

Initial Population Analysis and Mycorrhizal Fungi of the Leafless Epiphytic Orchid, Campylocentrum pachyrrhizum, in Florida

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Survey Methods
2.3. Isolation and Identification of Mycorrhizal Fungi
2.4. Molecular Confirmation
3. Results
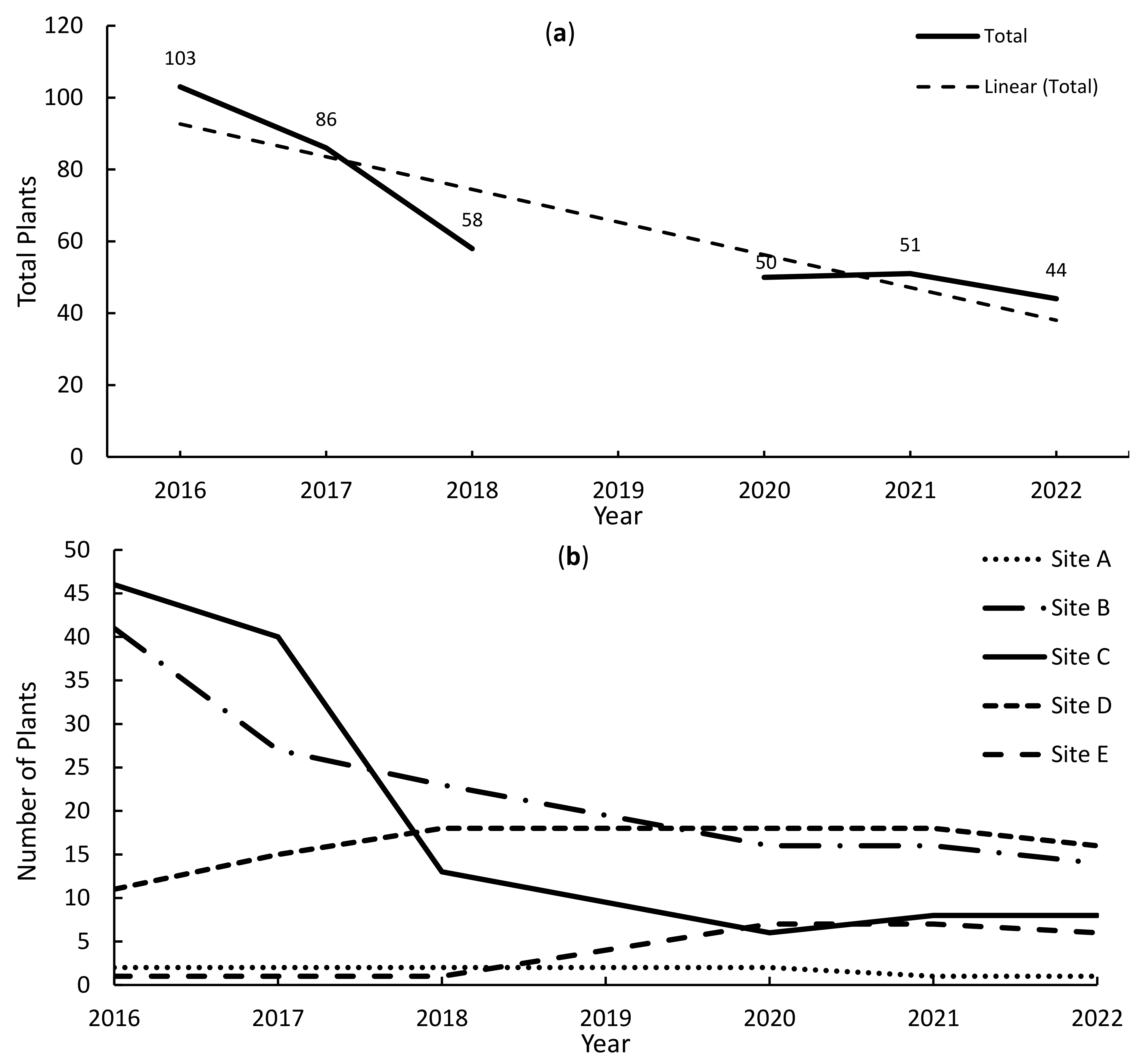
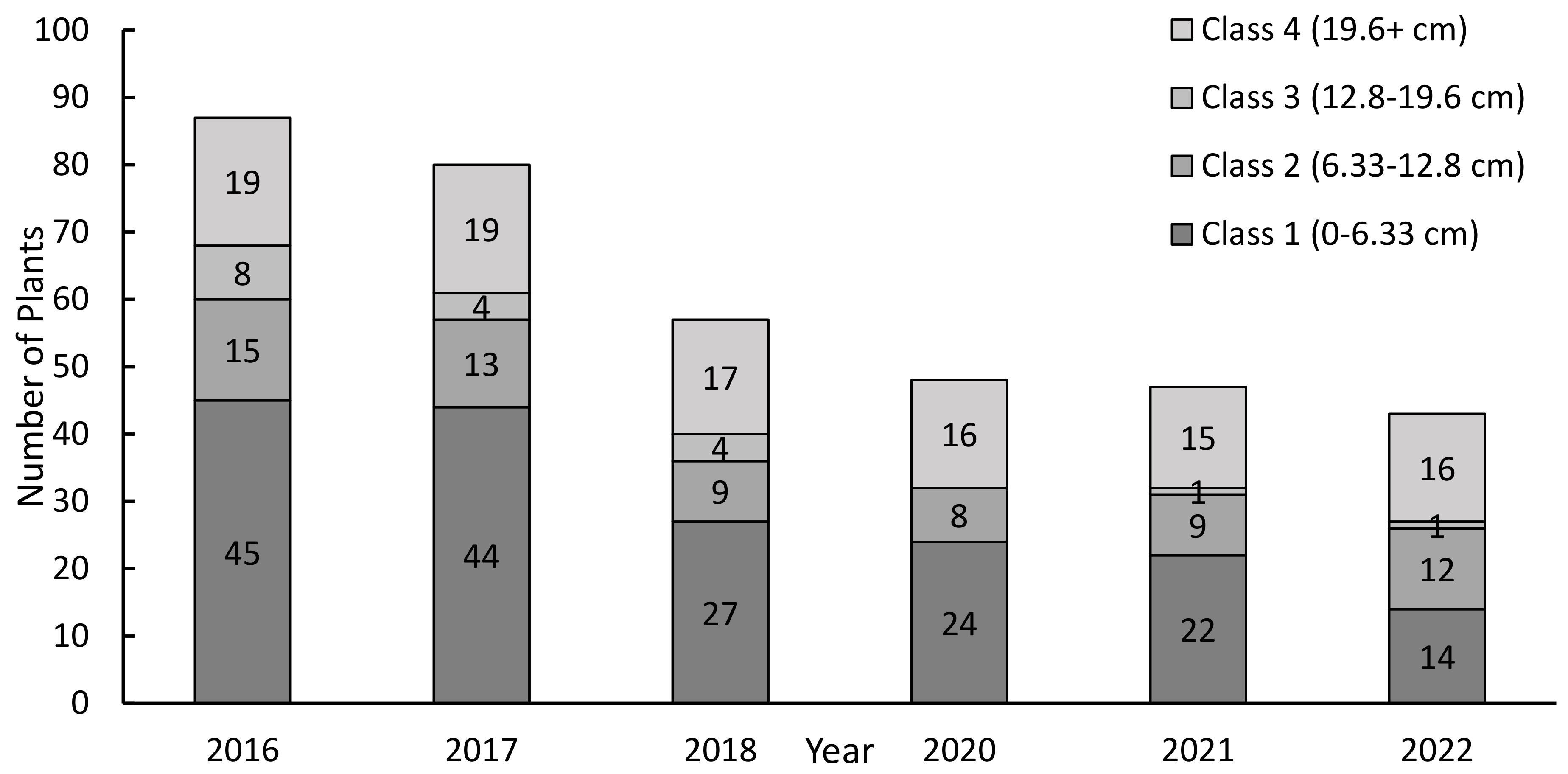
3.1. Orchid Observations
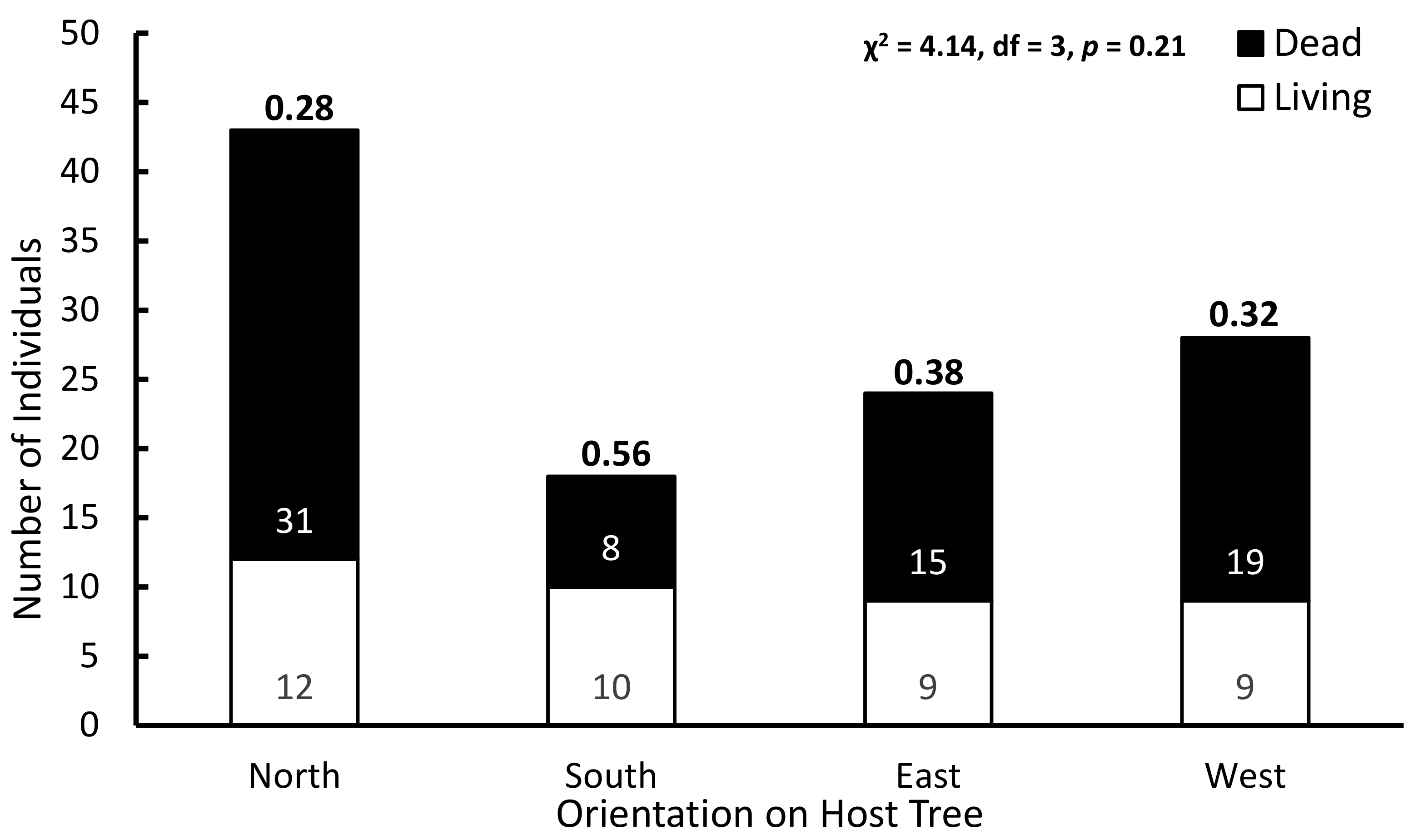
3.2. Host Tree Numbers and Interactions
3.3. Mycorrhizal Fungi
4. Discussion
4.1. Hosting a Funeral: Host Trees, Ribbon Orchid Numbers, and Mortality
4.2. Mycorrhizal Fungi
4.3. Seedling Observations—Living Dangerously?
4.4. Recommendations for Immediate Conservation
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- North American Orchid Conservation Center. Available online: https://northamericanorchidcenter.org/ (accessed on 20 October 2022).
- Brown, P.M.; Folsom, S. Wild Orchids of Florida: With References to the Atlantic and Gulf Coastal Plains; University Press of Florida: Gainesville, FL, USA, 2002. [Google Scholar]
- Luer, C.A. The Native Orchids of Florida; New York Botanical Garden: New York, NY, USA, 1972. [Google Scholar]
- Swarts, N.D.; Dixon, K.W. Terrestrial Orchid Conservation in the Age of Extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Mújica, E.B.; Herdman, A.R.; Danaher, M.W.; González, E.H.; Zettler, L.W. Projected status of the Ghost Orchid (Dendrophylax lindenii) in Florida during the next decade based on temporal dynamic studies spanning six years. Plants 2021, 10, 1579. [Google Scholar] [CrossRef]
