
A Cocktail of Plankton and Organochlorines for Whale Shark in the Foraging Areas of Nosy Be (Madagascar)

 ,
, 

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
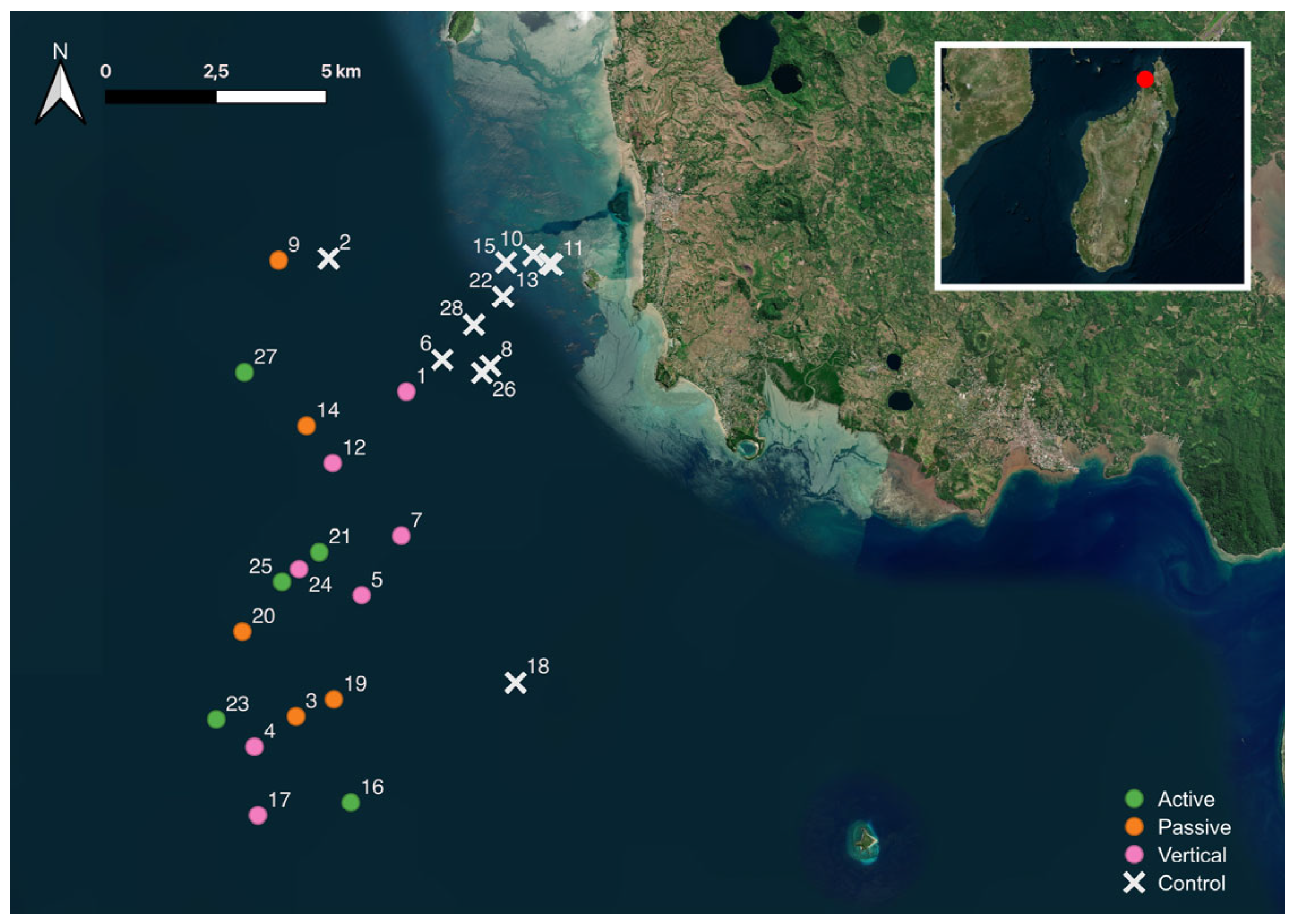
2.1. Study Area
2.2. Samples Collection
2.3. Sample Preparation and OC Determination
2.4. Data Analysis
3. Results and Discussion
3.1. POP Concentrations in Plankton Samples
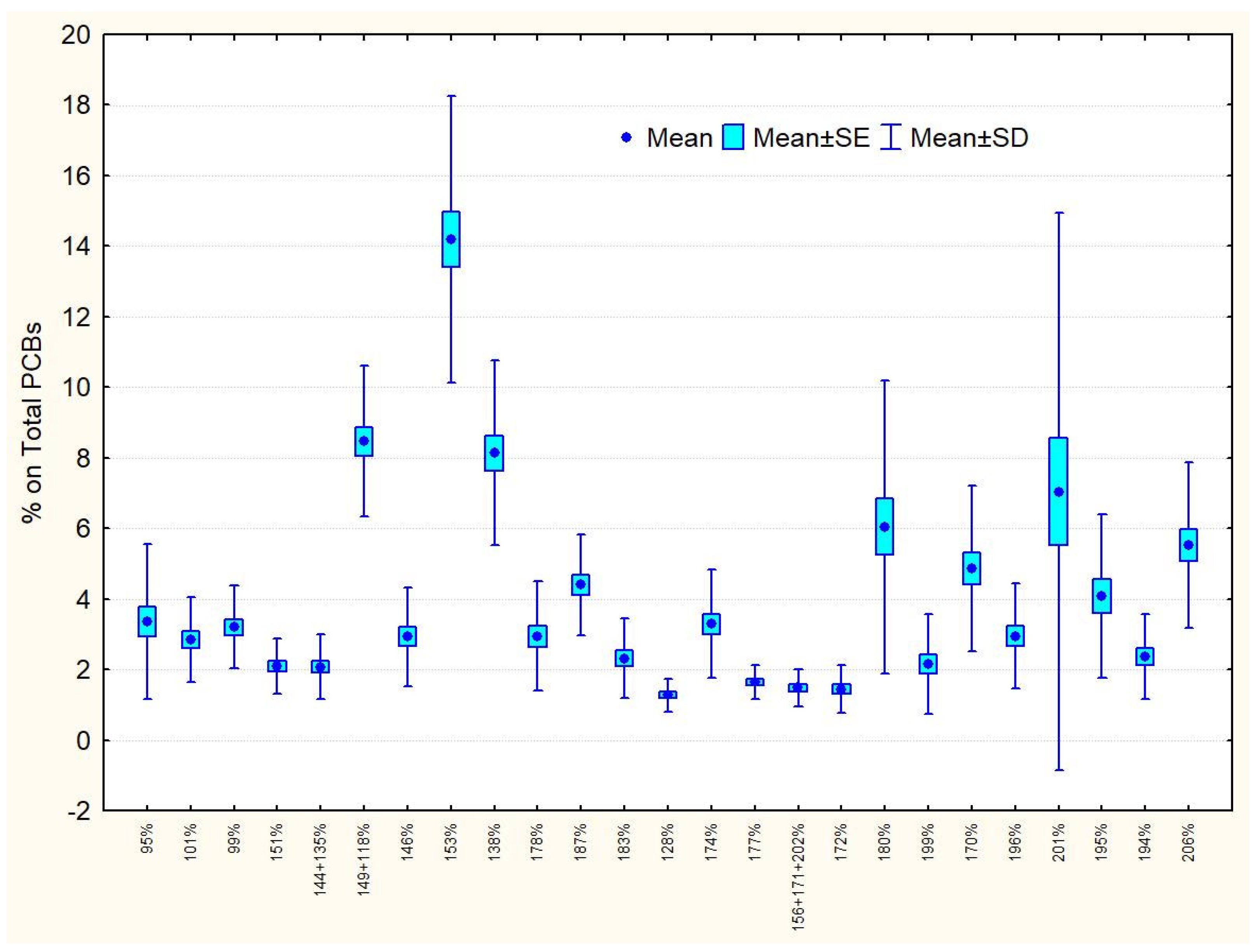
3.2. PCB Congeners Composition
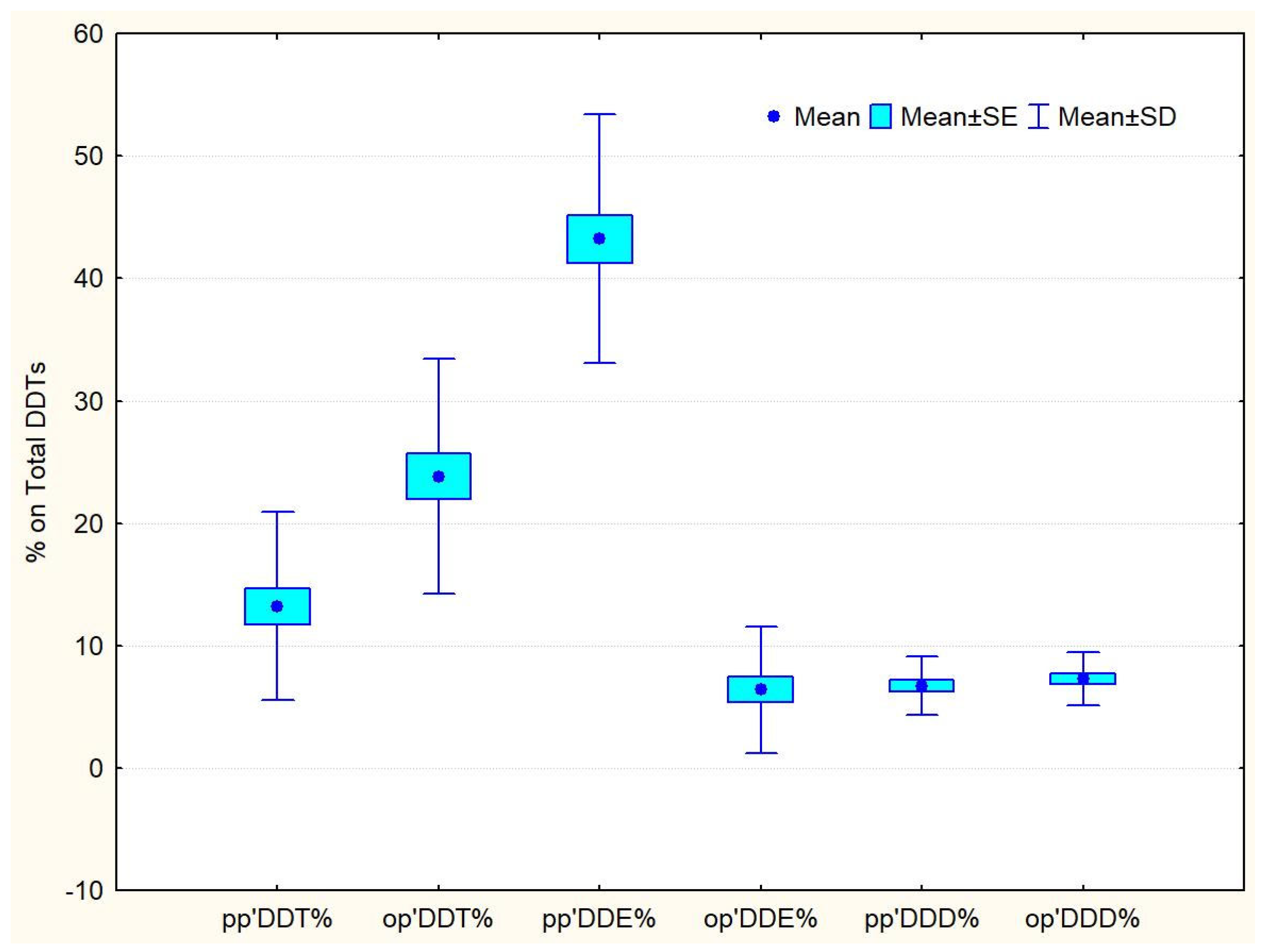
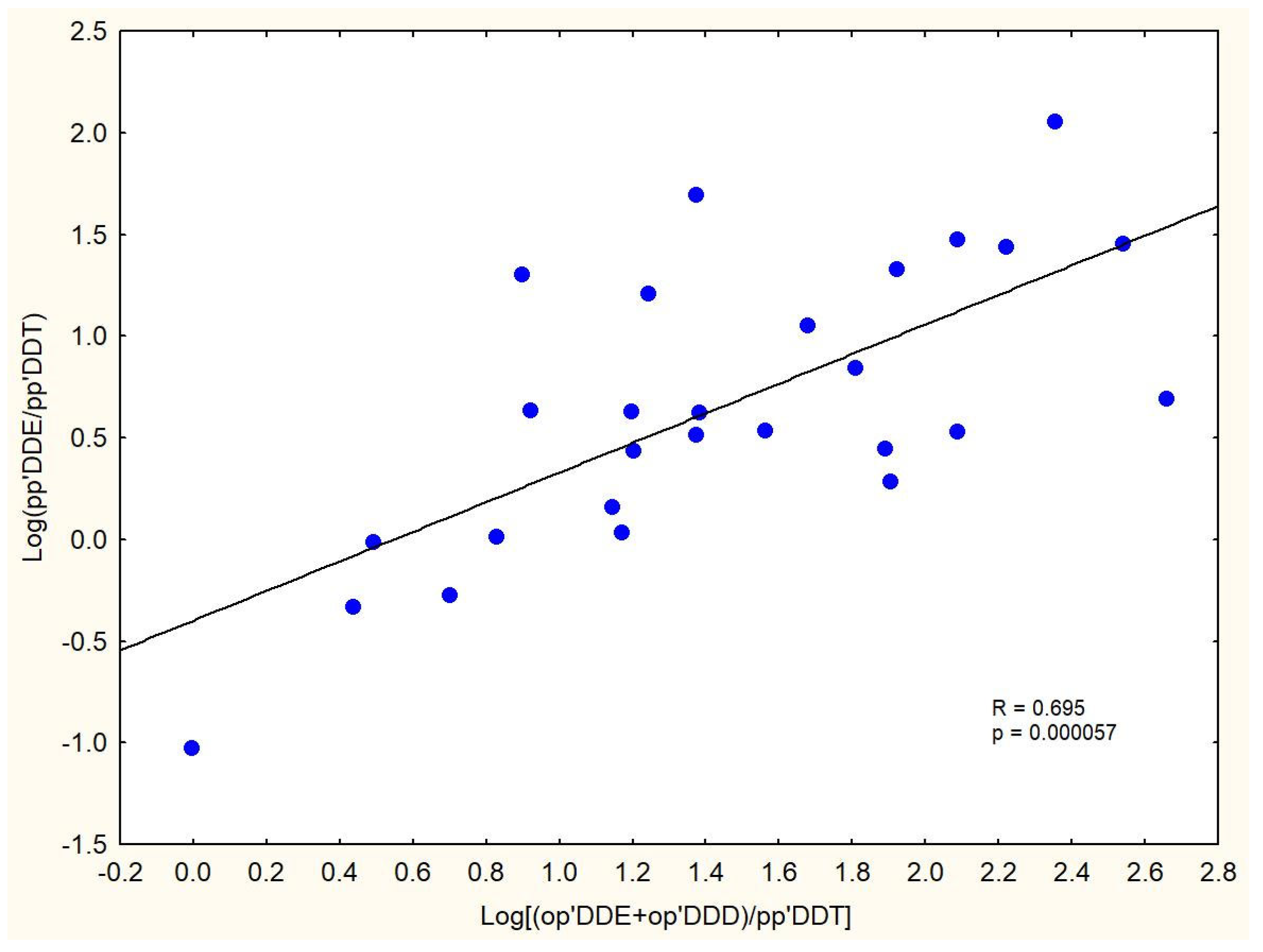
3.3. DDT Isomer Composition and Ratios
3.4. Potential POPs Uptake in the Whale Shark
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bettinetti, R.; Manca, M. Understanding the Role of Zooplankton in Transfer of Pollutants through Trophic Food Webs. In Zooplankton: Species Diversity, Distribution and Seasonal Dynamics; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2013; p. 18. [Google Scholar]
- Boldrocchi, G.; Moussa Omar, Y.; Rowat, D.; Bettinetti, R. First Results on Zooplankton Community Composition and Contamination by Some Persistent Organic Pollutants in the Gulf of Tadjoura (Djibouti). Sci. Total Environ. 2018, 627, 812–821. [Google Scholar] [CrossRef]
- Nybakken, J.W. Marine Biology: An Ecological Approach, 5th ed.; Benjamin Cummings: San Francisco, CA, USA, 2001; ISBN 9780321030764. [Google Scholar]
- Compagno, L.J.V. Bullhead, Mackerel and Carpet Sharks: Heterodontiformes, Lamniformes and Orectolobiformes. In Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; ISBN 9789251045435. [Google Scholar]
- Norman, B. Rhincodon typus. In The IUCN Red List of Threatened Species 2000; IUCN: Gland, Switzerland, 2000. [Google Scholar]
- Norman, B. Rhincodon typus. In The IUCN Red List of Threatened Species 2005; IUCN: Gland, Switzerland, 2005. [Google Scholar]
- Pierce, S.J.; Norman, B. Rhincodon typus. In The IUCN Red List of Threatened Species 2016; IUCN: Gland, Switzerland, 2016. [Google Scholar]
- Colman, J.G. A Review of the Biology and Ecology of the Whale Shark. J. Fish Biol. 1997, 51, 1219–1234. [Google Scholar] [CrossRef]
- Motta, P.J.; Maslanka, M.; Hueter, R.E.; Davis, R.L.; de la Parra, R.; Mulvany, S.L.; Habegger, M.L.; Strother, J.A.; Mara, K.R.; Gardiner, J.M.; et al. Feeding Anatomy, Filter-Feeding Rate, and Diet of Whale Sharks Rhincodon Typus during Surface Ram Filter Feeding off the Yucatan Peninsula, Mexico. Zoology 2010, 113, 199–212. [Google Scholar] [CrossRef]
- Hacohen-Domené, A.; Galván-Magaña, F.; Ketchum-Mejia, J. Abundance of Whale Shark (Rhincodon Typus) Preferred Prey Species in the Southern Gulf of California, Mexico. Cybium 2006, 30, 99–102. [Google Scholar]
- Reyes-Mendoza, O.; Cárdenas-Palomo, N.; Herrera-Silveira, J.; Mimila-Herrera, E.; Trujillo-Córdova, J.; Chiappa-Carrara, X.; Arceo-Carranza, D. Quantity and Quality of Prey Available for the Whale Shark (Rhincodon typus, Smith 1828) at the Mexican Caribbean Aggregation Site. Reg. Stud. Mar. Sci. 2021, 43, 101696. [Google Scholar] [CrossRef]
- Bava, P.; Micarelli, P.; Buttino, I. Zooplankton Assemblage Diversity in the Whale Shark Rhincodon Typus Aggregation Area of Nosy Be (Madagascar). Estuar. Coast. Shelf Sci. 2022, 279, 108159. [Google Scholar] [CrossRef]
- Petatán-Ramírez, D.; Whitehead, D.A.; Guerrero-Izquierdo, T.; Ojeda-Ruiz, M.A.; Becerril-García, E.E. Habitat Suitability of Rhincodon Typus in Three Localities of the Gulf of California: Environmental Drivers of Seasonal Aggregations. J. Fish Biol. 2020, 97, 1177–1186. [Google Scholar] [CrossRef]
- Taylor, L.R.; Compagno, L.J.V.; Struhsaker, P.J. Megamouth—A New Species, Genus, and Family of Lamnoid Shark (Megachasma pelagios, Family Megachasmidae) from the Hawaiian Islands. Proc. Calif. Acad. Sci. 1983, 43, 87–110. [Google Scholar]
