
Heterochlamydomonas uralensis sp. nov. (Chlorophyta, Chlamydomonadaceae), New Species Described from the Mountain Tundra Community in the Subpolar Urals (Russia)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites, Sample Collection and Culturing Conditions
2.2. Light and Transmission Electron Microscopy
2.3. DNA Extraction, Amplification and Sequencing
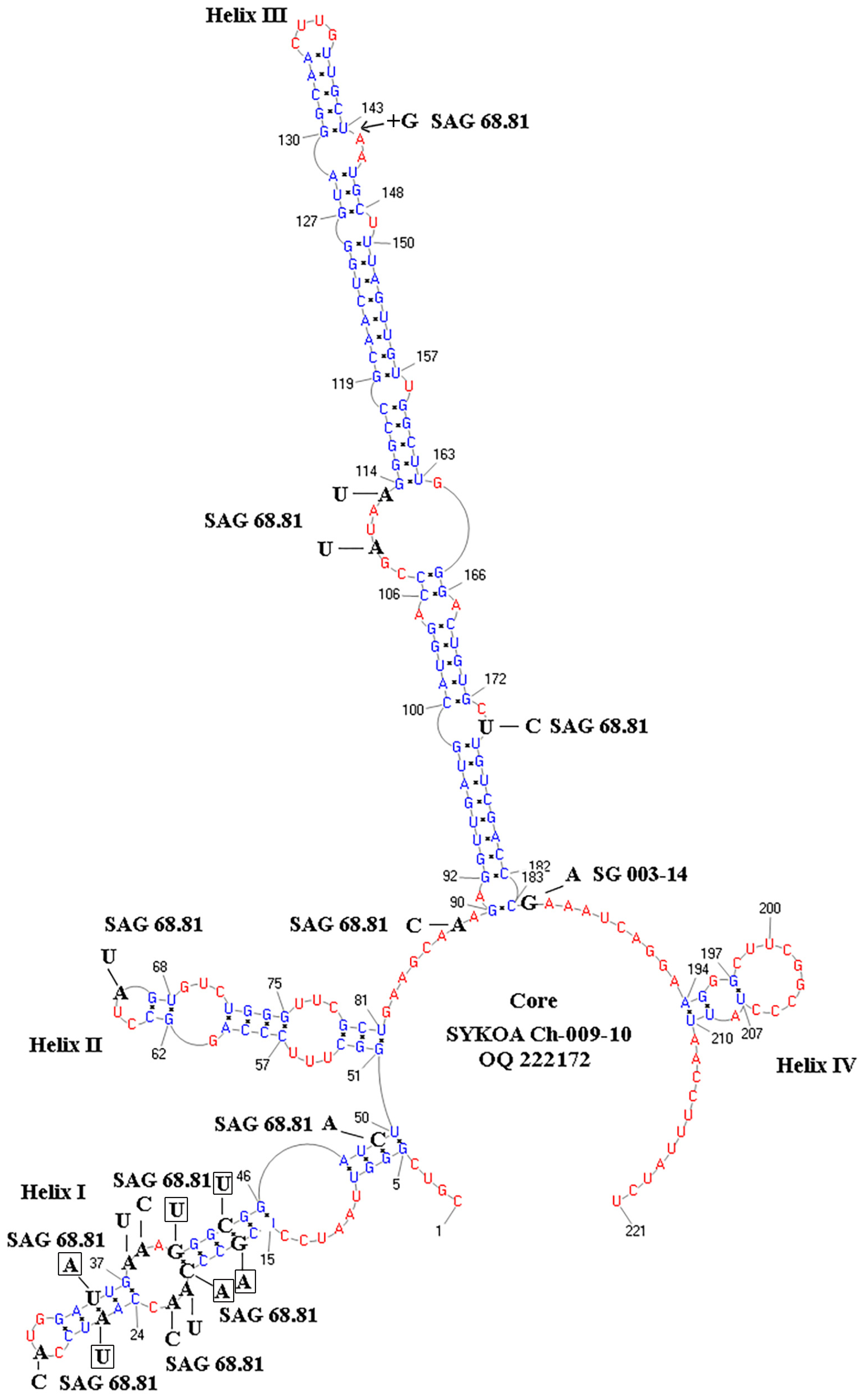
2.4. Modeling the Secondary Structure of the ITS2 Region
3. Results
3.1. Characteristics of Habitat Conditions
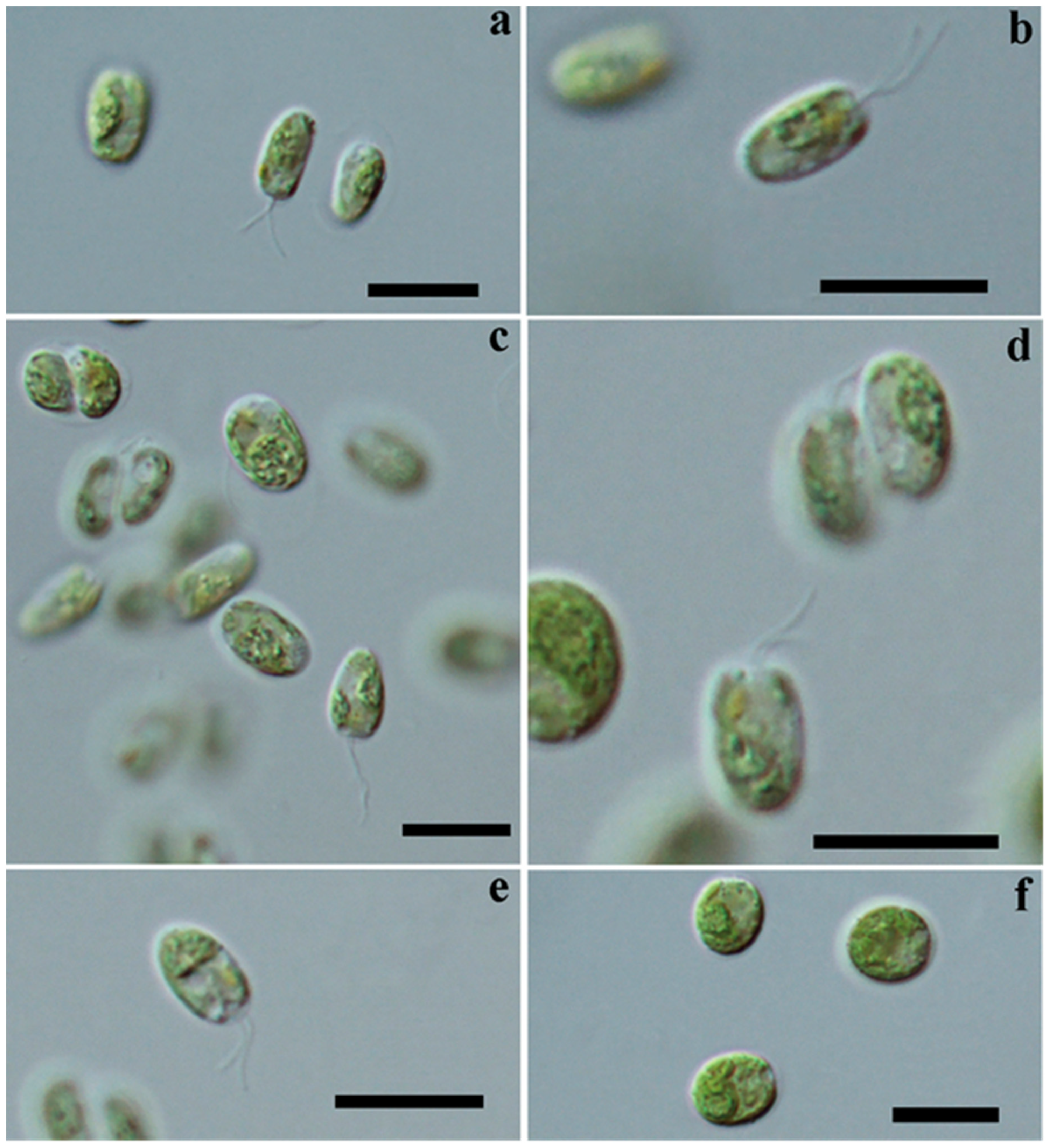
3.2. Cell Morphology: Light Microscopy
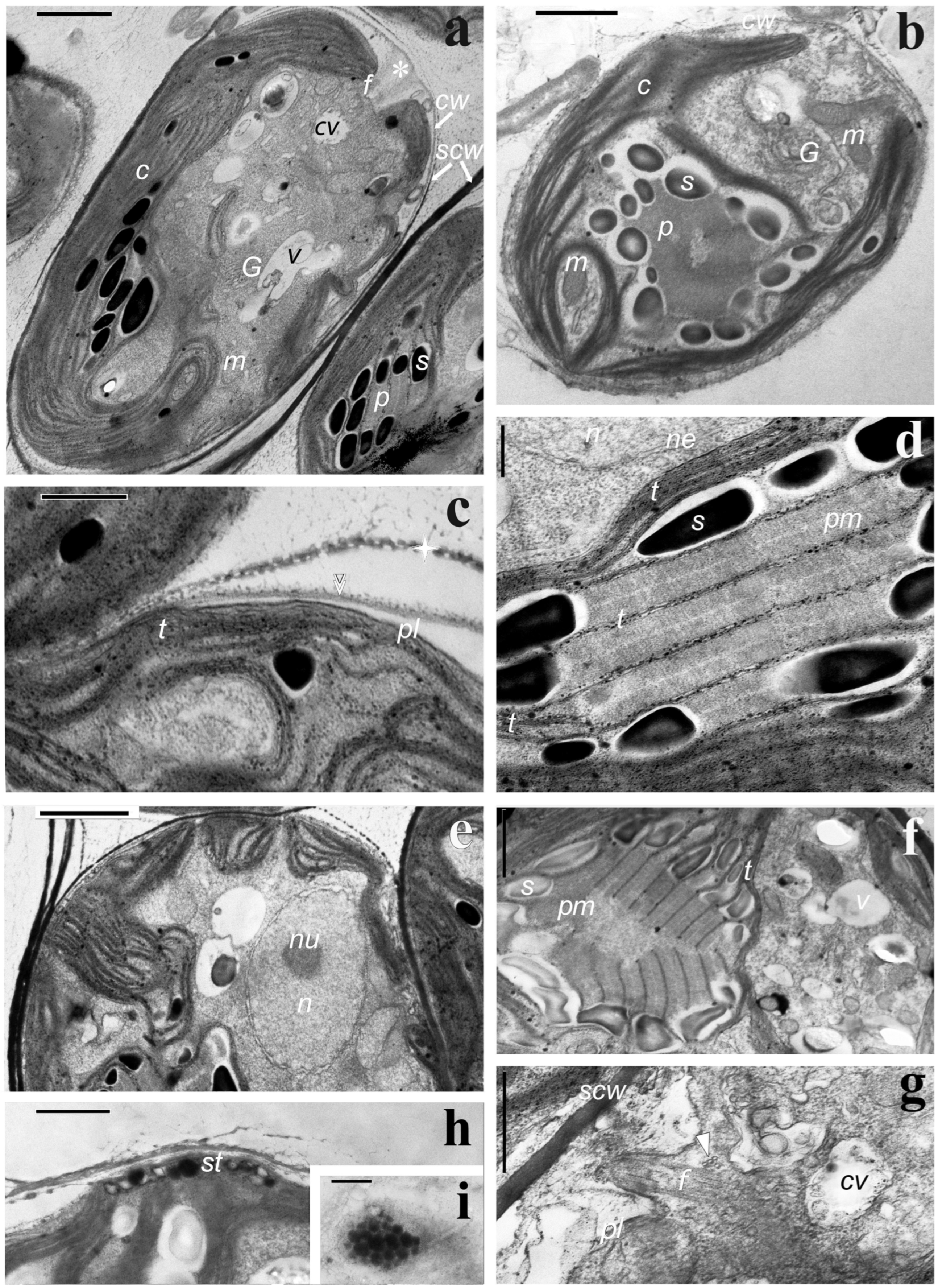
3.3. Cell Ultrastructure. Transmission Electron Microscopy
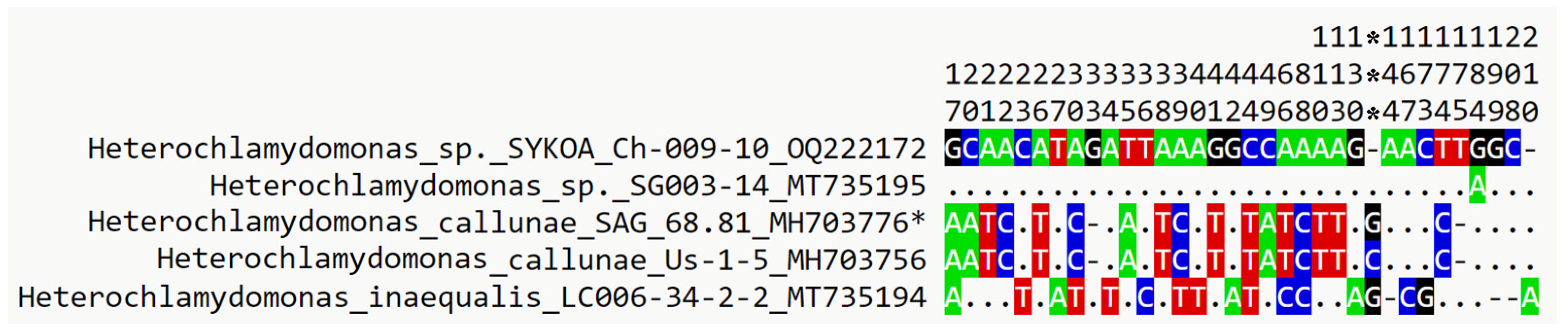
3.4. Molecular Phylogenetic Analysis
4. Discussion
Taxonomic Assessment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cox, E.R.; Deason, T.R. Heterochlamydomonas, a new alga from Tennessee. J. Tenn. Acad. Sci. 1969, 44, 105–107. [Google Scholar]
- Shoup, S.; Lewis, L.A. Polyphyletic origin of parallel basal bodies in swimming cells of chlorophycean green algae (Chlorophyta). J. Phycol. 2003, 39, 789–796. [Google Scholar] [CrossRef]
- Watanabe, S. Reclassification of Chlamydomonas monticola as Heterochlamydomonas (Volvocales, Chlorophyceae). Phycol. Res. 2020, 68, 332–335. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2023. [Google Scholar]
- Langford, M.F.; Cox, E.R. Additions to the genus Heterochlamydomonas (Chlorophycophyta). J. Tenn. Acad. Sci. 1971, 46, 66–71. [Google Scholar]