- Stewart, S.L.; Richardson, L. Orchids of the Florida Panther National Wildlife Refuge. North Am. Nativ. Orchid. J. Collier 2008, 14, 70–104. [Google Scholar]
- Florida Administrative Code & Florida Administrative Register; 5B-40.0055: Regulated Plant Index—Florida Administrative Rules, Law, Code. Available online: https://www.flrules.org/gateway/ruleno.asp?ID=5B-40.0055 (accessed on 25 October 2022).
- Mújica, E.B.; Mably, J.J.; Skarha, S.M.; Corey, L.L.; Richardson, L.W.; Danaher, M.W.; González, E.H.; Zettler, L.W. A comparision of Ghost Orchid (Dendrophylax lindenii) habitats in Florida and Cuba, with particular reference to seedling recruitment and mycorrhizal fungi. Bot. J. Linn. Soc. 2018, 186, 572–586. [Google Scholar] [CrossRef]
- Raventós, J.; González, E.; Mújica, E.; Bonet, A. Transient population dynamics of two epiphytic orchid species after hurricane Ivan: Implications for management. Biotropica 2015, 47, 441–448. [Google Scholar] [CrossRef]
- Zettler, L.W.; Corey, L.L. Orchid Mycorrhizal Fungi: Isolation and Identification Techniques; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar] [CrossRef]
- Clements, M.A.; Ellyard, R.K. The symbiotic germination of Australian terrestrial orchids [Pterostylis, Diuris, Thelymitra Inocultates with Mycorrhizal Fungi Tulasnella and Ceratobandium]. Am. Orchid Soc. Bull. 1979, 48, 810–815. [Google Scholar]
- Currah, R.S.; Sigler, L.; Hambleton, S. New records and new taxa of fungi from the mycorrhizae of terrestrial orchids of Alberta. Can. J. Bot. 1987, 65, 2473–2482. [Google Scholar] [CrossRef]