- Rowat, D.; Brooks, K.; March, A.; McCarten, C.; Jouannet, D.; Riley, L.; Jeffreys, G.; Perri, M.; Vely, M.; Pardigon, B.; et al. Long-Term Membership of Whale Sharks (Rhincodon typus) in Coastal Aggregations in Seychelles and Djibouti. Mar. Freshw. Res. 2011, 62, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Sequeira, A.M.M.; Mellin, C.; Fordham, D.A.; Meekan, M.G.; Bradshaw, C.J.A. Predicting Current and Future Global Distributions of Whale Sharks. Glob. Change Biol. 2014, 20, 778–789. [Google Scholar] [CrossRef]
- Castro, A.L.F.; Stewart, B.S.; Wilson, S.G.; Hueter, R.E.; Meekan, M.G.; Motta, P.J.; Bowen, B.W.; Karl, S.A. Population Genetic Structure of Earth’s Largest Fish, the Whale Shark (Rhincodon typus). Mol. Ecol. 2007, 16, 5183–5192. [Google Scholar] [CrossRef]
- Pierce, S.J.; Méndez-Jiménez, A.; Collins, K.; Rosero-Caicedo, M.; Monadjem, A. Developing a Code of Conduct for Whale Shark Interactions in Mozambique. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 782–788. [Google Scholar] [CrossRef]
- Berumen, M.L.; Braun, C.D.; Cochran, J.E.M.; Skomal, G.B.; Thorrold, S.R. Movement Patterns of Juvenile Whale Sharks Tagged at an Aggregation Site in the Red Sea. PLoS ONE 2014, 9, e103536. [Google Scholar] [CrossRef] [Green Version]
- Fossi, M.C.; Baini, M.; Panti, C.; Galli, M.; Jiménez, B.; Muñoz-Arnanz, J.; Marsili, L.; Finoia, M.G.; Ramírez-Macías, D. Are Whale Sharks Exposed to Persistent Organic Pollutants and Plastic Pollution in the Gulf of California (Mexico)? First Ecotoxicological Investigation Using Skin Biopsies. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 199, 48–58. [Google Scholar] [CrossRef]
- Heyman, W.D.; Graham, R.T.; Kjerfve, B.; Johannes, R.E. Whale Sharks Rhincodon Typus Aggregate to Feed on Fish Spawn in Belize. Mar. Ecol. Prog. Ser. 2001, 215, 275–282. [Google Scholar] [CrossRef]
- Meekan, M.G.; Jarman, S.N.; McLean, C.; Schultz, M.B.; Meekan, M.G.; Jarman, S.N.; McLean, C.; Schultz, M.B. DNA Evidence of Whale Sharks (Rhincodon typus) Feeding on Red Crab (Gecarcoidea natalis) Larvae at Christmas Island, Australia. Mar. Freshw. Res. 2009, 60, 607–609. [Google Scholar] [CrossRef]
- Meekan, M.G.; Bradshaw, C.J.A.; Press, M.; McLean, C.; Richards, A.; Quasnichka, S.; Taylor, J.G. Population Size and Structure of Whale Sharks Rhincodon typus at Ningaloo Reef, Western Australia. Mar. Ecol. Prog. Ser. 2006, 319, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Boldrocchi, G.; Monticelli, D.; Butti, L.; Omar, M.; Bettinetti, R. First Concurrent Assessment of Elemental- and Organic-Contaminant Loads in Skin Biopsies of Whale Sharks from Djibouti. Sci. Total Environ. 2020, 722, 137841. [Google Scholar] [CrossRef]
- Davis, D.; Banks, S.; Birtles, A.; Valentine, P.; Cuthill, M. Whale Sharks in Ningaloo Marine Park: Managing Tourism in an Australian Marine Protected Area. Tour. Manag. 1997, 18, 259–271. [Google Scholar] [CrossRef]
- Graham, R.; Irvine, T.R.; Keesing, J.K. Iterative Planning and Adaptive Management of Whale Shark Tourism in Belize: Global Implications of Lessons Learnt from 1998 and 2004. In CSIRO Marine and Atmospheric Research; CSIRO: Canberra, Australia, 2007; pp. 73–74. [Google Scholar]
- Quiros, A.L. Tourist Compliance to a Code of Conduct and the Resulting Effects on Whale Shark (Rhincodon typus) Behavior in Donsol, Philippines. Fish. Res. 2007, 84, 102–108. [Google Scholar] [CrossRef]
- Rowat, D.; Meekan, M.G.; Engelhardt, U.; Pardigon, B.; Vely, M. Aggregations of Juvenile Whale Sharks (Rhincodon typus) in the Gulf of Tadjoura, Djibouti. Environ. Biol. Fishes 2007, 80, 465–472. [Google Scholar] [CrossRef]
