridionalis (Arvicolinae, Rodentia). J. Mamm. Evol. 2019, 26, 267–282. [Google Scholar] [CrossRef]
- Markova, E.; Bobretsov, A.; Borodin, A.; Rakitin, S.; Sibiriakov, P.; Smirnov, N.; Yalkovskaya, L.; Zykov, S. The effects of population bottlenecks on dental phenotype in extant arvicoline rodents: Implications for studies of the Quaternary fossil record. Quat. Sci. Rev. 2020, 228, 106045. [Google Scholar] [CrossRef]
- Markova, E.; Smirnov, N. Phenotypic diversity arising from a limited number of founders: A study of dental variation in laboratory colonies of collared lemmings, Dicrostonyx (Rodentia: Arvicolinae). Biol. J. Linn. Soc. 2018, 125, 777–793. [Google Scholar] [CrossRef]
- Montuire, S.; Royer, A.; Lemanik, A.; Gilg, O.; Sokolova, N.; Sokolov, A.; Desclaux, E.; Nadachowski, A.; Navarro, N. Molar shape differentiation during range expansions of the collared lemming (Dicrostonyx torquatus) related to past climate changes. Quat. Sci. Rev. 2019, 221, 105886. [Google Scholar] [CrossRef]
- Markova, E.; Malygin, V.; Montuire, S.; Nadachowski, A.; Quéré, J.-P.; Ochman, K. Dental variation in sibling species Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia): Between-species comparisons and geography of morphotype dental patterns. J. Mamm. Evol. 2010, 17, 121–139. [Google Scholar] [CrossRef]
- Polly, P.D.; Killick, L.; Ruddy, M. Using left-right asummetry to estimate non-genetic variation in vole teeth (Arvicolinae, Muridae, Rodentia). Palaeont. Electr. 2011, 14, 12. [Google Scholar]
- Maricic, T.; Whitten, M.; Pääbo, S. Multiplexed DNA sequence capture of mitochondrial genomes using PCR products. PLoS ONE 2010, 5, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.R.; Durbin, R.; Subgroup. 1000 genome project data processing the sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample | Lib | Reads | Unique Mapped Reads | Assigned Species | Mean mtDNA Coverage | Fraction of mtDNA Genome Recovered * | Estimated Age (Cal BP) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| M. arvalis | S. anglicus | M. guentheri | Median | 95% HPD | ||||||
| MI2868 | SS | 990,916 | 217 | 1439 | 345 | S. anglicus | 8.4 | 0.83 | 19,290 | [22, 212, 16, 004] |
| DS | 593,827 | 403 | 1133 | 489 | ||||||
| MI2869 | SS | 4,776,656 | 475 | 2440 | 629 | S. anglicus | 17.5 | 0.95 | 20,819 | [23, 474, 16, 858] |
| DS | 2,436,718 | 1048 | 2500 | 1102 | ||||||
| MI2870 | SS | 701,333 | 110 | 608 | 161 | S. anglicus | 5.1 | 0.69 | 19,735 | [22, 575, 15, 943] |
| DS | 808,330 | 316 | 874 | 352 | ||||||
| MI2871 | SS | 86,906 | 238 | 75 | 123 | M. arvalis | 2.9 | 0.46 | n.a. | n.a. |
| DS | 280,844 | 521 | 316 | 431 | ||||||
| MI2872 | SS | 3,510,503 | 8339 | 2837 | 4884 | M. arvalis | 27.8 | 0.96 | 23,410 | [27, 369, 19, 290] |
| DS | 113,370 | 862 | 416 | 639 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadachowski, A.; Lemanik, A.; Fontana, L.; Popović, D.; Golubiński, M.; Bujalska, B.; Baca, M. Ancient DNA Contradicts the Presence of Social Voles (Genus Microtus, Subgenus Sumeriomys) in the Late Pleistocene of Western Europe. Diversity 2023, 15, 538. https://doi.org/10.3390/d15040538
Nadachowski A, Lemanik A, Fontana L, Popović D, Golubiński M, Bujalska B, Baca M. Ancient DNA Contradicts the Presence of Social Voles (Genus Microtus, Subgenus Sumeriomys) in the Late Pleistocene of Western Europe. Diversity. 2023; 15(4):538. https://doi.org/10.3390/d15040538
Chicago/Turabian StyleNadachowski, Adam, Anna Lemanik, Laure Fontana, Danijela Popović, Michał Golubiński, Barbara Bujalska, and Mateusz Baca. 2023. "Ancient DNA Contradicts the Presence of Social Voles (Genus Microtus, Subgenus Sumeriomys) in the Late Pleistocene of Western Europe" Diversity 15, no. 4: 538. https://doi.org/10.3390/d15040538
APA StyleNadachowski, A., Lemanik, A., Fontana, L., Popović, D., Golubiński, M., Bujalska, B., & Baca, M. (2023). Ancient DNA Contradicts the Presence of Social Voles (Genus Microtus, Subgenus Sumeriomys) in the Late Pleistocene of Western Europe. Diversity, 15(4), 538. https://doi.org/10.3390/d15040538