- Currah, R.; Smreciu, E.A.; Hambleton, S. Mycorrhizae and mycorrhizal fungi of boreal species of Platanthera and Coeloglossum (Orchidaceae). Can. J. Bot. 1990, 68, 1171–1181. [Google Scholar] [CrossRef]
- Richardson, K.A. Basidiomycetous endophytes from the roots of neotropical epiphytic orchidaceae. Lindleyana 1993, 8, 127–137. [Google Scholar]
- Zettler, L.W.; Corey, L.L.; Jacks, A.L.; Gruender, L.T.; Lopez, A.M. Tulasnella irregularis (Basidiomycota: Tulasnellaceae) from roots of Encyclia tampensis in south Florida, and confirmation of its significance through symbiotic seed germination. Lankesteriana 2013, 13, 119–128. [Google Scholar] [CrossRef]
- Yokoya, K.; Zettler, L.W.; Kendon, J.P.; Bidartondo, M.I.; Stice, A.L.; Skarha, S.; Corey, L.L.; Knight, A.C.; Sarasan, V. Preliminary findings on identification of mycorrhizal fungi from diverse orchids in the central highlands of Madagascar. Mycorrhiza 2015, 25, 611–625. [Google Scholar] [CrossRef]
- Thixton, H.L.; Esselman, E.J.; Corey, L.L.; Zettler, L.W. Further evidence of Ceratobasidium D. P. Rogers (Basidiomycota) serving as the ubiquitous fungal associate of Platanthera leucophaea (Orchidaceae) in the north american tallgrass prairie. Bot. Stud. 2020, 61, 12. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, D.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Taylor, D.L.; McCormick, M.K. Internal transcribed spacer primers and sequences for improved characterization of basidiomycetous orchid mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Florida Panther National Wildlife Refuge Eastern Weather Station, Raw Land Cover: 2001 USGS DATA COURTESY OF: Bureau of Land Management and Dept of Interior Fish & Wildlife Service. Available online: https://mesowest.utah.edu/cgi-bin/droman/meso_base_dyn.cgi?stn=PSTF1&time=GMT (accessed on 1 November 2022).
