

Non-Indigenous Species Dynamics in Time and Space within the Coastal Waters of The Netherlands
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analyses Based on NIS Recorded within the Coastal Waters of The Netherlands
2.2. Pathways, Vectors, and Their Uncertainties
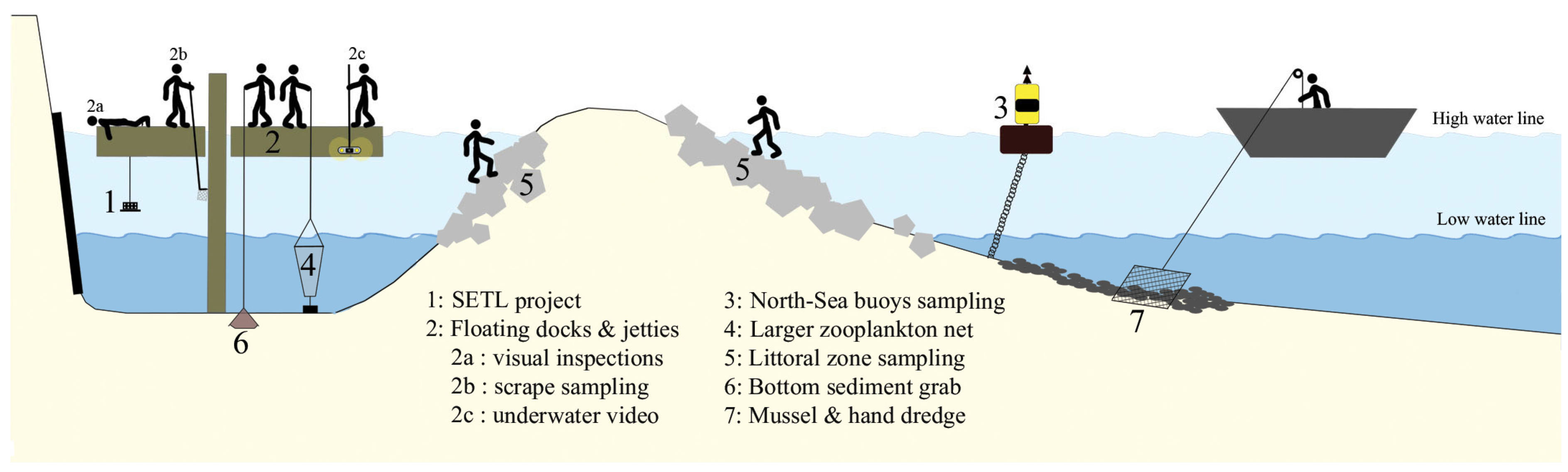
2.3. Marine Alien Species Detection Network


2.4. Further Data Collection
3. Results and Discussion
3.1. Non-Indigenous Species Dynamics in Time and Space: Origins, Pathways, and Vectors
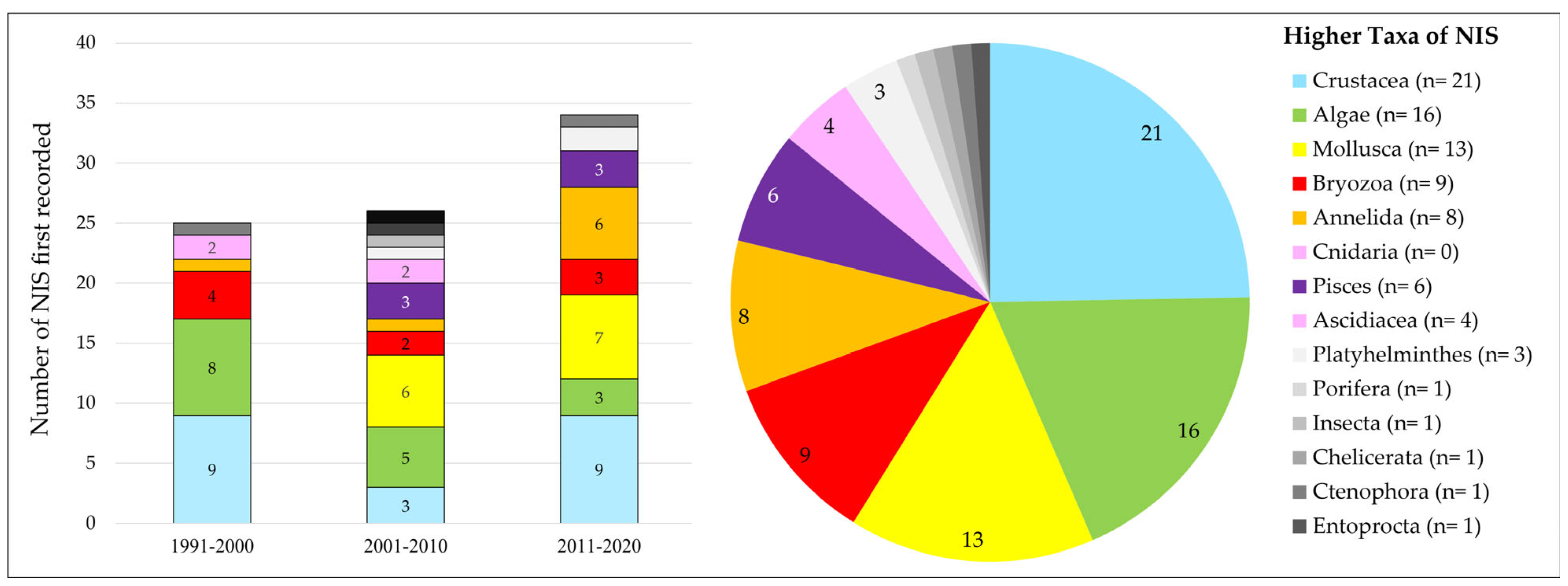
3.1.1. NIS Taxa Recorded over Time in the Coastal Waters of The Netherlands
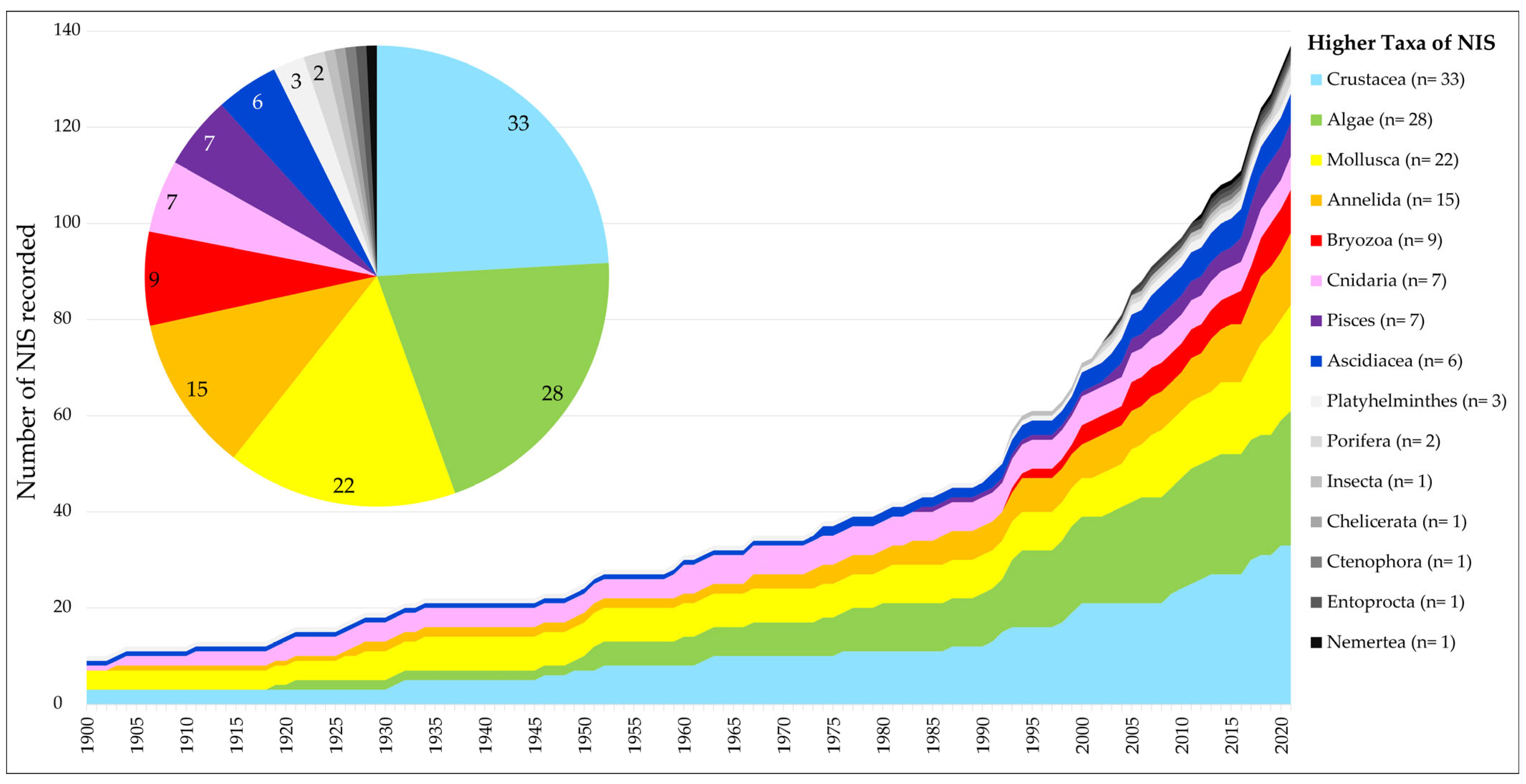
- Crustacea, followed by the (macro-)Algae and the Mollusca, form the main taxa of which NIS are recorded in the coastal waters.
