

Importance of Bark Physicochemical Properties in an Epiphytic Bryophyte Community within a Temperate Deciduous Broadleaf Forest

Abstract
:1. Introduction
2. Materials and Methods
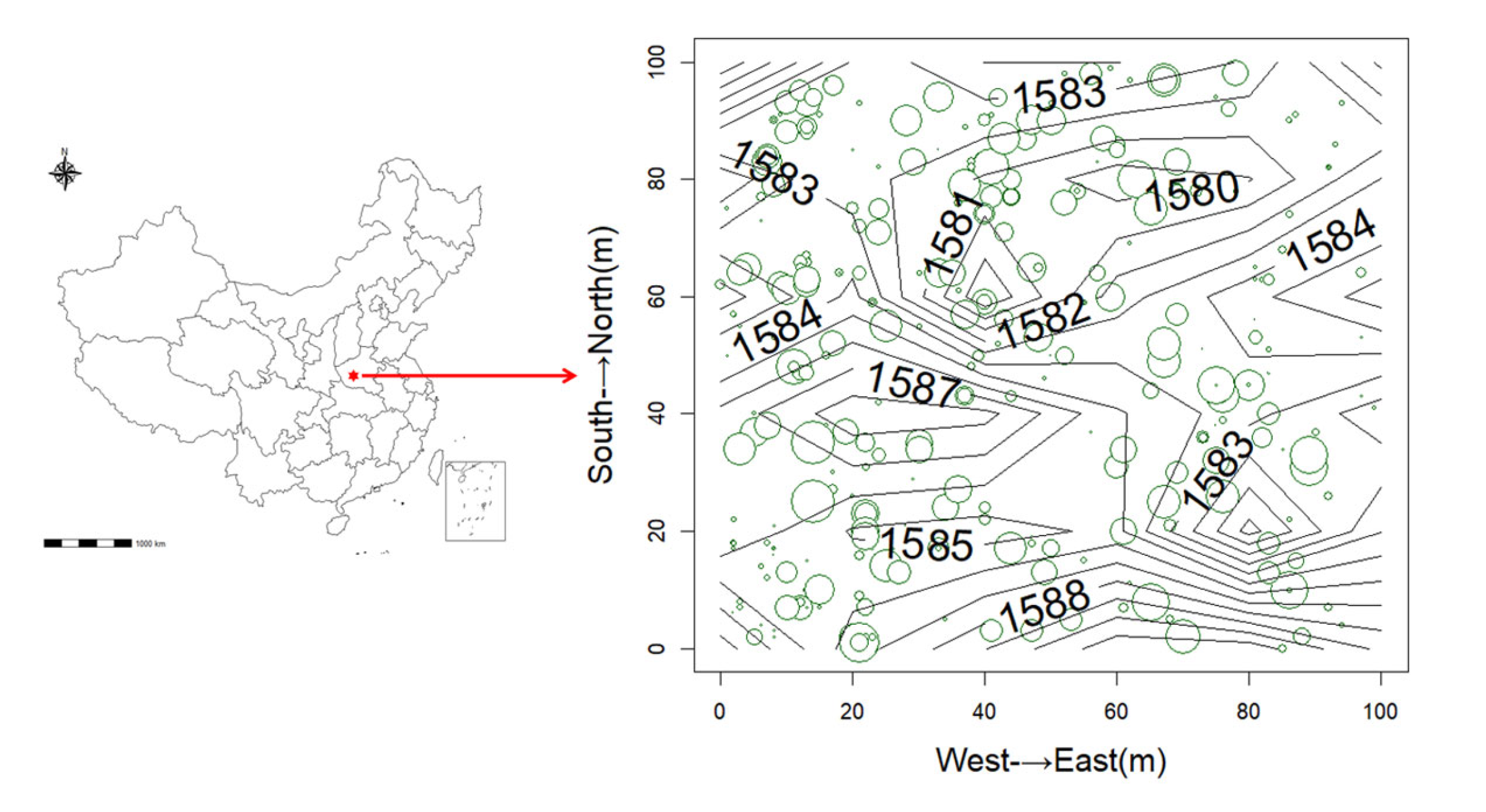
2.1. Study Location and Sampling Design
2.2. Epiphytic Bryophyte DNA and Functional-Trait Data Collection
2.3. Environmental Data
2.4. Statistical Procedures
3. Results
3.1. Composition of the Epiphytic Bryophyte Community
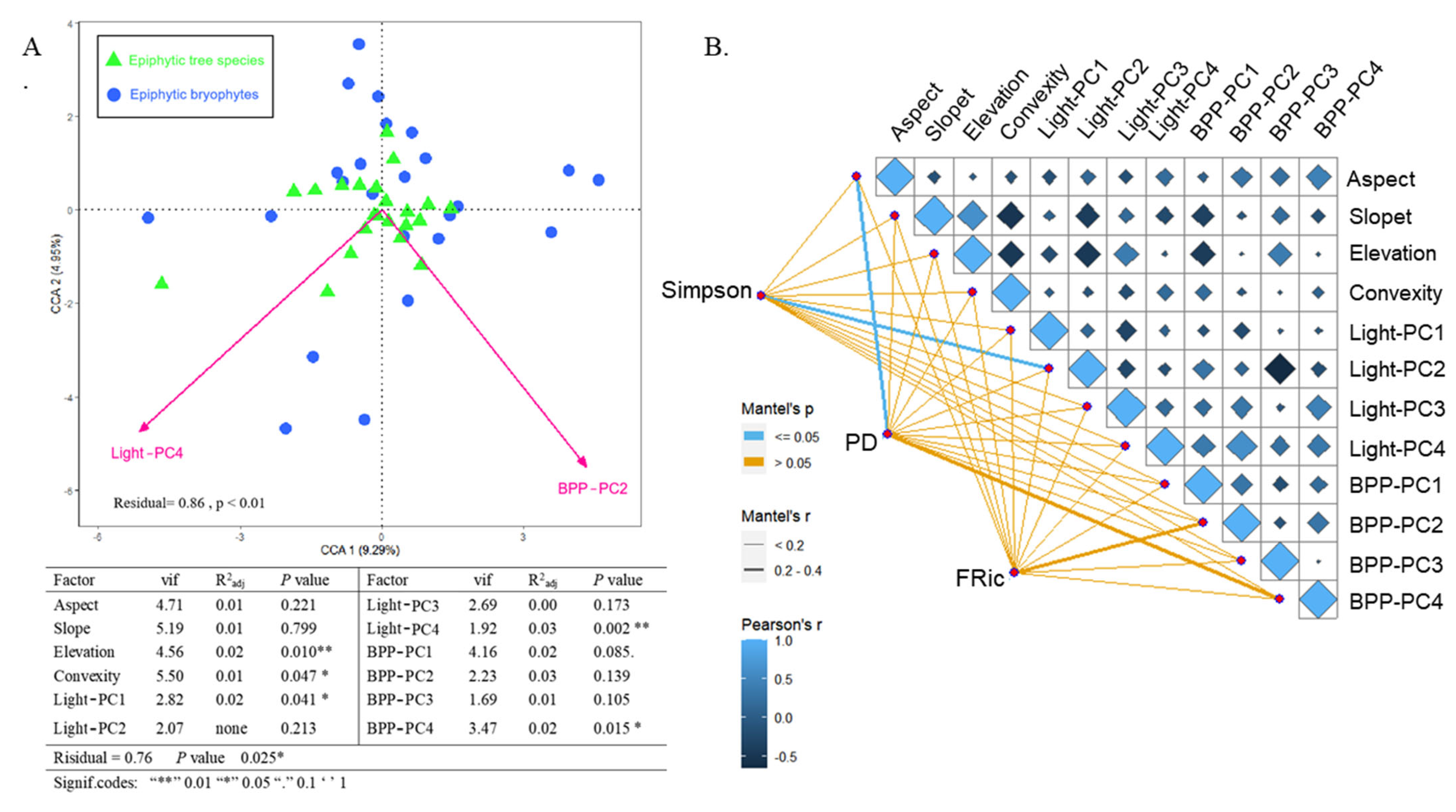
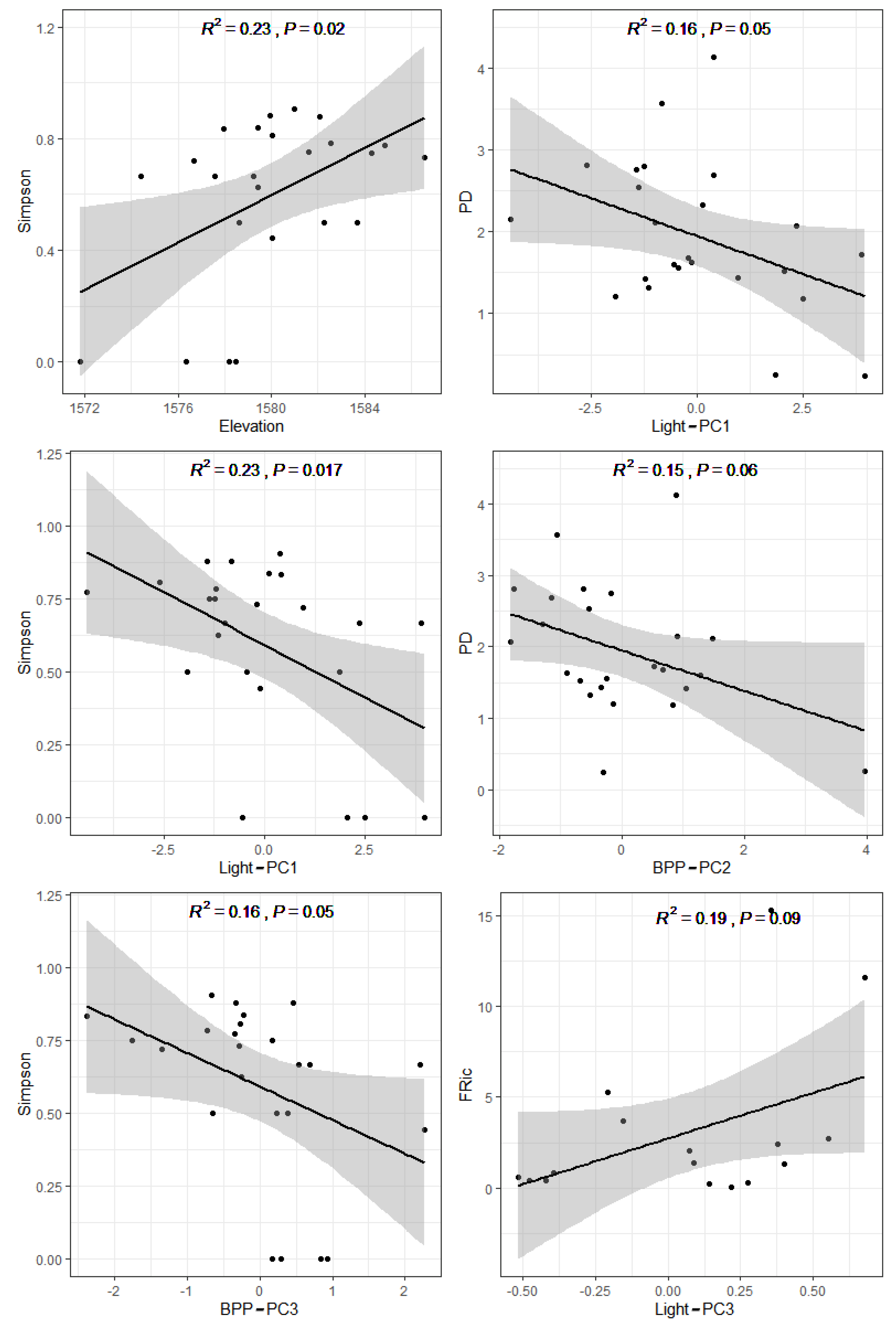
3.2. Effect of Environmental Variables on the Epiphytic Bryophyte Community
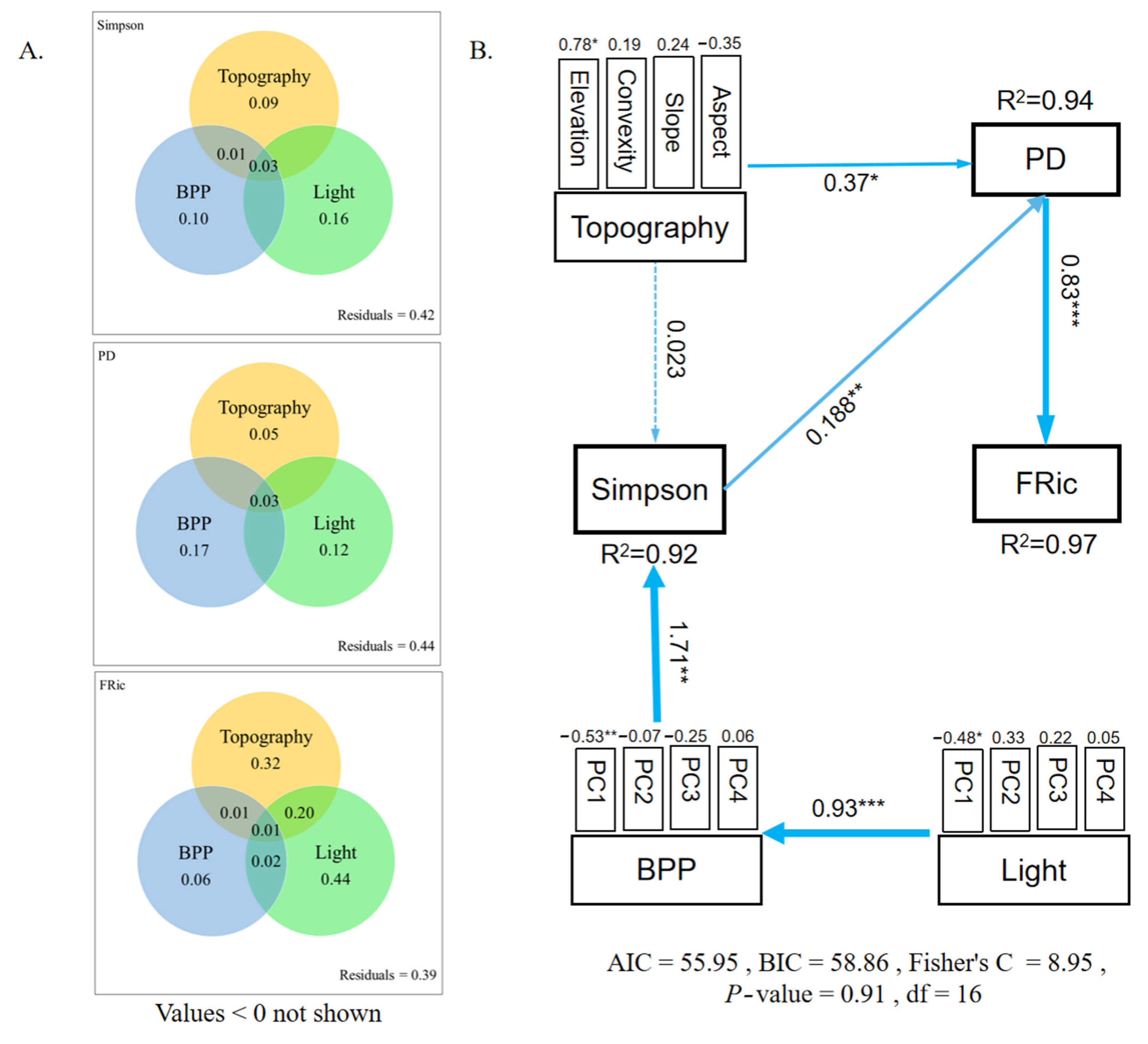
3.3. The Main Environmental Processes Determining the Coexistence of Bryophyte Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Goffnet, B.; Buck, W.R.; Shaw, A.J. Morphology, anatomy, and classification of the Bryophyta. Bryophyt. Biol. 2009, 2, 55–138. [Google Scholar]
- Gao, B.; Li, X.; Zhang, D.; Liang, Y.; Yang, H.; Chen, M.; Zhang, Y.; Zhang, J.; Wood, A.J. Desiccation tolerance in bryophytes: The dehydration and rehydration transcriptomes in the desiccation-tolerant bryophyte Bryum argenteum. Sci. Rep. 2017, 7, 7571. [Google Scholar] [CrossRef] [PubMed]