- Mikhailyuk, T.; Glaser, K.; Tsarenko, P.; Demchenko, E.; Karsten, U. Composition of biological soil crusts from sand dunes of the Baltic Sea coast in the context of an integrative approach to the taxonomy of microalgae and cyanobacteria. Eur. J. Phycol. 2019, 54, 263–290. [Google Scholar] [CrossRef]
- Ettl, H. Die Gattung Chlamydomonas Ehrenberg. Beih. Zur Nova Hedwig. 1976, 49, 1–1122. [Google Scholar]
- Watanabe, S. New and interesting green algae from soils of some Asian and Oceanian regions. Arch. Für Protistenkd. 1983, 127, 223–270. [Google Scholar] [CrossRef]
- Ettl, H.; Gärtner, G. Syllabus der Boden-, Luft-und Flechtenalgen; Auflage 2; Springer: Heidelberg/Berlin, Germany, 2014; p. 773. [Google Scholar]
- Kostikov, I.Y.; Romanenko, P.O.; Demchenko, E.M.; Darienko, T.M.; Mikhailyuk, T.I.; Rybchinskii, O.V.; Solonenko, A.M. Soil Algae of Ukraine: History and Methods of Analysis, System, and List of Flora; Fitosotsiotsentr: Kiev, Ukraine, 2001; p. 300. [Google Scholar]
- Zimonina, N.M. Soil Algae of the Oil Contaminated Land; VSPU: Kirov, Russia, 1998; p. 170. (In Russian) [Google Scholar]
- Patova, E.N.; Novakovskaya, I.V. Soil algae of the Northeastern European Russia. Nov. Sist. Nizshikh Rastenii 2018, 52, 311–353. [Google Scholar] [CrossRef]
- Watanabe, S.; Floyd, G.L. Comparative ultrastructure of zoospores with parallel basal bodies from the green algae Dictyochloris fragrans and Bracteacoccus sp. Am. J. Bot. 1992, 79, 551–555. [Google Scholar] [CrossRef]