- The number of recorded NIS has shown a steady increase from the early 20th century up to the early 1990s. From the 1990s on, the number of new marine NIS that were recorded for The Netherlands on an annual basis increased. Although this increase is clear for NIS from various higher taxonomical groups, bryozoans seem to play an exceptional role. While no records of alien bryozoan species are known from before the 1990s, they have become an increasingly important group of NIS since the 1990s. This is also clear in Figure 5, illustrating that 6 out of the 10 alien bryozoan species recorded were first recorded in the last decade, i.e., between 2011 and 2020. Although this may partly be explained by a recently increased research effort regarding bryozoans, it may also be linked to hull fouling becoming the most important introduction vector of marine NIS in recent years, as is also discussed in paragraph 3.1.4. Whatever the underlying reason, the sudden increase in the annual rate of new NIS discoveries appears typical for the Greater North Sea area in general [9,11]. In other NE Atlantic regions, i.e., the Bay of Biscay–Iberian Shelf, Celtic Seas, and Macaronesia, no such increase was apparent from the 1990s on [9].
- The main higher taxa of the NIS that were recorded as new to The Netherlands differed over the last three decades (Figure 5). These differences cannot easily be linked to research effort. Molluscs, for example, have always been a prominent part of various monitoring programs. It is, therefore, unclear why not even a single new non-indigenous molluscan species was recorded for The Netherlands during 1991–2000, while in the following two decades, six and seven molluscan species, respectively, were recorded. Vice versa, there appears to have been a negative trend over the last three decades where it concerns the number of non-indigenous macro-algal species, with only two new NIS recorded in the most recent decade, i.e., 2011–2020 (Figure 5). One does have to be careful, however, with the interpretation of such trends, as in 2021 alone, two additional non-indigenous seaweed species were recorded: Kapraunia schneideri (Stuercke & Freshwater) Savoie & G.W. Saunders, 2019, and Neopyropia yezoensis (Ueda) L.-E. Yang & J. Brodie, 2020.