- Fossi, M.C.; Coppola, D.; Baini, M.; Giannetti, M.; Guerranti, C.; Marsili, L.; Panti, C.; de Sabata, E.; Clò, S. Large Filter Feeding Marine Organisms as Indicators of Microplastic in the Pelagic Environment: The Case Studies of the Mediterranean Basking Shark (Cetorhinus maximus) and Fin Whale (Balaenoptera physalus). Mar. Environ. Res. 2014, 100, 17–24. [Google Scholar] [CrossRef]
- Tiktak, G.P.; Butcher, D.; Lawrence, P.J.; Norrey, J.; Bradley, L.; Shaw, K.; Preziosi, R.; Megson, D. Are Concentrations of Pollutants in Sharks, Rays and Skates (Elasmobranchii) a Cause for Concern? A Systematic Review. Mar. Pollut. Bull. 2020, 160, 111701. [Google Scholar] [CrossRef]
- Consales, G.; Marsili, L. Assessment of the Conservation Status of Chondrichthyans: Underestimation of the Pollution Threat. Eur. Zool. J. 2021, 87, 165–180. [Google Scholar] [CrossRef]
- Chiuchiolo, A.L.; Dickhut, R.M.; Cochran, M.A.; Ducklow, H.W. Persistent Organic Pollutants at the Base of the Antarctic Marine Food Web. Environ. Sci. Technol. 2004, 38, 3551–3557. [Google Scholar] [CrossRef] [Green Version]
- Castro-Jiménez, J.; Bănaru, D.; Chen, C.-T.; Jiménez, B.; Muñoz-Arnanz, J.; Deviller, G.; Sempéré, R. Persistent Organic Pollutants Burden, Trophic Magnification and Risk in a Pelagic Food Web from Coastal NW Mediterranean Sea. Environ. Sci. Technol. 2021, 55, 9557–9568. [Google Scholar] [CrossRef]
- Mäkinen, K.; Elfving, M.; Hänninen, J.; Laaksonen, L.; Rajasilta, M.; Vuorinen, I.; Suomela, J.-P. Fatty Acid Composition and Lipid Content in the Copepod Limnocalanus Macrurus during Summer in the Southern Bothnian Sea. Helgol. Mar. Res. 2017, 71, 11. [Google Scholar] [CrossRef] [Green Version]
- Harmelin-Vivien, M.; Bănaru, D.; Dromard, C.R.; Ourgaud, M.; Carlotti, F. Biochemical Composition and Energy Content of Size-Fractionated Zooplankton East of the Kerguelen Islands. Polar Biol. 2019, 42, 603–617. [Google Scholar] [CrossRef]
- Basu, S.; Chanda, A.; Gogoi, P.; Bhattacharyya, S. Organochlorine Pesticides and Heavy Metals in the Zooplankton, Fishes, and Shrimps of Tropical Shallow Tidal Creeks and the Associated Human Health Risk. Mar. Pollut. Bull. 2021, 165, 112170. [Google Scholar] [CrossRef]
- UNEP. Report of the Persistent Organic Pollutants Review Committee on the Work of Its Seventeenth Meeting. In Proceedings of the Seventeenth Meeting of the Persistent Organic Pollutants Review Committee, Geneva, Switzerland, 24–28 January 2022; p. 56. [Google Scholar]
- Marsili, L.; Jiménez, B.; Borrell, A. Chapter 7—Persistent Organic Pollutants in Cetaceans Living in a Hotspot Area: The Mediterranean Sea. In Marine Mammal Ecotoxicology; Fossi, M.C., Panti, C., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 185–212. ISBN 978-0-12-812144-3. [Google Scholar]
- Marsili, L.; Coppola, D.; Giannetti, M.; Casini, S.; Fossi, M.C.; van Wyk, J.; Sperone, E.; Tripepi, S.; Micarelli, P.; Rizzuto, S. Skin Biopsies as a Sensitive Non-Lethal Technique for the Ecotoxicological Studies of Great White Shark (Carcharodon carcharias) Sampled in South Africa. Expert Opin. Environ. Biol. 2016, 4, 1000126. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, R.F. Distribution Patterns of Living and Subfossil Podocopid Ostracodes in the Nosy Bé Area, Northern Madagascar. Univ. Kans. Paleontol. Contrib. 1966, 72, 12. [Google Scholar]
- Goodman, S.M.; Benstead, J.P. (Eds.) The Natural History of Madagascar; University of Chicago Press: Chicago, IL, USA, 2003; ISBN 9780226303062. [Google Scholar]
- Jonahson, M.; Harding, S. Occurrence of Whale Sharks (Rhincodon typus) in Madagascar. Fish. Res. 2007, 84, 132–135. [Google Scholar] [CrossRef]
- Kiszka, J.; van der Elst, R. 11. Elasmobranchs (Sharks and Rays): A Review of Status, Distribution and Interaction with Fisheries of the Southwest Indian Ocean. In Offshore Fisheries of the Southwest Indian Ocean; Oceanographic Research Institute: Durban, South Africa, 2015; pp. 365–390. [Google Scholar]