- Nakada, T.; Misawa, K.; Nozaki, H. Molecular systematics of Volvocales (Chlorophyceae, Chlorophyta) based on exhaustive 18S rRNA phylogenetic analyses. Mol. Phylogenetics Evol. 2008, 48, 281–291. [Google Scholar] [CrossRef]
- Floyd, G.L.; Wilcox, L.W.; Watanabe, S. Structure and flagellar beat in Heterochlamydomonas inaequalis, a motile, unicellular, green alga with unequal flagella and parallel basal bodies. Crypt. Bot. 1990, 1, 332–339. [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: New York, NY, USA, 2005; p. 589. [Google Scholar]
- Andreeva, V.M. Soil and Aerophilic Green Algae (Chlorophyta: Tetrasporales Chlorococcales, Chlorosarcinales); Nauka: St. Petersburg, Russia, 1998; p. 351. (In Russian) [Google Scholar]
- Konstantinova, I.A.; Boldina, O.N. Comparative analysis of the pyrenoid ultrastructure in green monad and coccoid algae. Russ. J. Plant Physiol. 2000, 47, 655–659. [Google Scholar]
- Nakazawa, A.; Nozaki, H. Phylogenetic analysis of the Tetrasporalean genus Asterococcus (Chlorophyceae) based on 18S ribosomal RNA gene sequences. J. Jpn. Bot. 2004, 79, 255–261. [Google Scholar]
- Nakada, T.; Nozaki, H. Re-evaluation of three Chlorogonium (Volvocales, Chlorophyceae) species based on 18S ribosomal RNA gene phylogeny. Eur. J. Phycol. 2007, 42, 177–182. [Google Scholar] [CrossRef]
- Nakada, T.; Shinkawa, H.; Ito, T.; Tomita, M. Recharacterization of Chlamydomonas reinhardtii and its relatives with new isolates from Japan. J. Plant Res. 2010, 123, 67–78. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Ed.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Schultz, J.; Müller, T.; Achtziger, M.; Seibel, P.N.; Dandekar, T.; Wolf, M. The internal transcribed spacer 2 database-a web server for (not only) low level phylogenetic analyses. Nucleic Acids Res. 2006, 34, 704–707. [Google Scholar] [CrossRef]
- Ankenbrand, M.J.; Keller, A.; Wolf, M.; Schultz, J.; Förster, F. ITS2 database V: Twice as much. Mol. Biol. Evol. 2015, 32, 3030–3032. [Google Scholar] [CrossRef]
- Caisová, L.; Marin, B.; Melkonian, M.A. Consensus Secondary Structure of ITS2 in the Chlorophyta Identied by Phylogenetic Reconstruction. Protist 2013, 164, 482–486. [Google Scholar] [CrossRef]
- Temraleeva, A.D.; Mincheva, E.V.; Bukin, Y.S.; Andreeva, A.M. Modern Methods of Isolation, Cultivation and Identification of Green Algae (Chlorophyta); Kostroma Printing House: Kostroma, Russia, 2014; 215p. (In Russian) [Google Scholar]
- Byun, Y.; Han, K. PseudoViewer: Web application and web service for visualizing RNA pseudoknots and secondary structures. Nucleic Acids Res. 2006, 34, 416–422. [Google Scholar] [CrossRef]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Schultz, J.; Wolf, M. 4SALE—A tool for synchronous RNA sequence and secondary structure alignment and editing. BMC Bioinform. 2006, 7, 498. [Google Scholar] [CrossRef]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Wolf, M. Synchronous visual analysis and editing of RNA sequence and secondary structure alignments using 4SALE. BMC Res. Notes 2008, 1, 91. [Google Scholar] [CrossRef]
- Coleman, A.W. Is there a molecular key to the level of ‘biological species’ in eukaryotes? A DNA guide. Mol. Phylogenetics. Evol. 2009, 50, 197–203. [Google Scholar] [CrossRef]
- Patova, E.N.; Novakovskaya, I.V.; Sivkova, M.D. Cyanobacteria and Algae in Biological Soil Crusts of Frost Boils in the Mountain Tundra of the Urals. Eurasian Soil Sci. 2023, 56, 184–197. [Google Scholar] [CrossRef]
- Samolov, E.; Baumann, K.; Büdel, B.; Jung, P.; Leinweber, P.; Mikhailyuk, T.; Karsten, U.; Glaser, K. Biodiversity of Algae and Cyanobacteria in Biological Soil Crusts Collected Along a Climatic Gradient in Chile Using an Integrative Approach. Microorganisms 2020, 8, 1047. [Google Scholar] [CrossRef]
- Roberts, K. Crystalline glycoprotein cell walls of algae: Their structure, composition and assembly. Philos. Trans. R. Soc. B 1974, 268, 129–146. [Google Scholar] [CrossRef]
- Coleman, A.W.; van Oppen, M.J.H. Secondary structure of the rRNA ITS2 region reveals key evolutionary patterns in acroporid corals. J. Mol. Evol. 2008, 67, 389–396. [Google Scholar] [CrossRef]
- Caisová, L.; Marin, B.; Melkonian, M. A close-up view on ITS2 evolution and speciation—A case study in the Ulvophyceae (Chlorophyta, Viridiplantae). BMC Evol. Biol. 2011, 11, 262. [Google Scholar] [CrossRef] [PubMed]







 —gap between positions 143 and 144.
—gap between positions 143 and 144.
—gap between positions 143 and 144.
—gap between positions 143 and 144.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Moisture, % | Temperature (Min.; Max.), °C | C, % | N, % | Fe, mg/kg | Mn, mg/kg | pH (H2O) | pH (KCl) | Exchangeable Cations, cmolc/kg | P2O5 mg/kg | K2O mg/kg | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | ||||||||||
| ω ± Δ | ω ± Δ | ω ± Δ | ω ± Δ | pH ± Δ | pH ± Δ | n ± Δ | n ± Δ | ω ± Δ | ω ± Δ | ||
| 29.7 | −19.0 +13.1 | 0.43 0.13 | 0.038 0.010 | 57 17 | 610 170 | 6.26 | 5.21 | 5.28 0.40 | 2.12 0.16 | 447.8 67.17 | 96.76 14.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novakovskaya, I.V.; Boldina, O.N.; Shadrin, D.M.; Patova, E.N. Heterochlamydomonas uralensis sp. nov. (Chlorophyta, Chlamydomonadaceae), New Species Described from the Mountain Tundra Community in the Subpolar Urals (Russia). Diversity 2023, 15, 673. https://doi.org/10.3390/d15050673
Novakovskaya IV, Boldina ON, Shadrin DM, Patova EN. Heterochlamydomonas uralensis sp. nov. (Chlorophyta, Chlamydomonadaceae), New Species Described from the Mountain Tundra Community in the Subpolar Urals (Russia). Diversity. 2023; 15(5):673. https://doi.org/10.3390/d15050673
Chicago/Turabian StyleNovakovskaya, Irina Vladimirovna, Olga Nikolaevna Boldina, Dmitry Mikhailovich Shadrin, and Elena Nikolaevna Patova. 2023. "Heterochlamydomonas uralensis sp. nov. (Chlorophyta, Chlamydomonadaceae), New Species Described from the Mountain Tundra Community in the Subpolar Urals (Russia)" Diversity 15, no. 5: 673. https://doi.org/10.3390/d15050673
APA StyleNovakovskaya, I. V., Boldina, O. N., Shadrin, D. M., & Patova, E. N. (2023). Heterochlamydomonas uralensis sp. nov. (Chlorophyta, Chlamydomonadaceae), New Species Described from the Mountain Tundra Community in the Subpolar Urals (Russia). Diversity, 15(5), 673. https://doi.org/10.3390/d15050673