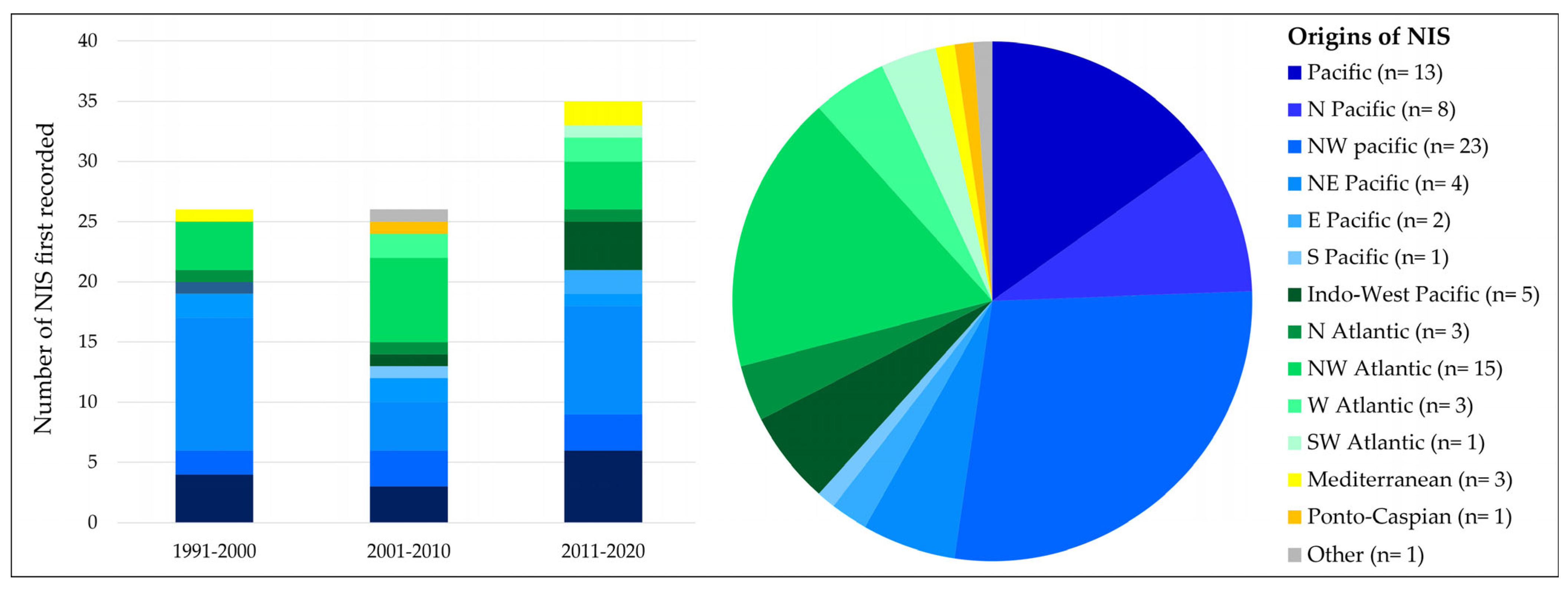
3.1.2. NIS Origins over Time
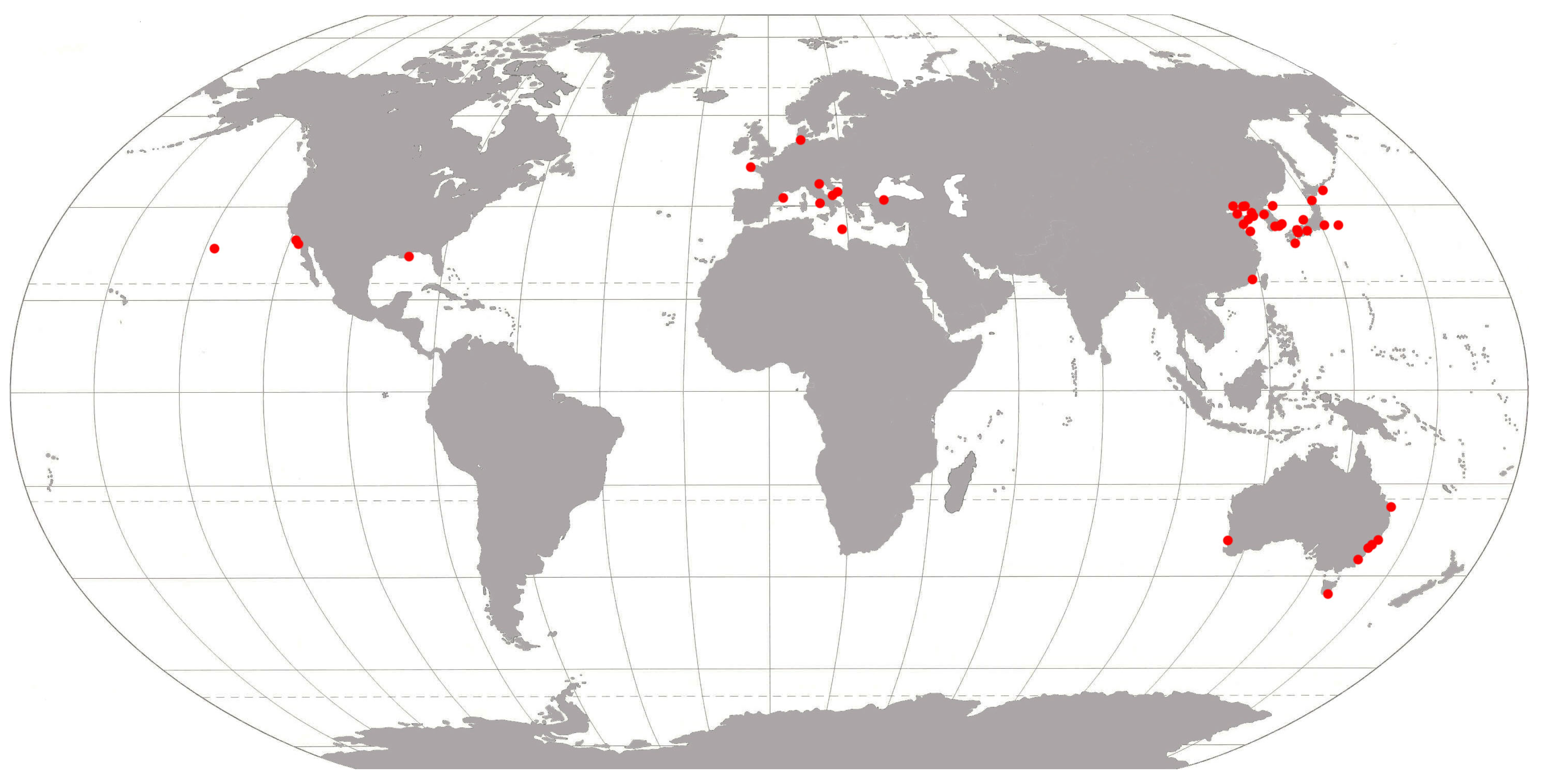
- A relatively small number of NIS originate from the Mediterranean and the Ponto-Caspian region. Although this may suggest that NIS are not often introduced from these seas, one should consider that the Mediterranean in particular can also function as a stepping stone for NIS primarily introduced there from the Pacific. For example, the cryptic moon jellyfish Aurelia coerulea von Lendenfeld, 1884, originates from the Pacific. It was recently found to be established in The Netherlands [23] and was most likely introduced by secondary spread from the Mediterranean Sea, where the species is known to be well-established [33]. Concerning the relatively low number of species from the Ponto-Caspian, it must be considered that most species coming from those seas prefer salinities lower than 5 ppt, and are, therefore, not included in the present study.
- Three decades ago, i.e., between 1991 and 2000, close to eighty percent of all newly recorded NIS originated from the Indo-Pacific. This is often linked [34] to the imports of the Pacific oyster Magallana gigas (Thunberg, 1793) directly from the Pacific, for example, by plane, in the 1970s and 1980s. Such Pacific oyster imports have no longer been allowed in most European countries since the 1990s, however, acknowledging the risk of NIS introductions. This may explain why a much lower percentage (about 50%) of all NIS that were first recorded in 2001–2010 originate from the Pacific.
- The relative decrease in the number of species originating from the Pacific in 2001–2010 does not explain why, in this decade, the absolute number of NIS originating from the W Atlantic had more than doubled.