- Haaland, D.; Estenoz, S.; Williams, L.; Williams, M. Petition to List the Ghost Orchid (Dendrophylax lindenii) as Threatened or Endangered under the Endangered Species Act and To Designate Critical Habitat. 2022. Available online: https://regionalconservation.org/ircs/pdf/2022_01_24_ghost_orchid_ESA_petition.pdf (accessed on 1 November 2022).
- Borrero, H.; Oviedo-Prieto, R.; Alvarez, J.C.; Ticktin, T.; Cisneros, M.; Liu, H. Populations of a tropical epiphytic orchid are destabilized in its peripheral range by hurricane and an exotic herbivore. Ecosphere 2023, 14, e4355. [Google Scholar] [CrossRef]
- Mújica, E.; Raventós, J.; González, E.; Bonet, A. Long-term hurricane effects on populations of two epiphytic orchid species from Guanahacabibes Peninsula, Cuba. Lankesteriana 2013, 13, 47–55. [Google Scholar] [CrossRef]
- Cabral, P.R.; Pansarin, E.R. Biologia reprodutiva de Campylocentrum micranthum (Orchidaceae, Angraecinae). Rodriguesia 2016, 67, 379–386. [Google Scholar] [CrossRef]
- Carlsward, B.S.; Whitten, W.M.; Williams, N.H.; Bytebier, B. Molecular phylogenetics of Vandeae (Orchidaceae) and the evolution of leaflessness. Am. J. Bot. 2006, 93, 770–786. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.L.; Kane, M.E. Symbiotic seed germination of Habenaria macroceratitis (Orchidaceae), a rare Florida terrestrial orchid. Plant Cell. Tissue Organ Cult. 2006, 86, 159–167. [Google Scholar] [CrossRef]