- Ehbrecht, M.; Seidel, D.; Annighöfer, P.; Kreft, H.; Köhler, M.; Zemp, D.C.; Puettmann, K.; Nilus, R.; Babweteera, F.; Willim, K.; et al. Global patterns and climatic controls of forest structural complexity. Nat. Commun. 2021, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Bowler, D.E.; Nilsen, E.B.; Bischof, R.; O’Hara, R.B.; Yu, T.T.; Oo, T.; Aung, M.; Linnell, J.D. Integrating data from different survey types for population monitoring of an endangered species: The case of the Eld’s deer. Sci. Rep. 2019, 9, 7766. [Google Scholar] [CrossRef]
- Slack, N.G. The Ecological Value of Bryophytes as Indicators of Climate Change. In Bryophyte Ecology and Climate Change; Cambridge University Press: Cambridge, UK, 2011; pp. 3–12. [Google Scholar]
- Kuusinen, M. Epiphyte Flora and Diversity on Basal Trunks of Six Old-Growth Forest Tree Species in Southern and Middle Boreal Finland. Lichenologist 1996, 28, 443–463. [Google Scholar] [CrossRef]
- Slack, N.G. Host specificity of bryophytic epiphytes in Eastern North America. J. Hattori Bot. Lab. 1976, 41, 107–132. [Google Scholar]
- Studlar, S.M. Host Specificity of Epiphytic Bryophytes near Mountain Lake, Virginia. Bryologist 1982, 85, 37–50. [Google Scholar] [CrossRef]
- Yu, S.; Zinati, G.M. Physical and chemical changes in container media in response to bark substitution for peat. Compost. Sci. Util. 2006, 14, 222–230. [Google Scholar] [CrossRef]
- Chemetova, C.; Quilhó, T.; Braga, S.; Fabio, A.M.; Ribeiro, D. Aged Acacia melanoxylon bark as an organic peat replacement in container media. J. Clean. Prod. 2019, 232, 1103–1111. [Google Scholar] [CrossRef]
- Pescador, D.S.; de Bello, F.; Valladares, F.; Escudero, A.; Weigelt, A. Plant Trait Variation along an Altitudinal Gradient in Mediterranean High Mountain Grasslands: Controlling the Species Turnover Effect. PLoS ONE 2015, 10, e0118876. [Google Scholar] [CrossRef]
- William, K.; Cornwell, D.W.; Schwilk, D.D.; Ackerly, A. Trait-Based Test for Habitat Filtering: Convex Hull Volume. Ecology 2006, 87, 1465–1471. [Google Scholar]