- Finally, looking at the origins of species in the most recent decade, 2011–2020, it is surprising to see that the number of NIS that originate from the Pacific has increased again to about the same number as in 1991–2000. As Pacific oyster imports from other continents are no longer allowed, a different pathway (maybe hull fouling) and, thereby, vector must have been responsible for this increase.
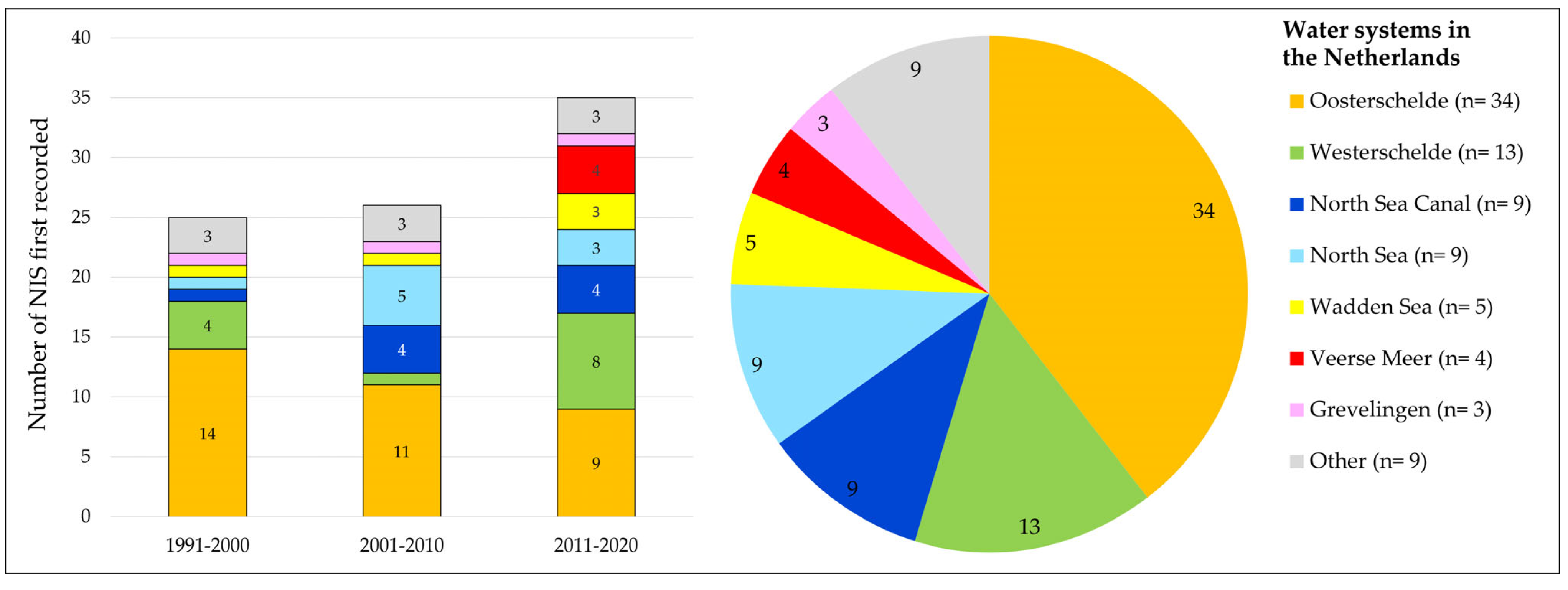
3.1.3. Water Systems Where NIS Were First Recorded
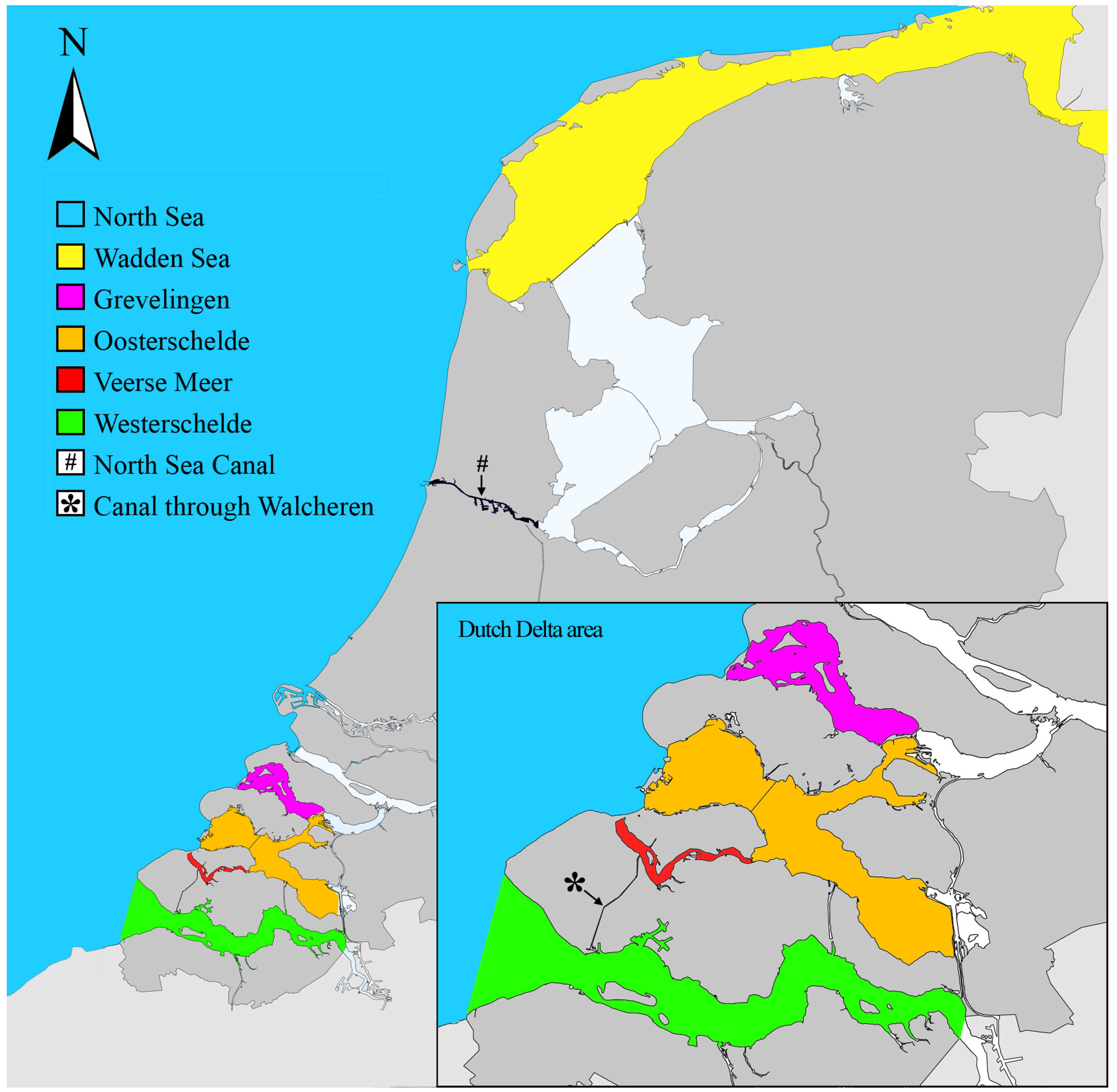
- The Oosterschelde, a Natura 2000 area, is the area in which most NIS were recorded for the first time in The Netherlands (Figure 7). However, the absolute number of new NIS recorded in the Oosterschelde has declined over the last 3 decades, from 14 in 1991–2000 to 9 in 2011–2020 (Figure 7). The relatively high number of NIS recorded in this water system may be due to a combination of causes:
- The Oosterschelde is the most popular Dutch area for scuba divers who are actively scoring the species present. These “citizen science” records, which also include NIS, are recorded by the ANEMOON Foundation and represent an important part of all new NIS recorded [31].
- Within the Oosterschelde, there are many marinas for pleasure crafts, which may act as introduction vectors for NIS.
- The Oosterschelde is an important area for shellfish transports, which may function as an introduction vector for NIS.
- Other water systems in The Netherlands are either brackish, have less hard substrata, and/or are in general less diverse in terms of micro-habitats, offering newcomers fewer niches to settle in.
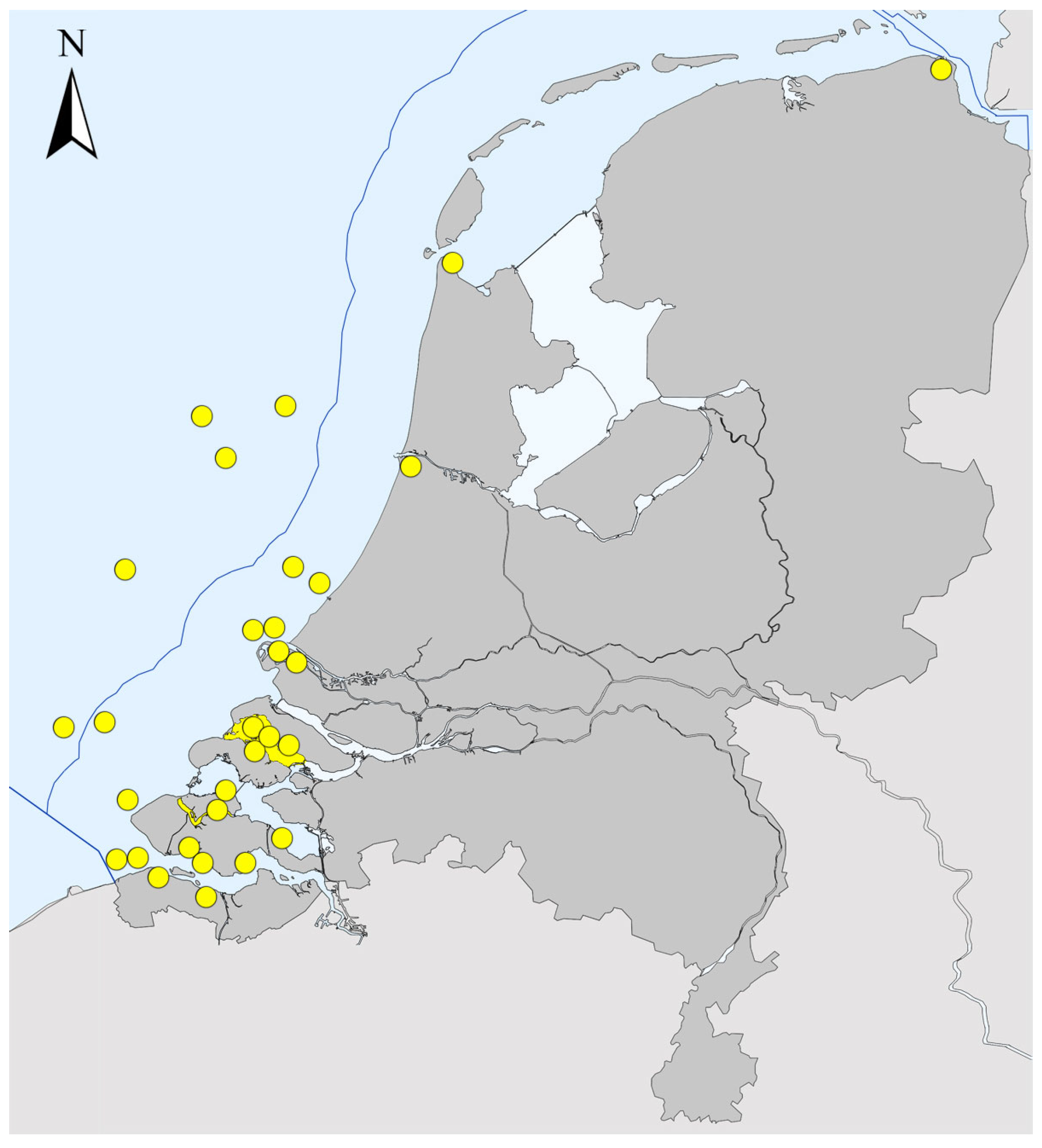
- Monitoring effort may explain in large part the trends over time visible in Figure 7. This becomes especially clear in the last period “2011–2020”, during which seven NIS new to The Netherlands were recorded in the Wadden Sea and Veerse Meer. This high number of new NIS records is directly linked to the NIS-focused surveys that were conducted in these waters since 2009 [16,17,18,19,20,23]. As these were the first NIS-focused surveys there, they resulted in the detection of a relatively high number of new NIS, for which it was not always clear how long they had already been established there.
- Concerning the number of new NIS recorded within the North Sea Canal (Figure 1 and Figure 7), it should be considered that more NIS than shown here, new to The Netherlands, were recorded. This concerns NIS that established themselves in parts of the canal with salinities <5 ppt, e.g., Laonome xeprovala Bick & Bastrop, 2018 (see [35]). As the present study focuses on species occurring in waters with salinities of >5 ppt, they were not included.
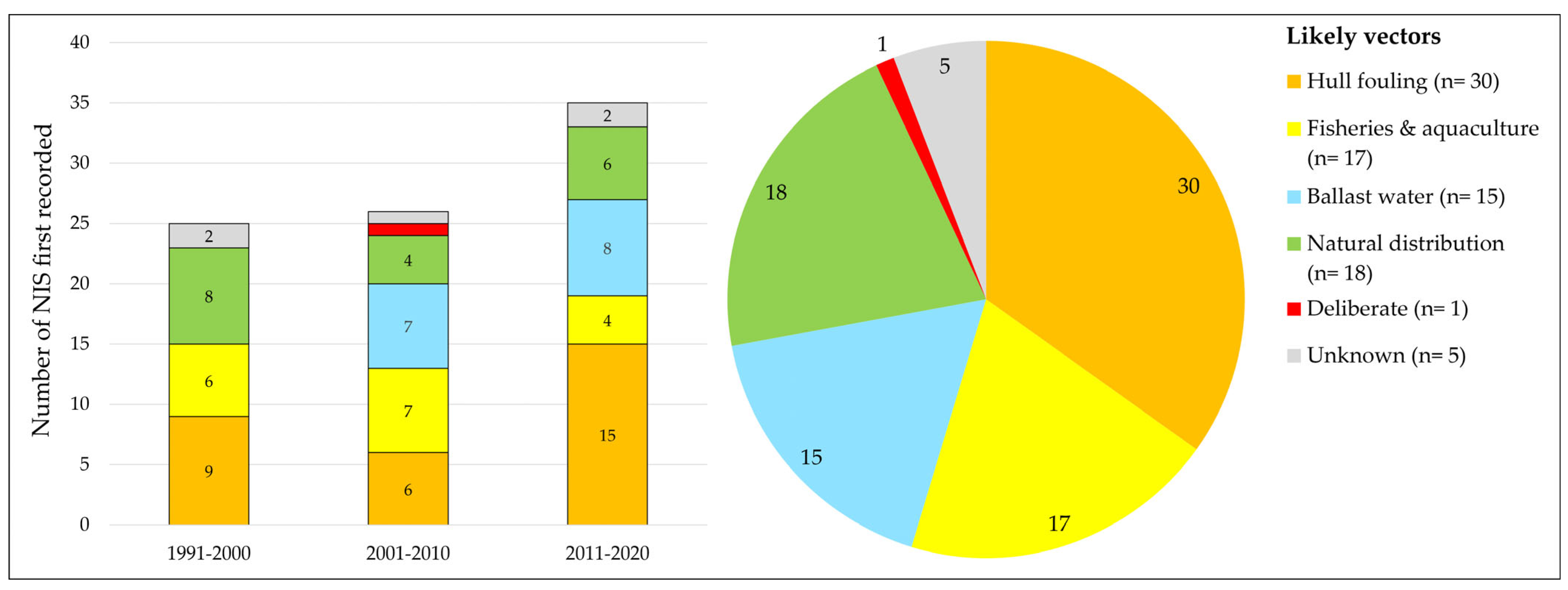
3.1.4. Likely Vectors with Which NIS Were First Introduced
- During the last three decades, i.e., between 1991 and 2020, but also before that time [34], hull fouling has generally been assumed to be the main vector connected to the introduction of marine NIS into the coastal waters of The Netherlands.
- After hull fouling, ballast water, fisheries, and aquaculture (mainly shellfish transports) have most frequently been linked to introductions, as well as the “natural distribution” of NIS that were introduced elsewhere in NW Europe and secondarily reached the Dutch coast by natural dispersal.
- Over time, not all vectors were assumed to play an equally important role concerning the introduction of species. Ballast water, for example, is assumed to be responsible for the introduction of 15 NIS in 2001 to 2020, while none of the NIS first recorded in 1991–2000 were assumed to have arrived by this vector.
- The number of NIS introduced by hull fouling appears to have almost tripled in 2011–2020 in comparison to 2001–2010, while the number of species assumed to be introduced by fisheries and aquaculture decreased slightly.
3.2. Unnoticed Establishments and Unexpected Pathways
3.2.1. Overlooked Establishment of the Cryptic Moon Jellyfish Aurelia coerulea
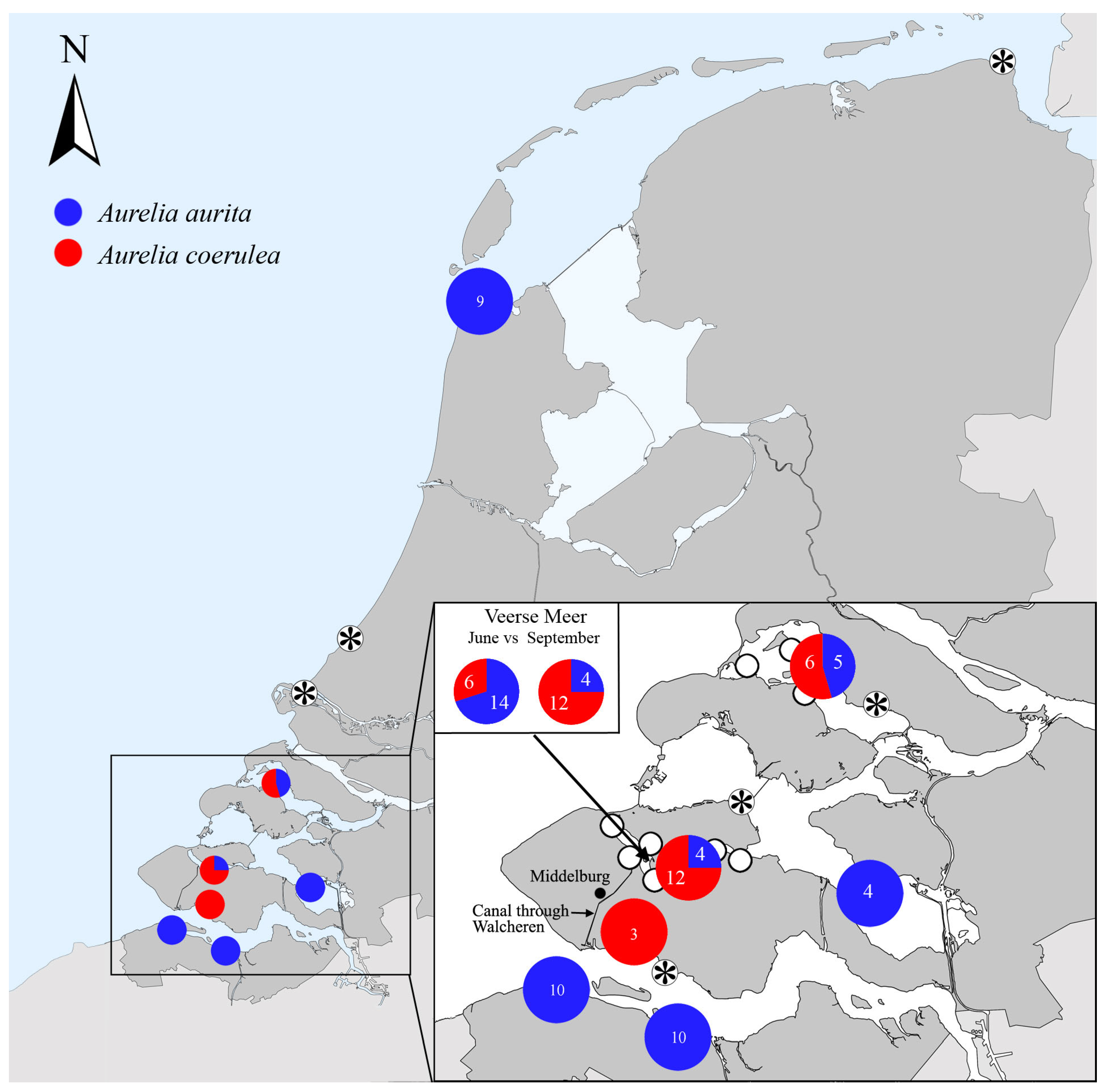
- Aurelia coerulea is established in a panmictic population together with A. aurita in the inland marine lakes Veerse Meer and Grevelingen (Figure 11). Moon jellyfish collected at more exposed sites along the North Sea and within the Oosterschelde and southern Westerschelde represented A. aurita. The establishment of A. coerulea within the sheltered inland marine lakes of The Netherlands agrees with the fact that this species has a lagoonal- or harbour-limited distribution within the Mediterranean [33]. The presence of A. coerulea in the northern Westerschelde, in the port of Vlissingen (Sloehaven), also supports this and may be linked to the canal through Walcheren, which connects the Veerse Meer with the Westerschelde, near the Sloehaven. The high numbers of moon jellyfish sighted in the canals in Middelburg in 2020, mentioned earlier, may have concerned this jellyfish species, as well.