- Yukawa, T.; Ogura-Tsujita, Y.; Shefferson, R.P.; Yokoyama, J. Mycorrhizal diversity in Apostasia (Orchidaceae) indicates the origin and evolution of orchid mycorrhiza. Am. J. Bot. 2009, 96, 1997–2009. [Google Scholar] [CrossRef] [PubMed]
- Chomicki, G.; Bidel, L.P.R.; Jay-Allemand, C. Exodermis structure controls fungal invasion in the leafless epiphytic orchid Dendrophylax lindenii (Lindl.) Benth. Ex Rolfe. Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 88–94. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial orchids: From seed to mycotrophic plant. In Terrestrial Orchids: From Seed to Mycotrophic Plant; American Society of Plant Taxonomists: Laramie, WY, USA, 1995; Volume 21. [Google Scholar] [CrossRef]
- Otero, J.T.; Ackerman, J.D.; Bayman, P. Diversity and host specificity of endophytic rhizoctonia-like fungi from tropical orchids. Am. J. Bot. 2002, 89, 1852–1858. [Google Scholar] [CrossRef] [PubMed]
- Hoang, N.H.; Kane, M.E.; Radcliffe, E.N.; Zettler, L.W.; Richardson, L.W. Comparative seed germination and seedling development of the Ghost Orchid, Dendrophylax lindenii (Orchidaceae), and molecular identification of Its mycorrhizal fungus from south florida. Ann. Bot. 2017, 119, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Zhang, W.; Feng, J.Q.; Zhang, S.B. Leafless epiphytic orchids share Ceratobasidiaceae mycorrhizal fungi. Mycorrhiza 2021, 31, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Unruh, S.A.; Pires, J.C.; Zettler, L.W.; Erba, L.; Grigoriev, I.; Barry, K.; Daum, C.; Lipzen, A.; Stajich, J.E. Shallow genome sequencing for phylogenomics of mycorrhizal fungi from endangered ochids. bioRxiv 2019. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop Ash | Pond Apple | |