- Brenier, A.; Vogel, A. Integrating Conservation and Development in Madagascar’s Marine Protected Areas. In Marine Protected Areas: Interactions with Fishery Livelihoods and Food Security; FAO: Rome, Italy, 2017; ISBN 9789251096062. [Google Scholar]
- Diamant, S.; Rohner, C.A.; Kiszka, J.J.; d’Echon, A.G.; d’Echon, T.G.; Sourisseau, E.; Pierce, S.J. Movements and Habitat Use of Satellite-Tagged Whale Sharks off Western Madagascar. Endanger. Species Res. 2018, 36, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Rohner, C.A.; Richardson, A.J.; Prebble, C.E.M.; Marshall, A.D.; Bennett, M.B.; Weeks, S.J.; Cliff, G.; Wintner, S.P.; Pierce, S.J. Laser Photogrammetry Improves Size and Demographic Estimates for Whale Sharks. PeerJ 2015, 3, e886. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.D.; Eckert, S.A. Foraging Ecology of Whale Sharks (Rhincodon typus) within Bahía de Los Angeles, Baja California Norte, México. Fish. Res. 2007, 84, 47–64. [Google Scholar] [CrossRef]
- Giam, C.S.; Wong, M.K.; Hanks, A.R.; Sackett, W.M.; Richardson, R.L. Chlorinated Hydrocarbons in Plankton from the Gulf of Mexico and Northern Caribbean. Bull. Environ. Contam. Toxicol. 1973, 9, 376–382. [Google Scholar] [CrossRef]
- Linko, R.R.; Raniamaki, P.; Urpo, K. PCB Residues in Plankton and Sediment in the Southwestern Coast of Finland. Bull. Environ. Contam. Toxicol. 1974, 12, 733–738. [Google Scholar] [CrossRef]
- Joiris, C.R.; Overloop, W. PCBs and Organochlorine Pesticides in Phytoplankton and Zooplankton in the Indian Sector of the Southern Ocean. Antarct. Sci. 1991, 3, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Focardi, S.; Bargagli, R.; Corsolini, S. Organochlorines in Antarctic Marine Food Chain at Terranova Bay (Ross Sea). Korean J. Polar Res. 1993, 4, 73–77. [Google Scholar]
- Ray, S.; Paranjape, M.A.; Koenig, B.; Paterson, G.; Metcalfe, T.; Metcalfe, C. Polychlorinated Biphenyls and Other Organochlorine Compounds in Marine Zooplankton off the East Coast of Newfoundland, Canada. Mar. Environ. Res. 1999, 47, 103–116. [Google Scholar] [CrossRef]
- Fossi, M.C.; Borsani, J.F.; Di Mento, R.; Marsili, L.; Casini, S.; Neri, G.; Mori, G.; Ancora, S.; Leonzio, C.; Minutoli, R.; et al. Multi-Trial Biomarker Approach in Meganyctiphanes Norvegica: A Potential Early Indicator of Health Status of the Mediterranean “Whale Sanctuary”. Mar. Environ. Res. 2002, 54, 761–767. [Google Scholar] [CrossRef]
- Quental, T.; Ferreira, A.M.; Vale, C. The Distribution of PCBs and DDTs in Seston and Plankton along the Portuguese Coast. Acta Oecol. 2003, 24, S333–S339. [Google Scholar] [CrossRef]
- Berrojalbiz, N.; Dachs, J.; Ojeda, M.J.; Valle, M.C.; Castro-Jiménez, J.; Wollgast, J.; Ghiani, M.; Hanke, G.; Zaldivar, J.M. Biogeochemical and Physical Controls on Concentrations of Polycyclic Aromatic Hydrocarbons in Water and Plankton of the Mediterranean and Black Seas. Glob. Biogeochem. Cycles 2011, 25, GB4003. [Google Scholar] [CrossRef] [Green Version]
- Frouin, H.; Dangerfield, N.; Macdonald, R.W.; Galbraith, M.; Crewe, N.; Shaw, P.; Mackas, D.; Ross, P.S. Partitioning and Bioaccumulation of PCBs and PBDEs in Marine Plankton from the Strait of Georgia, British Columbia, Canada. Prog. Oceanogr. 2013, 115, 65–75. [Google Scholar] [CrossRef]
- Desforges, J.-P.W.; Dangerfield, N.; Shaw, P.D.; Ross, P.S. Heightened Biological Uptake of Polybrominated Diphenyl Ethers Relative to Polychlorinated Biphenyls Near-Source Revealed by Sediment and Plankton Profiles along a Coastal Transect in British Columbia. Environ. Sci. Technol. 2014, 48, 6981–6988. [Google Scholar] [CrossRef]
- Morales, L.; Dachs, J.; Fernández-Pinos, M.-C.; Berrojalbiz, N.; Mompean, C.; González-Gaya, B.; Jiménez, B.; Bode, A.; Ábalos, M.; Abad, E. Oceanic Sink and Biogeochemical Controls on the Accumulation of Polychlorinated Dibenzo-p-Dioxins, Dibenzofurans, and Biphenyls in Plankton. Environ. Sci. Technol. 2015, 49, 13853–13861. [Google Scholar] [CrossRef]
- Kang, Y.; Zhang, R.; Yu, K.; Han, M.; Pei, J.; Chen, Z.; Wang, Y. Organochlorine Pesticides (OCPs) in Corals and Plankton from a Coastal Coral Reef Ecosystem, South China Sea. Environ. Res. 2022, 214, 114060. [Google Scholar] [CrossRef]
- Wolff, M.S.; Thornton, J.; Fischbein, A.; Lilis, R.; Selikoff, I.J. Disposition of Polychlorinated Biphenyl Congeners in Occupationally Exposed Persons. Toxicol. Appl. Pharmacol. 1982, 62, 294–306. [Google Scholar] [CrossRef]
- Safe, S.; Bandiera, S.; Sawyer, T.; Zmudzka, B.; Mason, G.; Romkes, M.; Denomme, M.A.; Sparling, J.; Okey, A.B.; Fujita, T. Effects of Structure on Binding to the 2,3,7,8-TCDD Receptor Protein and AHH Induction—Halogenated Biphenyls. Environ. Health Perspect. 1985, 61, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Bush, B.; Snow, J.; Koblintz, R. Polychlorobiphenyl (PCB) Congeners,p,P′-DDE, and Hexachlorobenzene in Maternal and Fetal Cord Blood from Mothers in Upstate New York. Arch. Environ. Contam. Toxicol. 1984, 13, 517–527. [Google Scholar] [CrossRef]
- Clarke, J.U. Structure-Activity Relationships in PCBs: Use of Principal Components Analysis to Predict Inducers of Mixed-Function Oxidase Activity. Chemosphere 1986, 15, 275–287. [Google Scholar] [CrossRef]
- Marsili, L.; D’Agostino, A.; Bucalossi, D.; Malatesta, T.; Fossi, M.C. Theoretical Models to Evaluate Hazard Due to Organochlorine Compounds (OCs) in Mediterranean Striped Dolphin (Stenella coeruleoalba). Chemosphere 2004, 56, 791–801. [Google Scholar] [CrossRef]
- World Health Organization (WHO) (Ed.) DDT and Its Derivatives: Environmental Aspects; World Health Organization: Geneva, Switzerland, 1989; ISBN 9789241542838. [Google Scholar]
- Aguilar, A. Relationship of DDE/ΣDDT in Marine Mammals to the Chronology of DDT Input into the Ecosystem. Can. J. Fish. Aquat. Sci. 1984, 41, 840–844. [Google Scholar] [CrossRef]
- Borrell, A.; Aguilar, A. Variations in DDE Percentage Correlated with Total DDT Burden in the Blubber of Fin and Sei Whales. Mar. Pollut. Bull. 1987, 18, 70–74. [Google Scholar] [CrossRef]
- Tsydenova, O.; Minh, T.B.; Kajiwara, N.; Batoev, V.; Tanabe, S. Recent Contamination by Persistent Organochlorines in Baikal Seal (Phoca Sibirica) from Lake Baikal, Russia. Mar. Pollut. Bull. 2004, 48, 749–758. [Google Scholar] [CrossRef]
- Qiu, X.; Zhu, T.; Li, J.; Pan, H.; Li, Q.; Miao, G.; Gong, J. Organochlorine Pesticides in the Air around the Taihu Lake, China. Environ. Sci. Technol. 2004, 38, 1368–1374. [Google Scholar] [CrossRef]
- Muñoz-Arnanz, J.; Jiménez, B. New DDT Inputs after 30 Years of Prohibition in Spain. A Case Study in Agricultural Soils from South-Western Spain. Environ. Pollut. 2011, 159, 3640–3646. [Google Scholar] [CrossRef]
- Nowell, L.H.; Capel, P.D.; Dileanis, P.D. Pesticides in Stream Sediment and Aquatic Biota: Distribution, Trends, and Governing Factors; Pesticides in the Hydrologic System; Lewis Publishers: Boca Raton, FL, USA, 1999; ISBN 9781566704694. [Google Scholar]
- Viselli, R. La Contaminazione da DDT e Dai Suoi Derivati Nei Comparti Abiotici del Bacino del Basso Toce e del Lago Maggiore; Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 1999. [Google Scholar]
- Qiu, X.; Zhu, T.; Yao, B.; Hu, J.; Hu, S. Contribution of Dicofol to the Current DDT Pollution in China. Environ. Sci. Technol. 2005, 39, 4385–4390. [Google Scholar] [CrossRef]
- Qiu, X.; Zhu, T. Using the o,P′-DDT/p,P′-DDT Ratio to Identify DDT Sources in China. Chemosphere 2010, 81, 1033–1038. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Hu, J. Global Inventory, Long-Range Transport and Environmental Distribution of Dicofol. Environ. Sci. Technol. 2015, 49, 212–222. [Google Scholar] [CrossRef]
- Beasley, V.R. Direct and Indirect Effects of Environmental Contaminants on Amphibians. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780124095489. [Google Scholar]
- European Commission. Commission Delegated Regulation (EU) 2022/643 of 10 February 2022 Amending Regulation (EU) No 649/2012 of the European Parliament and of the Council as Regards the Listing of Pesticides, Industrial Chemicals, Persistent Organic Pollutants and Mercury and an Update of Customs Codes (Text with EEA Relevance). Off. J. Eur. Union 2022, 118, 14–54. [Google Scholar]
- Turgut, C.; Gokbulut, C.; Cutright, T.J. Contents and Sources of DDT Impurities in Dicofol Formulations in Turkey. Environ. Sci. Pollut. Res. 2009, 16, 214–217. [Google Scholar] [CrossRef]
- Global Environment Facility—GEF. Improvement of DDT-Based Production of Dicofol and Introduction of Alternative Technologies Including IPM for Leaf Mites Control in China; Global Environment Facility—GEF: Washington, DC, USA, 2006. [Google Scholar]
- Wong, F.; Alegria, H.A.; Bidleman, T.F.; Alvarado, V.; Angeles, F.; Galarza, A.Á.; Bandala, E.R.; de la Cerda Hinojosa, I.; Estrada, I.G.; Reyes, G.G.; et al. Passive Air Sampling of Organochlorine Pesticides in Mexico. Environ. Sci. Technol. 2009, 43, 704–710. [Google Scholar] [CrossRef]
- Li, J.; Zhang, G.; Qi, S.; Li, X.; Peng, X. Concentrations, Enantiomeric Compositions, and Sources of HCH, DDT and Chlordane in Soils from the Pearl River Delta, South China. Sci. Total Environ. 2006, 372, 215–224. [Google Scholar] [CrossRef]
- Aguilar, A.; Borrell, A.; Pastor, T. Biological Factors Affecting Variability of Persistent Pollutant Levels in Cetaceans. J. Cetacean Res. Manag. 1999, 83–116. [Google Scholar] [CrossRef]
- United States Department of State. Madagascar; Background Notes Series; Bureau of Public Affairs: Washington, DC, USA, 1987; pp. 1–7. [Google Scholar]
- Jenkins, C.N.; Van Houtan, K.S. Global and Regional Priorities for Marine Biodiversity Protection. Biol. Conserv. 2016, 204, 333–339. [Google Scholar] [CrossRef]
- Di Capua, I.; Micarelli, P.; Tempesti, J.; Reinero, F.R.; Buttino, I. Zooplankton Size Structure in the Gulf of Tadjoura (Djibouti) during Whale Shark Sighting: A Preliminary Study. Cah. Biol. Mar. 2021, 62, 290–294. [Google Scholar] [CrossRef]
- Diamant, S.; Pierce, S.J.; Rohner, C.A.; Graham, R.T.; Guillemain d’Echon, A.; Guillemain d’Echon, T.; Sourisseau, E.; Fidiarisandratra, L.C.; Bakary, G.; Trélanche, S.; et al. Population Structure, Residency, and Abundance of Whale Sharks in the Coastal Waters off Nosy Be, North-Western Madagascar. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 3492–3506. [Google Scholar] [CrossRef]
- Boldrocchi, G.; Bettinetti, R. Whale Shark Foraging on Baitfish off Djibouti. Mar. Biodivers. 2019, 49, 2013–2016. [Google Scholar] [CrossRef]
- Fox, S.; Foisy, I.; De La Parra Venegas, R.; Galván Pastoriza, B.E.; Graham, R.T.; Hoffmayer, E.R.; Holmberg, J.; Pierce, S.J. Population Structure and Residency of Whale Sharks Rhincodon Typus at Utila, Bay Islands, Honduras. J. Fish Biol. 2013, 83, 574–587. [Google Scholar] [CrossRef]
- Araujo, G.; Agustines, A.; Tracey, B.; Snow, S.; Labaja, J.; Ponzo, A. Photo-ID and Telemetry Highlight a Global Whale Shark Hotspot in Palawan, Philippines. Sci. Rep. 2019, 9, 17209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, J.; McGinty, N.; Machete, M.; Afonso, P. Whale Shark-Tuna Associations, Insights from a Small Pole-and-Line Fishery from the Mid-North Atlantic. Fish. Res. 2020, 229, 105598. [Google Scholar] [CrossRef]
- Montero-Quintana, A.N.; Ocampo-Valdez, C.F.; Vázquez-Haikin, J.A.; Sosa-Nishizaki, O.; Osorio-Beristain, M. Correction to: Whale Shark (Rhincodon Typus) Predatory Flexible Feeding Behaviors on Schooling Fish. J. Ethol. 2022, 40, 117. [Google Scholar] [CrossRef]
- Matsumoto, R.; Toda, M.; Matsumoto, Y.; Ueda, K.; Nakazato, M.; Sato, K.; Uchida, S. Chapter 2. Notes on Husbandry of Whale Sharks, Rhincodon Typus, in Aquaria. In The Elasmobranch Husbandry Manual II: Recent Advances in the Care of Sharks, Rays and Their Relatives; Ohio Biological Survey: Columbus, OH, USA, 2017; p. 504. [Google Scholar]
- FAO; WHO. Joint FAO/WHO Food Standards Programme: Codex Maximum Limits for Pesticides Residues; FAO: Rome, Italy; WHO: Geneva, Switzerland, 1986. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food Chain [CONTAM] Related to the Presence of Non Dioxin-like Polychlorinated Biphenyls (PCB) in Feed and Food. EFSA J. 2005, 3, 284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vertical | ||||
| Compound | N | Mean ± SD (Min–Max) | Median | SE |
| HCB | 7 | 2.80 ± 4.02 (0.47–10.9) | 1.35 | 1.64 |
| PCBs | 7 | 114.55 ± 160.04 (26.16–436.49) | 47.39 | 65.33 |
| DDTs | 7 | 44.20 ± 29.26 (15.17–93.57) | 34.65 | 11.95 |
| DDTs/PCBs | 7 | 0.64 ± 0.22 (0.21–0.83) | 0.64 | 0.09 |
| Passive | ||||
| Compound | N | Mean ± SD (Min–Max) | Median | SE |
| HCB | 5 | 1.65 ± 1.05 (0.70–2.93) | 1.08 | 0.47 |
| PCBs | 5 | 65.81 ± 25.68 (38.95–98.05) | 57.16 | 11.48 |
| DDTs | 5 | 44.40 ± 21.06 (23.65–74.98) | 37.27 | 9.42 |
| DDTs/PCBs | 5 | 0.65 ± 0.06 (0.61–0.76) | 0.64 | 0.03 |
| Active | ||||
| Compound | N | Mean ± SD (Min–Max) | Median | SE |
| HCB | 5 | 0.72 ± 0.19 (0.51–1.01) | 0.68 | 0.09 |
| PCBs | 5 | 41.07 ± 16.10 (28.55–67.91) | 34.69 | 7.20 |
| DDTs | 5 | 25.85 ± 6.02 (18.11–32.96) | 26.28 | 2.70 |
| DDTs/PCBs | 5 | 0.68 ± 0.22 (0.44–0.95) | 0.63 | 0.10 |
| Control | ||||
| Compound | N | Mean ± SD (Min–Max) | Median | SE |
| HCB | 11 | 0.89 ± 0.81 (0.41–3.31) | 0.68 | 0.24 |
| PCBs | 11 | 47.76 ± 14.49 (27.99–73.62) | 49.74 | 4.37 |
| DDTs | 11 | 30.21 ± 10.16 (14.54–54.46) | 26.81 | 3.06 |
| DDTs/PCBs | 11 | 0.65 ± 0.20 (0.46–1.09) | 0.60 | 0.06 |
| Area | Sample Type | PCBs | DDTs | HCB | Ref. |
|---|---|---|---|---|---|
| Gulf of Mexico and Caribbean | Zooplankton | <3–678 ng/g w.w. | 0.2–34 ng/g w.w. | [48] | |
| Turku Arcipelago (Finland) | Zooplankton | 38 ppm l.w. | [49] | ||
| Southern Ocean | Zooplankton and phytoplankton | 0.30–0.37 ng/g d.w. | 19 ng/g d.w. | [50] | |
| Terranova Bay (Antartide) | Zooplankton (copepods) | 575 ng/g l.w. | 400 ng/g l.w. | 109 ng/g l.w. | [51] |
| East coast of Newfoundland (Canada) | Zooplankton | 85.7 ng/g l.w. | 22.3 ng/g lw | 6.4 ng/g l.w. | [52] |
| Pelagos Sanctuary (Maditerranean Sea) | Zooplankton (Meganyctiphanes norvegica) | 84.6–210.2 ng/g w.w. | 45.3–163.2 ng/g w.w. | 3.5–11.6 ng/g w.w. | [53] |
| Portugal | Plankton | 61–159 ng/g d.w. (February) | [54] | ||
| 68–155 ng/g d.w. (April) | 48–76 ng/g d.w. (north) | ||||
| 12–63 ng/g d.w. (July) | 3–7 ng/g d.w. (south) | ||||
| Maditerranean Sea | Zooplankton | 0.76–353 ng/g d.w. | 2.5 ng/g d.w. | [55] | |
| Strait of Georgia British Columbia (Canada) | Zooplankton | 52.2–364 ng/g l.w. | [56] | ||
| Coastal Transect in British Columbia (Canada) | Zooplankton | 0.2–0.8 ng/g l.w. (north) | [57] | ||
| 0.6–1.2 ng/g l.w. (south) | |||||
| Atlantic, Indian and Pacific Oceans | Zooplankton | 30–692 pg/g d.w. | [58] | ||
| Gulf of Tadjoura (Djibouti) | Zooplankton | 109.7–636.1 ng/g d.w. | 21.42–79.2 ng/g d.w. | [2] | |
| Weizhou Island (China) | Zooplankton | 0.77 ± 0.20 ng/g d.w. | 0.20 ± 0.08 ng/g d.w. | [59] |
| Compound | Vertical N = 7 | Passive N = 5 | Active N = 5 | Control N = 11 |
|---|---|---|---|---|
| 95 | 3.48 ± 1.46 | 2.74 ± 2.41 | 3.26 ± 0.59 | 3.61 ± 2.94 |
| 101 | 3.14 ± 1.92 | 2.68 ± 1.49 | 2.81 ± 1.04 | 2.78 ± 0.71 |
| 99 | 3.44 ± 1.60 | 2.02 ± 0.75 | 3.73 ± 0.94 | 3.37 ± 0.92 |
| 151 | 2.28 ± 1.16 | 1.61 ± 0.55 | 2.45 ± 0.44 | 2.02 ± 0.70 |
| 144 + 135 | 1.89 ± 1.10 | 1.69 ± 0.67 | 2.32 ± 0.43 | 2.25 ± 1.08 |
| 149 + 118 | 8.40 ± 2.62 | 8.01 ± 3.18 | 8.40 ± 2.10 | 8.75 ± 1.53 |
| 146 | 2.57 ± 0.91 | 2.63 ± 2.77 | 3.02 ± 1.37 | 3.23 ± 0.79 |
| 153 | 14.82 ± 3.38 | 15.16 ± 3.56 | 13.80 ± 1.25 | 13.59 ± 5.51 |
| 138 | 9.18 ± 3.01 | 9.14 ± 1.81 | 7.36 ± 1.22 | 7.45 ± 3.07 |
| 178 | 2.38 ± 0.87 | 4.09 ± 1.16 | 3.33 ± 2.45 | 2.54 ± 1.34 |
| 187 | 4.43 ± 1.02 | 5.47 ± 2.04 | 4.32 ± 0.66 | 3.94 ± 1.49 |
| 183 | 2.05 ± 0.43 | 2.85 ± 2.34 | 1.98 ± 0.58 | 2.38 ± 0.84 |
| 128 | 1.37 ± 0.48 | 1.61 ± 0.78 | 1.16 ± 0.26 | 1.10 ± 0.29 |
| 174 | 2.85 ± 1.01 | 3.90 ± 1.46 | 4.05 ± 2.52 | 2.90 ± 1.22 |
| 177 | 1.75 ± 0.49 | 1.76 ± 0.55 | 1.71 ± 0.51 | 1.46 ± 0.46 |
| 156 + 171 + 202 | 1.55 ± 0.39 | 1.93 ± 0.84 | 1.24 ± 0.42 | 1.32 ± 0.30 |
| 172 | 1.23 ± 0.40 | 1.41 ± 0.74 | 2.34 ± 0.85 | 1.23 ± 0.44 |
| 180 | 7.66 ± 6.94 | 8.02 ± 5.26 | 5.06 ± 1.12 | 4.70 ± 1.75 |
| 199 | 2.17 ± 1.24 | 2.49 ± 2.67 | 3.15 ± 1.06 | 1.53 ± 0.40 |
| 170 | 5.54 ± 2.94 | 5.51 ± 3.28 | 3.77 ± 2.09 | 4.68 ± 1.69 |
| 196 | 3.40 ± 0.70 | 2.09 ± 0.52 | 2.62 ± 0.97 | 3.23 ± 2.10 |
| 201 | 4.43 ± 3.34 | 5.20 ± 2.07 | 4.31 ± 1.54 | 10.56 ± 11.44 |
| 195 | 2.85 ± 1.82 | 4.02 ± 3.43 | 4.61 ± 2.26 | 4.44 ± 2.39 |
| 194 | 2.29 ± 0.96 | 1.95 ± 1.19 | 2.97 ± 1.17 | 2.31 ± 1.40 |
| 206 | 5.31 ± 2.91 | 4.53 ± 2.68 | 6.69 ± 1.51 | 5.48 ± 2.31 |
| Compound | Vertical N = 7 | Passive N = 5 | Active N = 5 | Control N = 11 |
|---|---|---|---|---|
| op′DDE | 6.11±4.59 | 8.84 ± 7.45 | 2.98 ± 1.17 | 7.25 ± 5.15 |
| op′DDD | 8.10 ± 2.43 | 5.41 ± 2.30 | 7.94 ± 1.89 | 7.45 ± 1.79 |
| op′DDT | 20.86 ± 10.12 | 24.91 ± 13.42 | 20.01 ± 5.30 | 26.66 ± 9.15 |
| pp′DDE | 49.45 ± 9.90 | 39.78 ± 9.25 | 42.49 ± 8.21 | 41.73 ± 11.25 |
| pp′DDT | 8.59 ± 3.60 | 13.95 ± 8.96 | 20.20 ± 11.85 | 12.27 ± 4.57 |
| pp′DDD | 6.89 ± 1.61 | 7.12 ± 3.00 | 6.38 ± 2.36 | 6.62 ± 2.80 |
| pp′DDE/pp′DDT | 6.91 ± 3.68 | 4.39 ± 3.67 | 2.89 ± 1.91 | 4.08 ± 2.43 |
| (pp′DDE + pp′DDD)/pp′DDT | 7.80 ± 3.96 | 5.26 ± 4.52 | 3.30 ± 2.18 | 4.68 ± 2.59 |
| pp′DDE/DDTs | 0.49 ± 0.10 | 0.40 ± 0.09 | 0.42 ± 0.08 | 0.42 ± 0.11 |
| op′DDTs/DDTs | 0.35 ± 0.09 | 0.39 ± 0.11 | 1.47 ± 1.33 | 0.40 ± 0.10 |
| op′DDT/pp′DDT | 2.91 ± 2.45 | 2.70 ± 2.10 | 1.47 ± 1.33 | 2.45 ± 1.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsili, L.; Consales, G.; Romano, P.; Rosai, R.; Bava, P.; Reinero, F.R.; Micarelli, P. A Cocktail of Plankton and Organochlorines for Whale Shark in the Foraging Areas of Nosy Be (Madagascar). Diversity 2023, 15, 911. https://doi.org/10.3390/d15080911
Marsili L, Consales G, Romano P, Rosai R, Bava P, Reinero FR, Micarelli P. A Cocktail of Plankton and Organochlorines for Whale Shark in the Foraging Areas of Nosy Be (Madagascar). Diversity. 2023; 15(8):911. https://doi.org/10.3390/d15080911
Chicago/Turabian StyleMarsili, Letizia, Guia Consales, Patrizia Romano, Rachele Rosai, Paolo Bava, Francesca Romana Reinero, and Primo Micarelli. 2023. "A Cocktail of Plankton and Organochlorines for Whale Shark in the Foraging Areas of Nosy Be (Madagascar)" Diversity 15, no. 8: 911. https://doi.org/10.3390/d15080911
APA StyleMarsili, L., Consales, G., Romano, P., Rosai, R., Bava, P., Reinero, F. R., & Micarelli, P. (2023). A Cocktail of Plankton and Organochlorines for Whale Shark in the Foraging Areas of Nosy Be (Madagascar). Diversity, 15(8), 911. https://doi.org/10.3390/d15080911