
Resettlement of Eurytemora velox (Crustacea: Copepoda) in Europe, the Urals and Western Siberia



Abstract
:1. Introduction
2. Material and Methods
2.1. Material and Methods Used for Morphological Analysis
2.2. Genetic Studies
3. Results
3.1. Genetic Studies
3.2. Morphological Studies
4. Discussion
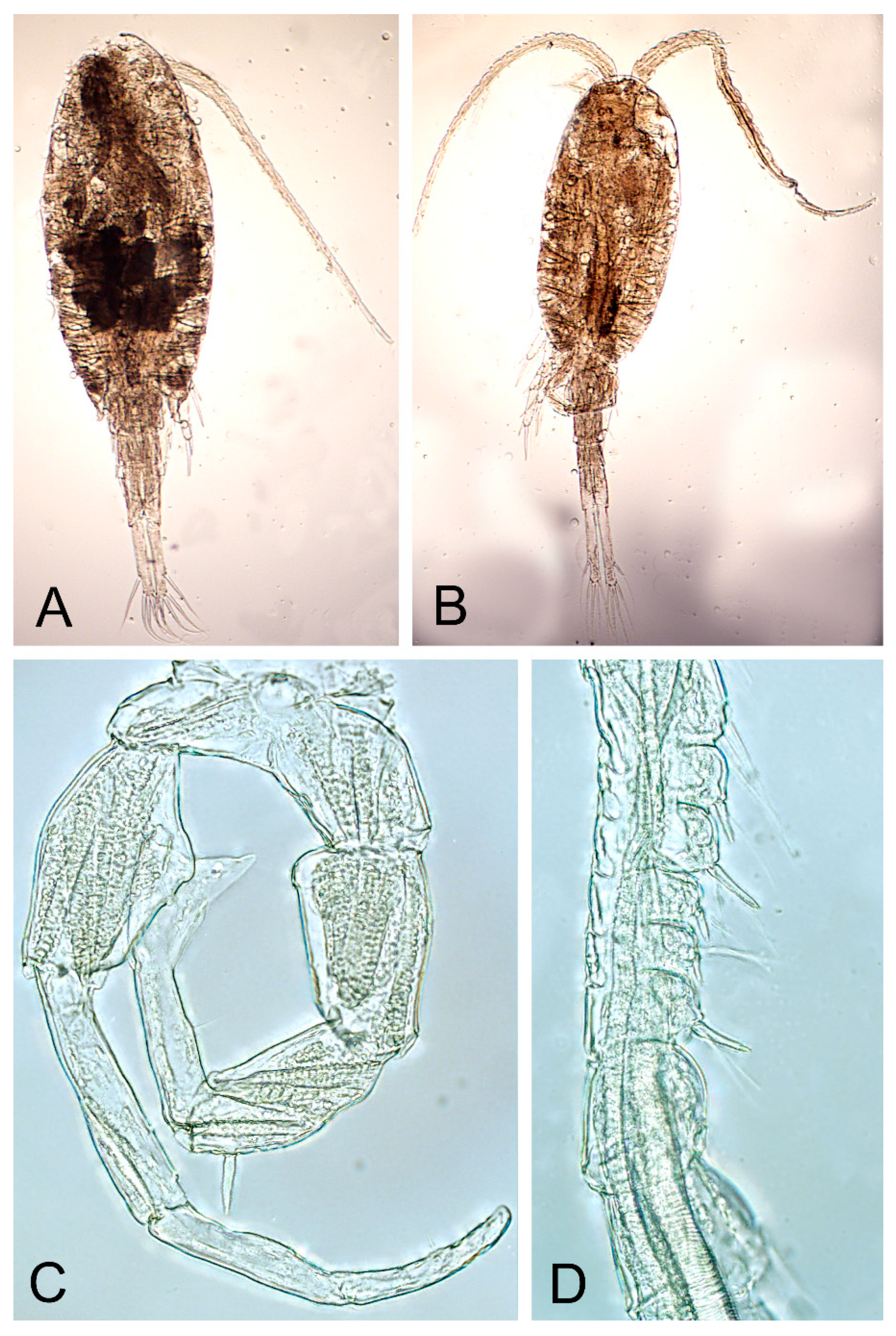
4.1. Morphological Studies
4.2. Genetic Studies
4.3. Urals and Western Siberian Populations
4.4. Baltic Population
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lilljeborg, W. De Crustaceis ex Ordinibus Tribus: Cladocera, Ostracoda et Copepoda, in Scania Occurrentibus; Berlingska Boktryckeriet: Lund, Sweden, 1853; pp. 1–222. Available online: https://www.biodiversitylibrary.org/page/39045453 (accessed on 12 December 2023).
- Sars, G.O. Temorella lacinulata (Fischer). In Yearbook of the Zoological Museum of the Imperial Academy of Sciences; Printing House of the Imperial Academy of Sciences: St. Petersburg, Russia, 1897; p. 66. [Google Scholar]
- Pandourski, I.; Stoichev, S. Sur la faune de l’eau interstitielle littorale de la bande sableuse entre le système lacustre de “Chabla-Ezeretz” et la Mer Noire. Hist. Nat. Bulg. 1999, 10, 125–131. [Google Scholar]
- Pandourski, I.; Evtimova, V. First record of Eurytemora velox (Lilljeborg, 1853) (Crustacea, Copepoda, Calanoida) in Iceland with morphological notes. Hist. Nat. Bulg. 2006, 17, 35–38. [Google Scholar]
- Vranovský, M. Eurytemora velox (Lilljeborg, 1853) (Crustacea, Copepoda), a new immigrant in the middle Danube. Biol. Bratisl. 1994, 49, 167–172. [Google Scholar]
- Samchyshyna, L.; Gromova, Y.; Zorina-Sakharova, K. Recent distribution of Eurytemora velox (Lilljeborg, 1853) (Copepoda, Calanoida) in brackish and fresh waters of Ukraine. Crustaceana 2020, 93, 275. [Google Scholar] [CrossRef]
- Quintana, X.; Antón-Pardo, M.; Bas-Silvestre, M.; Boix, D.; Casamitjana, X.; Compte, J.; Cunillera-Montcusí, D.; Gascón, S.; Menció, A.; Obrador, B.; et al. Identifying critical transitions in seasonal shifts of zooplankton composition in a confined coastal salt marsh. Aquat. Sci. 2021, 83, 69. [Google Scholar] [CrossRef]
- Borutsky, E.S.; Stepanova, L.A.; Kos, M.S. Key to Calanoida Fresh Waters of the USSR; Nauka: Leningrad, Russia, 1991; p. 504. (In Russian) [Google Scholar]
- Sabaneev, P.L. Eurytemora velox (Lill.) in the flat part of Dnieper. Zb. Pr. Dnipr. Biol. Stn. 1930, 5, 201–203. [Google Scholar]
- Ermolaeva, N.I. Zooplankton of different types of water bodies of the Yamal Peninsula in 2015. Sci. Bull. Yamalo-Nenets Auton. Okrug 2016, 2, 56–62. (In Russian) [Google Scholar]
- Ermolaeva, N.I. Species composition and spatial distribution of zooplankton in the Gulf of Ob and Gydan Bay. In Proceedings of the III All-Russian Scientific Conference with International Participation, Yekaterinburg, Russia, 5–10 October 2020; Institute of Water and Environmental Problems SB RAS: Novosibirsk, Russia, 2017; pp. 91–99. (In Russian). [Google Scholar]
- Abdullina, G.K.; Bondar, M.S. Zooplankton of water bodies of the Arctic tundra of the Yamal Peninsula. In Proceedings of the XXI International Scientific and Practical Conference I, Moscow, Russia, 20 May 2019; pp. 297–304. (In Russian). [Google Scholar]
- Gerasimova, A.; Sharapova, T.; Gerasimov, A.; Babushkin, E.; Gerasimova, A. Alien Species of Hydrobionts in the Tyumen Region. Version 1.4. Institute of the Problems of Northern Development is a Subdivision of Tyumen Scientific Centre SB RAS. Occurrence Dataset. 2022. Available online: https://www.gbif.org/dataset/93d951ea-c37d-4062-9007-dbe966f7f9d9 (accessed on 29 August 2023).
- Semenova, L.A.; Aleksyuk, V.A. Zooplankton of the Lower Ob. Bull. Ecol. For. Landsc. Sci. 2010, 156–169. (In Russian) [Google Scholar]
- Ermolaeva, N.I. The fauna of brackish-water and freshwater invertebrates (Rotifera, Cladocera, Copepoda) of lakes in the south of Western Siberia. In Proceedings of the Materials of the International Scientific Internet Conference «Biodiversity of Terrestrial and Aquatic Animals and Bioresources», Kazan, Russia, 12 February 2013. (In Russian). [Google Scholar]
- Yermolaeva, N. Some results concerning zooplankton species composition in the lakes of the south of Ob-Irtysh interfluve. World Sci. Cult. Educ. 2015, 50, 390–398. [Google Scholar]
- Abramova, E.; Vishnyakova, I.; Boike, J.; Abramova, A.; Solovyev, G.; Martynov, F. Structure of freshwater zooplankton communities from tundra waterbodies in the Lena River Delta, Russian Arctic, with a discussion on new records of glacial relict copepods. Polar Biol. 2017, 40, 1629–1643. [Google Scholar] [CrossRef]
- Chertoprud, E.S.; Novichkova, A.A. Crustaceans in the meiobenthos and plankton of the thermokarst lakes and polygonal ponds in the Lena River Delta (Northern Yakutia, Russia): Species composition and factors regulating assemblage structures. Water 2021, 13, 1936. [Google Scholar] [CrossRef]
- Chertoprud, E.S.; Novichkova, A.A.; Tsyganov, A.N.; Vorobjeva, L.V.; Esaulov, A.S.; Krylenko, S.V.; Mazei, Y.A. Species diversity and driving factors of benthic and zooplanktonic assemblages at different stages of thermokarst lake development: A case study in the Lena River delta (Middle Siberia). Diversity 2023, 15, 511. [Google Scholar] [CrossRef]
- Rechkalov, V.V.; Kuznetsova, A. Study of zooplankton in the Magnitogorsk Reservoir. Bull. Chelyabinsk St. Univ. 2008, 17, 91–97. (In Russian) [Google Scholar]
- Rechkalov, V.V.; Permyakova, T.A. Composition and seasonal dynamics of zooplankton in Lake Second (Chelyabinsk region). Bull. Chelyabinsk St. Univ. 2010, 4, 86–93. (In Russian) [Google Scholar]
- Golubok, O.V.; Rechkalov, V.V. Identification of signs of anthropogenic eutrophication of dimictic lakes based on indicators of zooplankton development. Bull. Orenbg. State Univ. 2013, 10, 61–68. (In Russian) [Google Scholar]
- Muraveysky, S.D. Observations on the spring plankton of the river. The Urals and its oxbow lakes. Russ. Hydrobiol. J. 1923, 11, 14–23. (In Russian) [Google Scholar]
- Bening, A.L. Materials on the Hydrobiology of the River Ural. Proc. KazFAN USSR 1938, 11, 153–157. (In Russian) [Google Scholar]
- Saduakasova, P.E. Zooplankton in the lower reaches of the Ural River. Fish resources of water bodies of Kazakhstan and their use. Alma-Ata Sci. 1966, 5, 346–351. (In Russian) [Google Scholar]
- Demesinova, G.T.; Kurashov, E.A. Current state of zooplankton in the lower reaches of the Ural River. Water Chem. Ecol. 2016, 12, 42–53. [Google Scholar]
- Troshina, T.T.; Idayatov, P.V.; Gorelov, O.P. Zooplankton of Lake Chelkar (June, 2005). Biodiversity of the animal world of Kazakhstan, problems of conservation and use. In Book of Abstracts of Conference: 75-th Anniversary of the Institute of Zoology “Biodiversity of the Fauna of Kazakhstan, Problems of Conservation and Use.” 17–20 October; Adebiet: Almaty, Kazakhstan, 2007; pp. 81–83. (In Russian) [Google Scholar]
- Troshina, T.T.; Idayatov, P.V.; Gorelov, O.P. Structural and functional characteristics of zooplankton in the Bitik reservoir and lake. In Book of Abstracts of Conference: «Biodiversity of the Animal World of Kazakhstan, Problems of Conservation and Use". 17–20 October; Adebiet: Almaty, Kazakhstan, 2007; pp. 84–86. (In Russian) [Google Scholar]
- Krupa, E.G.; Dobrokhotova, O.V.; Stuge, T.S. Fauna of Calanoida (Crustacea: Copepoda) of Kazakhstan and Adjacent Territories; Etalon Print: Almaty, Kazakhstan, 2016; p. 208. (In Russian) [Google Scholar]
- Vezhnovets, V.; Litvinova, A. Representatives of the genus Eurytemora Giesbrecht, 1881 (Calanoida, