


Characteristics of the Stone Crayfish Population along a Disturbance Gradient—A Case Study of the Kustošak Stream, Croatia

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Data Collection
2.2. Data Analyses
2.2.1. Population Size
2.2.2. Size Class Frequency
2.2.3. Fulton’s Condition Factor
2.2.4. Analysis of Physicochemical Parameters of the Water
2.3. Statistical Analyses
2.4. Genetic Diversity and Structure
2.4.1. Mitochondrial DNA Analyses
2.4.2. Microsatellite DNA Analyses
3. Results
3.1. Descriptive Statistics
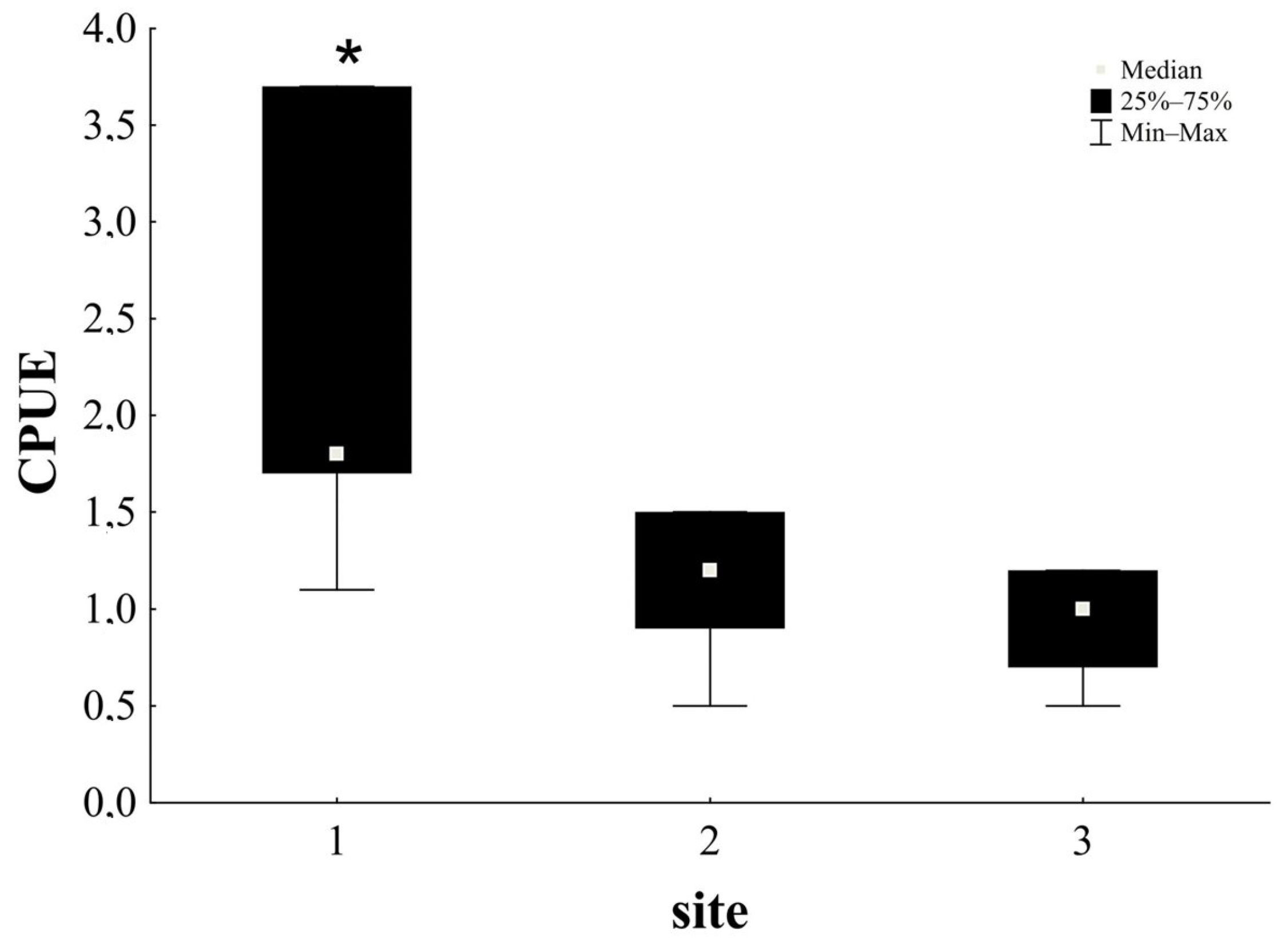
3.2. Estimation of Total and Relative Population Size
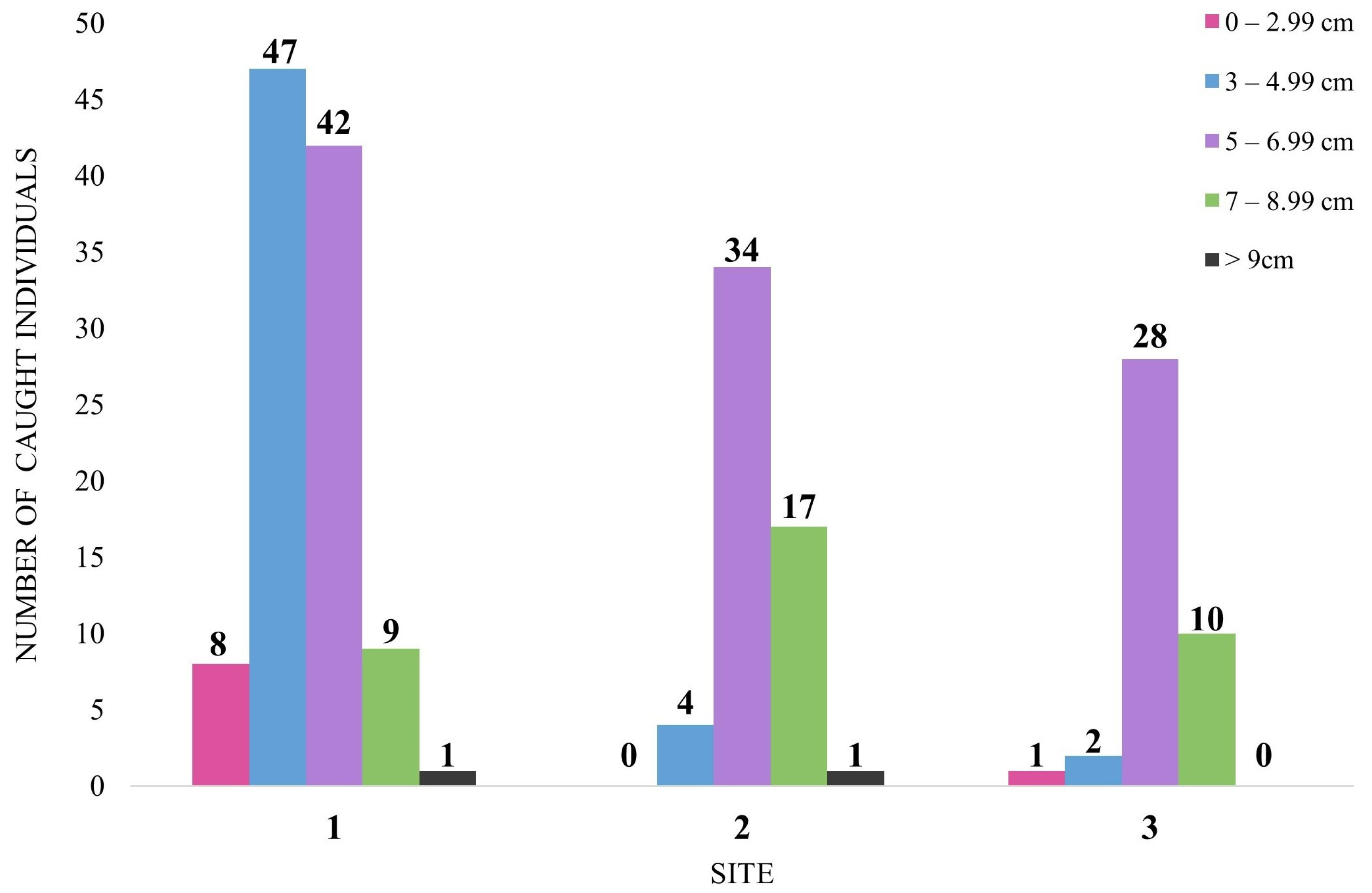
3.3. Size Structure and Crayfish Condition
3.4. Effect of Physicochemical Parameters of Water Quality on CPUE
3.5. Genetic Diversity and Structure
3.5.1. Mitochondrial DNA Analyses
3.5.2. Microsatellite DNA Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kouba, A.; Petrusek, A.; Kozák, P. Continental-wide distribution of crayfish species in Europe: Update and maps. Knowl. Manag. Aquat. Ecosyst. 2014, 413, 5. [Google Scholar] [CrossRef]
- Souty-Grosset, C.; Holdich, D.; Noel, P.; Reynolds, J.D. (Eds.) Atlas of Crayfish in Europe; Muséum National D’histoire Naturelle: Paris, France, 2006. [Google Scholar]
- Maguire, I.; Klobučar, G.; Žganec, K.; Jelić, M.; Lucić, A.; Hudina, S. Recent changes in distribution pattern of freshwater crayfish in Croatia—Threats and perspectives. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 2. [Google Scholar] [CrossRef]
- Jussila, J.; Vrezec, A.; Jaklič, T.; Kukkonen, H.; Makkonen, J.; Kokko, H. Aphanomyces astaci isolate from latently infected stone crayfish (Austropotamobius torrentium) population is virulent. J. Invertebr. Pathol. 2017, 149, 15–20. [Google Scholar] [CrossRef]
- Kozák, P.; Ďuriš, Z.; Petrusek, A.; Buřič, M.; Horká, I.; Kouba, A.; Kozubíková-Balcarová, E.; Policar, T. Crayfish Biology and Culture; University of South Bohemia in České Budějovice, Faculty of Fisheries and Protection of Waters: České Budějovice, Czechia, 2015. [Google Scholar]
- Marn, N.; Hudina, S.; Haberle, I.; Dobrović, A.; Klanjšček, T. Physiological performance of native and invasive crayfish species in a changing environment: Insights from Dynamic Energy Budget models. Conserv. Physiol. 2022, 10, coac031. [Google Scholar] [CrossRef] [PubMed]
- Article 17 Web Tool. Available online: https://www.eionet.europa.eu/article17/species/summary/?period=5&group=Arthropods&subject=Austropotamobius+torrentium®ion= (accessed on 5 December 2022).
- Convention of 19 September 1979 on the Conservation of European Wildlife and Natural Habitats; Council of Europe: Strasbourg, France, 1979.
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Maguire, I.; Jelić, M.; Klobučar, G. Update on the distribution of freshwater crayfish in Croatia. Knowl. Manag. Aquat. Ecosyst. 2011, 401, 31. [Google Scholar] [CrossRef]
- Füreder, L.; Gherardi, F.; Souty-Grosset, C. Austropotamobius torrentium (errata version published in 2017). IUCN Red List. Threat. Species 2010, 4, T2431A121724677. [Google Scholar]
- Klobučar, G.I.V.; Podnar, M.; Jelić, M.; Franjević, D.; Faller, M.; Štambuk, A.; Gottstein, S.; Simić, V.; Maguire, I. Role of the Ddinaric Karst (western Balkans) in shaping the phylogeographic structure of the threatened crayfish Austropotamobius torrentium. Freshw. Biol. 2013, 58, 1089–1105. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Pérez-Moreno, J.L.; Panaiotu, C.; Drăguț, L.; Schrimpf, A.; Popovici, I.; Zaharia, C.; Weiperth, A.; Gál, B.; Schubart, C.D.; et al. A journey on plate tectonics sheds light on European crayfish phylogeography. Ecol. Evol. 2019, 9, 1957–1971. [Google Scholar] [CrossRef]
- Lovrenčić, L.; Bonassin, L.; Boštjančić, L.L.; Podnar, M.; Jelić, M.; Klobučar, G.; Jaklič, M.; Slavevska-Stamenković, V.; Hinić, J.; Maguire, I. New insights into the genetic diversity of the stone crayfish: Taxonomic and conservation implications. BMC Evol. Biol. 2020, 20, 146. [Google Scholar] [CrossRef]
- Lucić, A.; Žganec, K. Istraživanje Populacija Potočnih Rakova (Austropotamobius torrentium) u Parku Prirode Medvednica Konačno Izvješće; Udruga za biološka istraživanja—BIOM: Zagreb, Croatia, 2010. [Google Scholar]
- Lucić, A.; Žganec, K.; Hudina, S. Monitoring Potočnih Rakova (Austropotamobius torrentium) na Području PP Medvednica Konačno Izvješće; Udruga za biološka istraživanja—BIOM: Zagreb, Croatia, 2012. [Google Scholar]
- Lucić, A.; Petljak, K.V.; Žganec, K. Monitoring Potočnih Rakova (Austropotamobius torrentium) na Području PP Medvednica Konačni Izvještaj; Udruga za biološka istraživanja—BIOM: Zagreb, Croatia, 2015. [Google Scholar]
- Maguire, I. Nacionalni Programi za Praćenje Stanja Očuvanosti Vrsta i Staništa u Hrvatskoj. Potočni rak ili rak Kamenjar Austropotamobius torrentium (Schrank, 1803); Državni Zavod za Zaštitu Prirode: Zagreb, Croatia, 2014; Available online: https://www.haop.hr/sites/default/files/uploads/dokumenti/03_prirodne/monitoring_prog/FINAL%20Austropotamobius_torrentium%2008-17.pdf (accessed on 18 November 2022).
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Allendorf, F.W.; Luikart, G.; Aitken, S.N. Conservation and the Genetics of Populations; Wiley-Blackwell: Oxford, UK, 2013. [Google Scholar]
- Schnabel, Z.E. The Estimation of total fish population of a lake. Am. Math. Mon. 1938, 45, 348–352. [Google Scholar] [CrossRef]
- Schumacher, F.X.; Eschmeyer, R.W. The estimate of fish population in lakes or ponds. Tenn. Acad. Sci. 1943, 18, 228–249. [Google Scholar]
- Maguire, I.; Klobučar, G. Size structure, maturity size, growth and condition index of stone crayfish (Austropotamobius torrentium) in North-West Croatia. Knowl. Manag. Aquat. Ecosyst. 2011, 401, 12. [Google Scholar] [CrossRef]
- Fulton, T.W. Twenty-Second Annual Report. The Rate of Growth of Fishes; Fisheries Board of Scotland: Edinburgh, UK, 1902; Volume 3, pp. 326–446. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.R.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial Cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 224–229. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Villesen, P. FaBox: An online toolbox for Fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Lovrenčić, L.; Temunović, M.; Bonassin, L.; Grandjean, F.; Austin, C.M.; Maguire, I. Climate change threatens unique genetic diversity within the Balkan biodiversity hotspot—The case of the endangered stone crayfish. Glob. Ecol. Conserv. 2022, 39, e02301. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT Version 2.9.4: A Program to Estimate and Test Population Genetics Parameters. Updated from Goudet (1995). Available online: https://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 18 October 2022).
- Piry, S.; Luikart, G.; Cornuet, J.-M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE—A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; Von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Jussila, J.; Edsman, L.; Maguire, I.; Diéguez-Uribeondo, J.; Theissinger, K. Money kills native ecosystems: European crayfish as an example. Front. Ecol. Evol. 2021, 9, 648495. [Google Scholar] [CrossRef]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef]
- Bohl, E. An isolated population of the white-clawed crayfish (Austropotamobius pallipes) in the principality of Liechtenstein. Bull. Français De La Pêche Et De La Piscic. 1997, 347, 701–712. [Google Scholar] [CrossRef]
- Cooper, A.R.; Infante, D.M.; Daniel, W.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Assessment of dam effects on streams and fish assemblages of the conterminous USA. Sci. Total Environ. 2017, 586, 879–889. [Google Scholar] [CrossRef]
- Nevers, R.J. A State-of-The-Unionids Address. In Conservation and Management of Freshwater Mussels. Proceedings of a UMRCC Symposium; Upper Mississippi River Conservation Committee: Rock Island, TN, USA, 1993; pp. 1–10. [Google Scholar]
- Page, L.M.; Pyron, M.; Cummings, K.S. Impacts of fragmentation on midwestern aquatic organisms. In Conservation in Highly Fragmented Landscapes; Springer: Boston, MA, USA, 1997; pp. 189–212. [Google Scholar]
- Light, T.; Erman, D.C.; Myrick, C.; Clarke, J. Decline of the shasta crayfish (Pacifastacus fortis Faxon) of Northeastern California. Conserv. Biol. 1995, 9, 1567–1577. [Google Scholar] [CrossRef]
- Maguire, I.; Gottstein-Matočec, S. The distribution pattern of freshwater crayfish in Croatia. Crustaceana 2004, 77, 25–48. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. Quantifying threats to imperiled species in the United States. BioScience 1998, 48, 607–615. [Google Scholar] [CrossRef]
- Lodge, D.M.; Hill, A.M. Factors governing species composition, population size, and productivity of cool-water crayfishes. Nord. J. Freshw. Res. 1994, 69, 111–136. [Google Scholar]
- Smith, G.R.T.; Learner, M.A.; Slater, F.M.; Foster, J. Habitat features important for the conservation of the native crayfish Austropotamobius pallipes in Britain. Biol. Conserv. 1996, 75, 239–246. [Google Scholar] [CrossRef]
- Huber, M.G.J.; Sschubart, C.D. Distribution and reproductive biology of Austropotamobius torrentium in Bavaria and documentation of a contact zone with the alien crayfish Pacifastacus leniusculus. Bull. Français De La Pêche Et De La Piscic. 2005, 376–377, 759–776. [Google Scholar] [CrossRef]
- Streissl, F.; Hödl, W. Growth, Morphometrics, size at maturity, sexual dimorphism and condition index of Austropotamobius torrentium Schrank. Hydrobiologia 2002, 477, 201–208. [Google Scholar] [CrossRef]
- Pöckl, M.; Streissl, F. Austropotamobius torrentium as an Indicator for Habitat Quality in Running Waters? Bull. Français De La Pêche Et De La Piscic. 2005, 376–377, 743–758. [Google Scholar] [CrossRef]
- Kadlecová, K.; Bílý, M.; Maciak, M. Movement Patterns of the Co-Occurring Species Astacus Astacus (Noble Crayfish) and Austropotamobius Torrentium (Stone Crayfish). Fundam. Appl. Limnol. Arch. Für Hydrobiol. 2012, 180, 351–360. [Google Scholar] [CrossRef]
- Anastácio, P.M.; Marques, J.C. Crayfish (Procambarus clarkii) condition throughout the year in the lower Mondego River Valley, Portugal. Crustaceana 1998, 71, 593–602. [Google Scholar] [CrossRef]
- Bohl, E. Comparative studies on crayfish brooks in Bavaria (Astacus astacus L., Austropotamobius torrentium Schr.). Freshw. Crayfish 1987, 7, 287–294. [Google Scholar]
- Demers, A.; Soutty-Grosset, C.; Trouilhé, M.C.; Füreder, L.; Renai, B.; Gherardi, F. Tolerance of three European native species of cayfish to hypoxia. Hydrobiologia 2006, 560, 425–432. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Pacioglu, O.; Hamchevici, C. The assessment of the habitat and water quality requirements of the stone crayfish (Austropotamobius torrentium) and noble crayfish (Astacus astacus) species in the rivers from the Anina Mountains (SW Romania). Knowl. Manag. Aquat. Ecosyst. 2011, 401, 03. [Google Scholar] [CrossRef]
- Holdich, D.M. Biology of Freshwater Crayfish; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- Carvalho, C.S.; Lanes, É.C.M.; Silva, A.R.; Caldeira, C.F.; Carvalho-Filho, N.; Gastauer, M.; Imperatriz-Fonseca, V.L.; Júnior, W.N.; Oliveira, G.; Siqueira, J.O.; et al. Habitat loss does not always entail negative genetic consequences. Front. Genet. 2019, 10, 1011. [Google Scholar] [CrossRef]
- Gladstone, N.S.; Garrison, N.L.; Lane, T.; Johnson, P.D.; Garner, J.; Whelan, N.V. Population genomics reveal low differentiation and complex demographic histories in a highly fragmented and endangered freshwater mussel. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 1235–1248. [Google Scholar] [CrossRef]
- Hurt, C.; Hildreth, P.; Williams, C. A Genomic perspective on the conservation status of the endangered nashville crayfish (Faxonius shoupi). Conserv. Genet. 2022, 23, 589–604. [Google Scholar] [CrossRef]
- Markert, J.A.; Champlin, D.M.; Gutjahr-Gobell, R.; Grear, J.S.; Kuhn, A.; McGreevy, T.J.; Roth, A.; Bagley, M.J.; Nacci, D.E. Population genetic diversity and fitness in multiple environments. BMC Evol. Biol. 2010, 10, 205. [Google Scholar] [CrossRef]
- Wcenter, L.I.; Tregenza, T.; Hosken, D.J. Inbreeding, inbreeding depression and extinction. Conserv. Genet. 2007, 9, 833–843. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.-P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- Leigh, D.M.; Hendry, A.P.; Vázquez-Domínguez, E.; Friesen, V.L. Estimated six per cent loss of genetic variation in wild populations since the industrial revolution. Evol. Appl. 2019, 12, 1505–1512. [Google Scholar] [CrossRef]
- Lino, A.; Fonseca, C.; Rojas, D.; Fischer, E.; Ramos Pereira, M.J. A meta-analysis of the effects of habitat loss and fragmentation on genetic diversity in mammals. Mamm. Biol. 2019, 94, 69–76. [Google Scholar] [CrossRef]
- Barnett, Z.C.; Adams, S.B.; Ochs, C.A.; Garrick, R.C. Crayfish populations genetically fragmented in streams impounded for 36–104 years. Freshw. Biol. 2020, 65, 768–785. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Abbreviation | Measuring Unit | |
|---|---|---|---|
| 1 | Dissolved oxygen | O2 | mg/L |
| 2 | Water temperature | w. temp. | °C |
| 3 | Water pH | pH | / |
| 4 | Oxidation reduction potential | ORP | mV |
| 5 | Water turbidity | turbidity | NTU |
| 6 | Chlorides | Cl− | mg/L |
| 7 | Nitrates | NO3− | mg/L |
| 8 | Sulphates | SO42− | mg/L |
| 9 | Sodium ions | Na+ | mg/L |
| 10 | Potassium ions | K+ | mg/L |
| 11 | Magnesium ions | Mg2+ | mg/L |
| 12 | Calcium ions | Ca2+ | mg/L |
| 13 | Ammonium | NH4+ | mg/L |
| 14 | Chemical oxygen demand | COD | mg/L |
| 15 | Nitrate nitrogen | NO3−N | mg/L |
| 16 | Nitrites | NO2− | mg/L |
| 17 | Total nitrogen | TN | mg/L |
| 18 | Total phosphorus | TP | mg/L |
| Site | Sex | TL [mm] | w [g] | FCF [g/mm3] | CPUE |
|---|---|---|---|---|---|
| 1 | male | 49.97585 ± 13.76868 | 6.57317 ± 6.50822 | 0.00435 ± 0.00086 | 2.19756 ± 0.96475 |
| 1 | female | 54.22271 ± 12.92085 | 6.87879 ± 4.47218 | 0.00390 ± 0.00074 | 2.51212 ± 0.99591 |
| 2 | male | 62.50467 ± 9.82911 | 10.5000 ± 5.34499 | 0.00402 ± 0.00052 | 1.13000 ± 0.30529 |
| 2 | female | 65.90153 ± 12.56042 | 11.30769 ± 5.50468 | 0.00417 ± 0.00318 | 1.10370 ± 0.31191 |
| 3 | male | 59.93421 ± 11.54536 | 10.23684 ± 5.46038 | 0.00556 ± 0.00659 | 0.94210 ± 0.24339 |
| 3 | female | 65.24272 ± 8.99621 | 10.54545 ± 4.51692 | 0.00358 ± 0.00046 | 0.95000 ± 0.25023 |
| Schumacher–Eschmeyer Method | Schnabel Method | |||
|---|---|---|---|---|
| site | estimation | estimation | lower confidence limit | upper confidence limit |
| 1 | 328 | 346 | 251 | 555 |
| 2 | 161 | 98 | 77 | 139 |
| 3 | 87 | 60 | 46 | 86 |
| Site | N | P | NA | AR | APR | uHE | HO | FIS | HW |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 15 | 1 | 4.00 | 3.73 | 5 | 0.597 | 0.524 | 0.126 | ns |
| 2 | 13 | 1 | 3.71 | 3.56 | 3 | 0.496 | 0.465 | 0.065 | ns |
| 3 | 12 | 1 | 3.86 | 3.73 | 5 | 0.569 | 0.502 | 0.122 | ns |
| Site | Wilcoxon Sign-Rank Test 1-Tail | Wilcoxon Sign-Rank Test 2t | ||||
|---|---|---|---|---|---|---|
| IAM | TPM | SMM | IAM | TPM | SMM | |
| 1 | 0.03 | 0.15 | 0.41 | 0.05 | 0.30 | 0.81 |
| 2 | 0.34 | 0.71 | 0.95 | 0.69 | 0.69 | 0.30 |
| 3 | 0.04 | 0.34 | 0.66 | 0.08 | 0.69 | 0.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarandek, A.; Lovrenčić, L.; Židak, L.; Topić, M.; Grbin, D.; Gregov, M.; Ćurko, J.; Hudina, S.; Maguire, I. Characteristics of the Stone Crayfish Population along a Disturbance Gradient—A Case Study of the Kustošak Stream, Croatia. Diversity 2023, 15, 591. https://doi.org/10.3390/d15050591
Tarandek A, Lovrenčić L, Židak L, Topić M, Grbin D, Gregov M, Ćurko J, Hudina S, Maguire I. Characteristics of the Stone Crayfish Population along a Disturbance Gradient—A Case Study of the Kustošak Stream, Croatia. Diversity. 2023; 15(5):591. https://doi.org/10.3390/d15050591
Chicago/Turabian StyleTarandek, Anita, Leona Lovrenčić, Lana Židak, Martina Topić, Dorotea Grbin, Marija Gregov, Josip Ćurko, Sandra Hudina, and Ivana Maguire. 2023. "Characteristics of the Stone Crayfish Population along a Disturbance Gradient—A Case Study of the Kustošak Stream, Croatia" Diversity 15, no. 5: 591. https://doi.org/10.3390/d15050591
APA StyleTarandek, A., Lovrenčić, L., Židak, L., Topić, M., Grbin, D., Gregov, M., Ćurko, J., Hudina, S., & Maguire, I. (2023). Characteristics of the Stone Crayfish Population along a Disturbance Gradient—A Case Study of the Kustošak Stream, Croatia. Diversity, 15(5), 591. https://doi.org/10.3390/d15050591