|---|---|---|
| Hosts Observed | 17 | 4 |
| Ind Obs Alive | 36 | 8 |
| Ind Obs Dead | 70 | 4 |
| Ind Obs totals | 106 | 12 |
| % survival of orchids | 34 | 66.7 |
| % of orchid pop | 89.8 | 10.2 |
| Corrugated Bark | Semi-Corrugated Bark | Smooth Bark | |
|---|---|---|---|
| Obs Alive | 11 | 31 | 2 |
| Obs Dead | 4 | 60 | 10 |
| Obs totals | 15 | 91 | 12 |
| % survival | 73.3 | 34.0 | 16.7 |
| % of pop | 12.7 | 77.1 | 10.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herdman, A.R.; Mújica, E.B.; Danaher, M.W.; Zettler, L.W.; Schulz, K.; Esselman, E. Initial Population Analysis and Mycorrhizal Fungi of the Leafless Epiphytic Orchid, Campylocentrum pachyrrhizum, in Florida. Diversity 2023, 15, 576. https://doi.org/10.3390/d15040576
Herdman AR, Mújica EB, Danaher MW, Zettler LW, Schulz K, Esselman E. Initial Population Analysis and Mycorrhizal Fungi of the Leafless Epiphytic Orchid, Campylocentrum pachyrrhizum, in Florida. Diversity. 2023; 15(4):576. https://doi.org/10.3390/d15040576
Chicago/Turabian StyleHerdman, Adam R., Ernesto B. Mújica, Mark W. Danaher, Lawrence W. Zettler, Kurt Schulz, and Elizabeth Esselman. 2023. "Initial Population Analysis and Mycorrhizal Fungi of the Leafless Epiphytic Orchid, Campylocentrum pachyrrhizum, in Florida" Diversity 15, no. 4: 576. https://doi.org/10.3390/d15040576
APA StyleHerdman, A. R., Mújica, E. B., Danaher, M. W., Zettler, L. W., Schulz, K., & Esselman, E. (2023). Initial Population Analysis and Mycorrhizal Fungi of the Leafless Epiphytic Orchid, Campylocentrum pachyrrhizum, in Florida. Diversity, 15(4), 576. https://doi.org/10.3390/d15040576