- Swenson, N.G.; Enquist, B.J.; Thompson, J.; Zimmerman, J.K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology 2007, 88, 1770–1780. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Renato, V.D.D. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.C.; Hernández, C.; Romoleroux, K.; Losos, E.; Magård, E.; Balslev, H. Tree Species Distributions and Local Habitat Variation in the Amazon: Large Forest Plot in Eastern Ecuador. J. Ecol. 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Opposing assembly mechanisms in a Neotropical dry forest: Implications for phylogenetic and functional community ecology. Ecology 2009, 90, 2161–2170. [Google Scholar] [CrossRef]
- Uriarte, M.; Swenson, N.G.; Chazdon, R.L.; Comita, L.S.; John Kress, W.; Erickson, D.; Forero-Montaña, J.; Zimmerman, J.K.; Thompson, J. Trait similarity, shared ancestry and the structure of neighbourhood interactions in a subtropical wet forest: Implications for community assembly. Ecol. Lett. 2010, 13, 1503–1514. [Google Scholar] [CrossRef]
- Hoffmann, S.; Beierkuhnlein, C.; Field, R.; Provenzale, A.; Chiarucci, A. Uniqueness of Protected Areas for Conservation Strategies in the European Union. Sci. Rep. 2018, 8, 6445. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Swenson, N.G.; Thompson, J.; Uriarte, M.; Zimmerman, J.K. Advances in the Use of DNA Barcodes to Build a Community Phylogeny for Tropical Trees in a Puerto Rican Forest Dynamics Plot. PLoS ONE 2010, 5, e15409. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Harms, K.; Richard, E.; Stephen, C.; Hubbell, P.; Robin, B. Foster Habitat Associations of Trees and Shrubs in a 50-Ha Neotropical Forest Plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Han, B.; Umaña, M.N.; Mi, X.; Liu, X.; Chen, L.; Wang, Y.; Liang, Y.; Wei, W.; Ma, K. The role of transcriptomes linked with responses to light environment on seedling mortality in a subtropical forest, China. J. Ecol. 2017, 105, 592–601. [Google Scholar] [CrossRef]
- Zheng, Y.; Hou, L.; Newell, S.; Liu, M.; Zhou, J.; Zhao, H.; Cheng, X. Community Dynamics and Activity of Ammonia-Oxidizing Prokaryotes in Intertidal Sediments of the Yangtze Estuary. Appl. Environ. Microb. 2014, 80, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Conant, R.T.; Zhou, G.; Wang, Y.; Wang, N.; Li, Y.; Zhang, K. Effects of moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pools. Sci. Rep. 2016, 6, 31480. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, Z.; Li, Z.; Xue, R.; Cui, W.; Sun, S.; Liu, T.; Zeng, R.; Song, Y. Deficiency in Silicon Transporter Lsi1 Compromises Inducibility of Anti-herbivore Defense in Rice Plants. Front. Plant. Sci. 2019, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. Package ‘FD’. 2014: 1-0. Available online: https://www.imsbio.co.jp/RGM/R_rdfile?f=FD/man/FD-package.Rd&d=R_CC (accessed on 3 April 2022).
- Legras, G.; Loiseau, N.; Gaertner, J.C. Functional richness: Overview of indices and underlying concepts. ACTA Oecol. 2018, 87, 34–44. [Google Scholar] [CrossRef]
- Daniel, P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 10. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0. 2012. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1654-1103.2003.tb02228.x (accessed on 19 April 2022).
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning Beta Diversity in a Subtropical Broad-Leaved Forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef]
- Grace, J.B. Structural Equation Modeling Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Lefcheck, J.S. PiecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Tian, P.; Liu, S.; Zhao, X.; Sun, Z.; Yao, X.; Niu, S.; Crowther, T.W.; Wang, Q. Past climate conditions predict the influence of nitrogen enrichment on the temperature sensitivity of soil respiration. Commun. Earth Env. Environ. 2021, 2, 251. [Google Scholar] [CrossRef]
- Tinya, F.; Márialigeti, S.; Király, I.; Németh, B.; Odor, P. The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant. Ecol. 2009, 204, 69–81. [Google Scholar] [CrossRef]
- Kovářová, M.; Pyszko, P.; Plášek, V. How Does the pH of Tree Bark Change with the Presence of the Epiphytic Bryophytes from the Family Orthotrichaceae in the Interaction with Trunk Inclination? Plants 2022, 11, 63. [Google Scholar] [CrossRef]
- Liu, M.; Che, Y.; Jiao, J.; Li, L.; Jiang, X. Exploring the community phylogenetic structure along the slope aspect of subalpine meadows in the eastern Qinghai–Tibetan Plateau, China. Ecol. Evol. 2019, 9, 5270–5280. [Google Scholar] [CrossRef]
- Win, K.T.; Okazaki, K.; Ookawa, T.; Yokoyama, T.; Ohwaki, Y. Influence of rice-husk biochar and Bacillus pumilus strain TUAT-1 on yield, biomass production, and nutrient uptake in two forage rice genotypes. PLoS ONE 2019, 14, e0220236. [Google Scholar]
- Gil-Martínez, M.; López-García, Á.; Domínguez, M.T.; Navarro-Fernández, C.M.; Kjøller, R.; Tibbett, M.; Marañón, T. Ectomycorrhizal Fungal Communities and Their Functional Traits Mediate Plant–Soil Interactions in Trace Element Contaminated Soils. Front. Plant Sci. 2018, 9, 1682. [Google Scholar] [CrossRef]
- Becker, D.; Dobson, A.; Klitgaard, K. Bark characteristics affect epiphytic bryophyte cover across tree species. In California Ecology and Conservation Research; University of California: Riverside, CA, USA, 2019; pp. 1–8. [Google Scholar]
- Schmitt, C.K.; Slack, N.G. Host specificity of epiphytic lichens and bryophyte: A comparison of the Adirondack Mountains (New York) and the Southern Blue Ridge Mountains (North Carolina). Bryologist 1990, 93, 661–665. [Google Scholar] [CrossRef]
- Trynoski, S.E.; Glime, J.M. Direction and height of bryophytes on four species of Northern Trees. Bryologist 1982, 85, 281–300. [Google Scholar] [CrossRef]
- González-Mancebo, J.M.; Romaguera, F.; Losada-Lima, A.; Suárez, A. Epiphytic bryophytes growing on Laurus azorica (Seub.) Franco in three laurel forest area in Tenerife (CanaryIs lands). ACTA Oecol. 2004, 25, 159–167. [Google Scholar] [CrossRef]
- González-Mancebo, J.M.; Losada-Lima, A.; McAlister, S. Host specificity of epiphytic bryophyte communities of a Laurel forest on Tenerife (Canary Islands, Spain). Bryologist 2003, 106, 383–394. [Google Scholar] [CrossRef]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef]
- Palmer, M.W. Pattern in Corticolous Bryophyte Communities of the North Carolina Piedmont: Do Mosses See the Forest or the Trees? Bryologist 1986, 89, 59–65. [Google Scholar] [CrossRef]
- Weiqiu, L.; Dai, X.; Wang, Y.; Lei, C. Analysis of environmental factors affecting thedistribution of epiphytic bryophyte at Heishiding Nature Reserve, GuangdongProvince. ShengTaiXueBao 2008, 28, 1080–1088. [Google Scholar]
- Turetsky, M.R. The Role of Bryophytes in Carbon and Nitrogen Cycling. Bryologist 2003, 106, 395–409. [Google Scholar] [CrossRef]
- Glime, J.M. Physiological Ecology. Vol.1. In Bryophyte Ecology; Michigan Technological University and the International Association of Bryologists: Houghton, MI, USA, 2007; Available online: http://www.bryoecol.mtu.edu (accessed on 10 April 2022).