- 2.
- At least within the Veerse Meer, Aurelia aurita is the dominant species in early summer, while A. coerulea is dominant in late summer. Thus, the two species peak after each other in early to late summer. As a result, high numbers of moon jellyfish (Aurelia spp.) can be encountered in the lake throughout the summer.
- 3.
- Collecting and sequencing Aurelia polyps from settlement plates in the summer of 2021 deployed at a depth of 1 m in the Veerse Meer resulted in the detection of only A. aurita. Specifically aimed at detecting the whereabouts of A. coerulea polyps in the lake, on 29 October 2022, several medusae were collected from a beach off Geersdijk, where polyps were collected from oyster shells the same day by scuba diving between 8 and 10 m in depth. Subsequent DNA analyses of four medusae and six polyps taken from different shells indicated that they all were A. coerulea (Genbank nrs: OQ940545-OQ940549). Detecting the polyps was not assumed to be simple, taking into account that only the polyps of A. aurita were detected in the Veerse Meer in 2021, whereas a recent study by van Walraven et al. [38], specifically focused on finding the polyps of all jellyfish species occurring in the southern North Sea area, also resulted in only the detection of A. aurita.
- -
- It suddenly occurs outside of its “traditional” habitat;
- -
- It suddenly occurs in different environmental conditions, such as temperature or salinity range;
- -
- Its seasonal patterns suddenly change.
3.2.2. Two Pacific Flatworms Introduced to the NE Atlantic
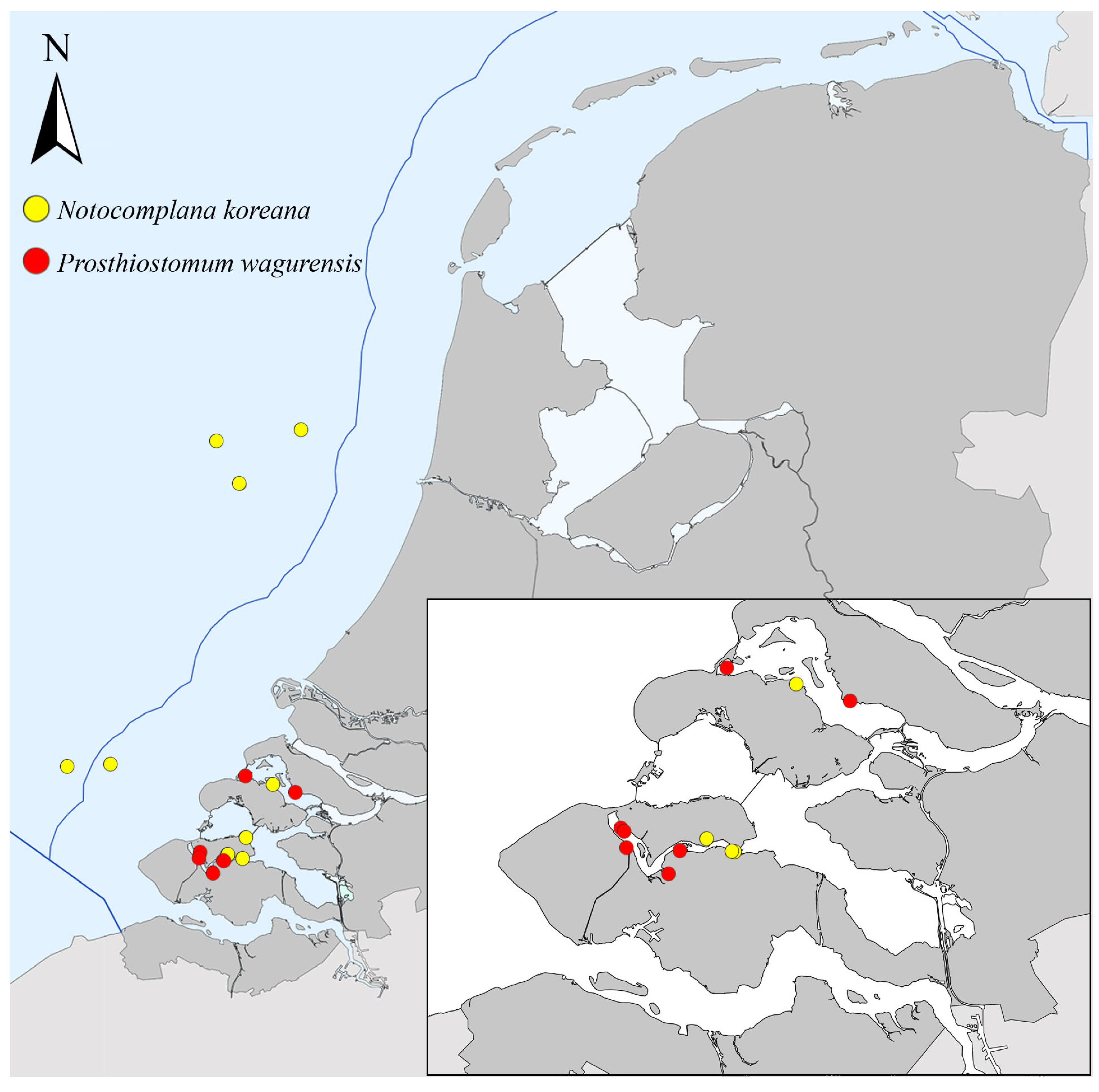
- Prosthiostomum wagurensis and Notocomplana koreana both originate from the Pacific and were both first identified based on samples collected in the Veerse Meer. Although one may expect a similar introduction pathway, their distribution outside of this inland seawater lake does not support this view.
- The presence of P. wagurensis could only be validated based on DNA analyses in sheltered inland seawater systems, such as the Veerse Meer and the Grevelingen (Figure 1). Taking into account that flatworms not distinguishable from P. wagurensis in terms of external features had already been noticed during earlier unpublished work in and near the ports of Rotterdam (Netherlands) on 22 May 2019, Vlissingen (Netherlands) on 27 May 2019, and Zeebrugge (Belgium) on 2 October 2019 (Faasse, unpublished), it appears most likely that the species was introduced into these ports and inland waters by ship hull fouling, possibly using the more sheltered parts of ports in Europe as stepping stones. Where in Europe the species was first introduced remains uncertain.
- The presence of N. koreana was validated based on DNA analyses in the same sheltered inland seawater systems as P. wagurensis, but various additional specimens were collected by scrape sampling from 5 out of the 13 navigational buoys on the open North Sea that were searched for NIS in 2021. This indicates that this species can establish itself well in highly exposed habitats, and that it may be using the navigational buoys on the open sea off the west coast of Europe, in addition to ports, as stepping stones. Its potential presence in ports was already indicated by a study based on samples taken in 2017 in the port of Vlissingen (Sloehaven), in the Westerschelde (Figure 1). Although no flatworms were noticed at that time, metabarcoding indicated the presence of the DNA (COI) of N. koreana in a bulk sample taken from a settlement plate [56].