- Steele, A. Bryophyte communities of central Idaho forests. Northwest Sci. 1978, 52, 310–322. [Google Scholar]
- Kenkel, N.C.; Bradfield, G.E. Ordination of Epiphytic Bryophyte Communities in a Wet-Temperate Coniferous Forest, South-Coastal British Columbia. Vegetatio 1981, 45, 147–154. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Epiphytic Bryophytes |
|---|---|
| Quercus aliena Bl. var. acuteserrata Maxim. ex Wenz. | Pylaisiadelpha yokohamae (Broth.) Buck. |
| Pinus armandii Franch. | Eurohypnum leptothallum (Müll. Hal.) Ando |
| Toxicodendron vernicifluum (Stokes) F.A.Barkl. | Pylaisiadelpha tenuirostris (Bruch et Schimp. ex Sull.) Buck |
| Betula platyphylla Sukaczev | Gammiella panchienii Tan et Jia |
| Lindera obtusiloba Blume | Eurhynchium latifolium Cardot |
| Litsea tsinlingensis Yen C. Yang & P. H. Huang | Frullania sinensis Steph. |
| Cornus kousa subsp. chinensis | Entodon macropodus (Hedw.) Müll. Hal. |
| Corylus heterophylla var. sutchuenensi Franch. | Haplohymenium triste (Ces.) Kindb. |
| Symplocos paniculata (Thunb.) Miq. | Heteroscyphus argutus (Reinw. et al.) Schiffn. |
| Carpinus turczaninowii Hance | Schwetschkeopsis fabronia (Schwaegr.) Broth. |
| Tilia paucicostata Maxim. | Brachythecium rivulare Schimp. |
| Sorbus alnifolia (Sieb. et Zucc.) K. Koch | Frullania ericoides (Nees) Nees & Mont. |
| Malus honanensis Rehder | Pylaisiopsis speciosa (Mitt.) Broth. |
| Prunus clarofolia C. K. Schneid. | Gollania ruginosa (Mitt.) Broth. |
| Quercus variabilis Bl. | Mnium laevinerve Cardot. |
| Tilia japonica (Miq.) Simonk. | Taxiphyllum taxirameum (Mitt.) M. Fleisch. |
| Cornus controversa Hemsl. | Pylaisia polyantha (Hedw.) Schimp. |
| Viburnum betulifolium Batal. | Rhynchostegium pallenticaule Müll. Hal. |
| Meliosma veitchiorum Hemsl. | Homalothecium sericeum (Hedw.) Bruch, Schimp. & W.Guembel |
| Pinus tabuliformis Carrière | Trocholejeunea sandvicensis (Gott.) Mizut. |
| Carpinus cordata Bl. | Trichostomum tenuirostre Lindb |
| Styrax obassis Siebold et Zucc. | Brachythecium buchananii Jaeger |
| Juglans cathayensis Dode | Plagiomnium cuspidatum T. Kop. |
| Celastrus orbiculatus Thunb. | Entodon cladorrhizans (Hedwig) Müller Hal. |
| Bryhnia brachycladula Cardot |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Wang, S.; Li, Y.; Chen, Y.; Zhao, H.; Wang, J.; Liu, F.; Yuan, Z. Importance of Bark Physicochemical Properties in an Epiphytic Bryophyte Community within a Temperate Deciduous Broadleaf Forest. Diversity 2023, 15, 688. https://doi.org/10.3390/d15050688
Shao Y, Wang S, Li Y, Chen Y, Zhao H, Wang J, Liu F, Yuan Z. Importance of Bark Physicochemical Properties in an Epiphytic Bryophyte Community within a Temperate Deciduous Broadleaf Forest. Diversity. 2023; 15(5):688. https://doi.org/10.3390/d15050688
Chicago/Turabian StyleShao, Yizhen, Senlin Wang, Yushan Li, Yun Chen, He Zhao, Jing Wang, Fengqin Liu, and Zhiliang Yuan. 2023. "Importance of Bark Physicochemical Properties in an Epiphytic Bryophyte Community within a Temperate Deciduous Broadleaf Forest" Diversity 15, no. 5: 688. https://doi.org/10.3390/d15050688
APA StyleShao, Y., Wang, S., Li, Y., Chen, Y., Zhao, H., Wang, J., Liu, F., & Yuan, Z. (2023). Importance of Bark Physicochemical Properties in an Epiphytic Bryophyte Community within a Temperate Deciduous Broadleaf Forest. Diversity, 15(5), 688. https://doi.org/10.3390/d15050688