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.B.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Cinar, M.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Marbuah, G.; Gren, I.-M.; McKie, B. Economics of Harmful Invasive Species: A Review. Diversity 2014, 6, 500–523. [Google Scholar] [CrossRef]
- Wainright, C.A.; Muhlfeld, C.C.; Elser, J.J.; Bourret, S.L.; Devlin, S.P. Species invasion progressively disrupts the trophic structure of native food webs. Proc. Natl. Acad. Sci. USA 2021, 118, e2102179118. [Google Scholar] [CrossRef] [PubMed]
- EU. Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. Available online: https://eur-lex.europa.eu/legalcontent/EN/TXT/?uri=celex%3A32014R1143 (accessed on 9 April 2023).
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- EU. Marine Strategy Framework Directive 2008/56/EC. In Official Journal of the European Union; EU: Maastricht, The Netherlands, 2008; pp. 19–40. [Google Scholar]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning-Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Massé, C.; Viard, F.; Humbert, S.; Antajan, E.; Auby, I.; Bachelet, G.; Bernard, G.; Bouchet, V.M.P.; Burel, T.; Dauvin, J.-C.; et al. An Overview of Marine Non-Indigenous Species Found in Three Contrasting Biogeographic Metropolitan French Regions: Insights on Distribution, Origins and Pathways of Introduction. Diversity 2023, 15, 161. [Google Scholar] [CrossRef]
- Jensen, K.R.; Andersen, P.; Andersen, N.R.; Bruhn, A.; Buur, H.; Carl, H.; Jakobsen, H.; Jaspers, C.; Lundgreen, K.; Nielsen, R.; et al. Reviewing Introduction Histories, Pathways, Invasiveness, and Impact of Non-Indigenous Species in Danish MarineWaters. Diversity 2023, 15, 434. [Google Scholar] [CrossRef]
- Gittenberger, A.; WG-AS. Trilateral Wadden Sea Management and Action Plan for Alien Species (MAPAS); Busch, J.A., Lüerßen, G., de Jong, F., Eds.; Common Wadden Sea Secretariat (CWSS): Wilhelmshaven, Germany, 2019; 45p. [Google Scholar]
- EU. Habitat Directive according to Council Directive 92/43/EEC of 21 May 1992. Available online: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm (accessed on 9 April 2023).
- Bleker, H. Beleidsregels van de Staatssecretaris van Economische Zaken, Landbouw en Innovatie van 6 juni 2012, nr. 267278, houdende vaststelling van beleidsregels inzake schelpdierverplaatsingen. Staatscourant 2012, 12068, 4p. [Google Scholar]
- Gittenberger, A.; Wesdorp, K.H.; Rensing, M. Biofouling as a Transport Vector of Non-Native Marine Species in the Dutch Delta, along the North Sea Coast and in the Wadden Sea. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2017; GiMaRIS Rapport 2017_03; 48p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Stegenga, H.; Hoeksema, B.W. Native and non-native species of hard substrata in the Dutch Wadden Sea. Ned. Faun. Meded. 2010, 33, 21–75. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Schrieken, N.; Stegenga, H. Waddenzee Inventarisatie van aan Hard Substraat Gerelateerde Organismen Met de Focus Op Exoten, Zomer 2011. Issued by Producentenorganisatie van de Nederlandse Mosselcultuur; GiMaRIS: Sassenheim, The Netherlands, 2012; GiMaRIS Rapport 2012_01; 61p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Dekker, R.; Niemantsverdriet, P.; Schrieken, N.; Stegenga, H. Native and Non-Native Species of the Dutch Wadden Sea in 2014. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2015; GiMaRIS Rapport 2015_08; 93p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; van der Veer, H.W.; Philippart, C.J.M.; van der Hoorn, B.; D’Hont, A.; Wesdorp, K.H.; Schrieken, N.; Klunder, L.; Kleine-Schaars, L.; et al. Native and Non-Native Species of the Dutch Wadden Sea in 2018. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2019; GiMaRIS Rapport 2019_09; 124p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Keeler-Perez, H.; Bijleveld, A.; Coppis, S. Native and non-native species of the Dutch Wadden Sea in 2022. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, in preparation.
- Gittenberger, A.; Rensing, M.; Niemantsverdriet, P.; Schrieken, N.; D’Hont, A.; Stegenga, H. Soorteninventarisatie Oesterputten en Oesterpercelen, 2015. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2015; GiMaRIS Rapport 2015_09; 23p. [Google Scholar]
- Wijnhoven, S.; Gittenberger, A.; Faasse, M.; Schellekens, T. Overview Alien Species Monitoring in the Western Scheldt: Current Status of Monitoring Efforts and Presence of Alien Species among Macrofauna and Algae; Ecoauthor: Heinkenszand, The Netherlands, 2017; Ecoauthor Report 2017-01; 56p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Faasse, M.A.; Keeler Perez, H.; Gittenberger, E. Native and Non-Native Species of the Veerse Meer, 2020–2021. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2021; GiMaRIS Rapport 2021_07; 153p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Keeler Perez, H. Marine Alien Species Detection Network of The Netherlands: 2021 Results. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2022; GiMaRIS Rapport 2022_15; 46p. [Google Scholar]
- Slijkerman, D.M.E.; Glorius, S.T.; Gittenberger, A.; Weide, B.E.v.d.; Bos, O.G.; Rensing, M.; Groot, G.A.d. Monitoring Groningen Sea Ports, Non-Indigenous Species and Risks from Ballast Water in Eemshaven and Delfzijl; Wageningen Marine Research (University & Research Centre): Wageningen, The Netherlands, 2017; Marine Research Report C045/17A; 81p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Niemantsverdriet, P.; Schrieken, N.; Stegenga, H. Port of Rotterdam Survey and Monitoring Non-Native Species Conform HELCOM/OSPAR Protocol. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2014; GiMaRIS Rapport 2014_31; 111p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Schrieken, N.; Stegenga, H. Port of Vlissingen Survey and Monitoring Non-Native Species in 2016 Conform HELCOM/OSPAR Protocol. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2017; GiMaRIS Rapport 2017_05; 49p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Stegenga, H. Non-Native Species Survey 2014 in the Military Harbour of Den Helder. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2015; GiMaRIS Rapport 2015_03; 10p. [Google Scholar]
- Gittenberger, A.; Rensing, M.; Keeler-Perez, H. Non-Native Species Focused Assessment in the Port of IJmuiden in 2022. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; GiMaRIS: Sassenheim, The Netherlands, 2022; GiMaRIS Rapport 2022_52; 32p. [Google Scholar]
- HELCOM/OSPAR. Joint Harmonised Procedure for the Contracting Parties of HELCOM and OSPAR on the Granting of Exemptions under the International Convention for the Control and Management of Ship’s Ballast Water and Sediments, Regulation A-4; Adopted as OSPAR Agreement 2013-09 and by HELCOM Ministerial Meeting Copenhagen 3 October 2013; OSAR Commission: Edinburg, TX, USA, 2013; 49p. [Google Scholar]
- Loos, L.M.v.d.; Gmelig Meyling, A.W. Het Duiken Gebruiken 4. Gegevensanalyse van Het Monitoringproject Onderwater Oever (MOO). Fauna-Onderzoek Met Sportduikers in Oosterschelde en Grevelingenmeer. Periode 1994 t/m 2018; Stichting ANEMOON: Bennebroek, The Netherlands, 2019; 85p. [Google Scholar]
- Didderen, K.; Have, T.M.v.d.; Dorenbosch, M. Analysis of the Potential to Detect Alien Species in Marine MWTL Monitoring. Review of Efficiency and Possible Improvement. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; Bureau Waardenburg: Culemborg, The Netherlands, 2015; Report nr 15-014; 94p. [Google Scholar]
- Scorrano, S.; Aglieri, G.; Boero, F.; Dawson, M.N.; Piraino, S. Unmasking Aurelia species in the Mediterranean Sea: An integrative morphometric and molecular approach. Zool. J. Linn. Soc. 2016, 180, 243–267. [Google Scholar] [CrossRef]
- Wolff, W.J. Non-indigenous marine and estuarine species in The Netherlands. Zool. Med. Leiden 2005, 79, 1–116. [Google Scholar]
- Capa, M.; van Moorsel, G.; Tempelman, D. The Australian feather-duster worm Laonome calida Capa, 2007 (Annelida: Sabellidae) introduced into European inland waters? BioInvasions Rec. 2014, 3, 1–11. [Google Scholar] [CrossRef]
- Lawley, J.W.; Gamero-Mora, E.; Maronna, M.M.; Chiaverano, L.M.; Stampar, S.N.; Hopcroft, R.R.; Collins, A.G.; Morandini, A.C. The importance of molecular characters when morphological variability hinders diagnosability: Systematics of the moon jellyfish genus Aurelia (Cnidaria: Scyphozoa). PeerJ 2021, 9, e11954. [Google Scholar] [CrossRef] [PubMed]
- Pitt, K.A.; Cathy, L.H.; Condon, R.H.; Duarte, C.M.; Stewart-Koster, B. Claims That Anthropogenic Stressors Facilitate Jellyfish Blooms Have Been Amplified Beyond the Available Evidence: A Systematic Review. Front. Mar. Sci. 2018, 5, 451. [Google Scholar] [CrossRef]
- van Walraven, L.; Driessen, F.; van Bleijswijk, J.; Bol, A.; Luttikhuizen, P.C.; Coolen, J.W.P.; Bos, O.G.; Gittenberger, A.; Schrieken, N.; Langenberg, V.T.; et al. Where are the polyps? Molecular identification, distribution and population differentiation of Aurelia aurita. Mar. Biol. 2016, 163, 172. [Google Scholar] [CrossRef]
- Gittenberger, A.; Gittenberger, E. Cryptic, adaptive radiation of endoparasitic snails: Sibling species in Leptoconchus (Gastropoda: Coralliophilidae) in corals. Org. Divers. Evol. 2011, 11, 21–41. [Google Scholar] [CrossRef]
- Dawson, M.N. Incipient speciation of Catostylus mosaicus (Scyphozoa, Rhizostomeae, Catostylidae), comparative phylogeography and biogeography in south-east Australia. J. Biogeogr. 2005, 32, 515–533. [Google Scholar] [CrossRef]
- EU. Regulation (EU) No 1100/2007 of the European Parliament and of the Council of 18 September 2007 on Establishing Measures for the Recovery of the Stock of European eel. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX:32007R1100 (accessed on 9 April 2023).
- Arai, M.N. A Functional Biology of Scyphozoa; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Suzuki, K.S.; Suzuki, K.W.; Kumakura, E.; Sato, K.; Oe, Y.; Sato, K.; Sawada, H.; Masuda, R.; Nogata, Y. Seasonal Alternation of the Ontogenetic Development of the Moon Jellyfish Aurelia coerulea in Maizuru Bay, Japan. PLoS ONE 2019, 14, 11. [Google Scholar] [CrossRef]
- Purcell, J.E.; Uye, S.; Lo, W.T. Anthropogenic Causes of Jellyfish Blooms and Their Direct Consequences for Humans: A Review. Mar. Ecol. Prog. 2007, 350, 153–174. [Google Scholar] [CrossRef]
- Lucas, C.H.; Gelcich, S.; Uye, S.-I. Living with Jellyfish: Management and Adaptation Strategies’. In Jellyfish Blooms; Pitt, K.A., Lucas, C.H., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 129–150. [Google Scholar] [CrossRef]
- Yoon, W.; Chae, J.; Koh, B.-S.; Han, C. Polyp Removal of a Bloom Forming Jellyfish, Aurelia coerulea, in Korean Waters and Its Value Evaluation’. Ocean Sci. J. 2018, 53, 499–507. [Google Scholar] [CrossRef]
- van Walraven, L.; Couperus, B. Kwalproblemen Sloecentrale: Eerste Fase Verkennend Onderzoek; Wageningen Marine Research: Wageningen, The Netherlands, 2023; Rapport C007/23; 63p. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological invasions and cryptogenic species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Folino-Rorem, N.C.; Darling, J.A.; D’Ausilio, C.A. Genetic analysis reveals multiple cryptic invasive species of the hydrozoan genus Cordylophora. Biol. Invasions 2009, 11, 1869–1882. [Google Scholar] [CrossRef]
- Styan, C.A.; McCluskey, C.F.; Sun, Y.; Kupriyanova, E.K. Cryptic sympatric species across the Australian range of the global estuarine invader Ficopomatus enigmaticus (Fauvel, 1923) (Serpulidae, Annelida). Aquat. Invasions 2017, 12, 53–65. [Google Scholar] [CrossRef]
- Faubel, A.; Gollasch, S. Cryptostylochus hullensis sp. nov. (Polycladida, Acotylea, Platyhelminthes): Apossible case of transoceanic dispersal on a ship’s hull. Helgol. Meeresunters. 1996, 50, 533–537. [Google Scholar] [CrossRef]
- Hofker, J. Faunistische Beobachtungen in der Zuidersee während der Trockenlegung. Z. Morphol. Ökol. Tiere 1930, 18, 189–216. [Google Scholar] [CrossRef]
- Sluys, R.; Faubel, A.; Rajagopal, S.; Velde, G. van der. A new and alien species of “oyster leech” (Platyhelminthes, Polycladida, Stylochidae) from the brackish North Sea Canal, The Netherlands. Helgol. Mar. Res. 2005, 59, 310–314. [Google Scholar] [CrossRef]
- Faasse, M.; Ates, R. De Nederlandse polyclade platwormen (Platyhelminthes: Turbellaria: Polycladida). 2. De uit Amerika afkomstige Euplana gracilis (Girard, 1850). Het. Zeepaard. 2003, 63, 57–60. [Google Scholar]
- Tsunashima, T.; Hagiya, M.; Yamada, R.; Koito, T.; Tsuyuki, N.; Izawa, S.; Kosoba, K.; Itoi, K.; Sugita, H. A molecular framework for the taxonomy and systematics of Japanese marine turbellarian flatworms (Platyhelminthes, Polycladida). Aquat. Biol. 2017, 26, 159–167. [Google Scholar] [CrossRef]
- van der Hoorn, B.; Gittenberger, A. Environmental DNA Sloehaven; A Multi-Substrate Metabarcoding Approach for Detecting Non-Indigenous Species in a Dutch Port. Issued by Office for Risk Assessment and Research, The Netherlands Food and Consumer Product Safety Authority; Naturalis Biodiversity Center: Leiden, The Netherlands, 2019; Naturalis Report 2018_01; 43p. [Google Scholar]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gittenberger, A.; Rensing, M.; Faasse, M.; van Walraven, L.; Smolders, S.; Keeler Perez, H.; Gittenberger, E. Non-Indigenous Species Dynamics in Time and Space within the Coastal Waters of The Netherlands. Diversity 2023, 15, 719. https://doi.org/10.3390/d15060719
Gittenberger A, Rensing M, Faasse M, van Walraven L, Smolders S, Keeler Perez H, Gittenberger E. Non-Indigenous Species Dynamics in Time and Space within the Coastal Waters of The Netherlands. Diversity. 2023; 15(6):719. https://doi.org/10.3390/d15060719
Chicago/Turabian StyleGittenberger, Arjan, Marjolein Rensing, Marco Faasse, Lodewijk van Walraven, Sander Smolders, Helena Keeler Perez, and Edmund Gittenberger. 2023. "Non-Indigenous Species Dynamics in Time and Space within the Coastal Waters of The Netherlands" Diversity 15, no. 6: 719. https://doi.org/10.3390/d15060719
APA StyleGittenberger, A., Rensing, M., Faasse, M., van Walraven, L., Smolders, S., Keeler Perez, H., & Gittenberger, E. (2023). Non-Indigenous Species Dynamics in Time and Space within the Coastal Waters of The Netherlands. Diversity, 15(6), 719. https://doi.org/10.3390/d15060719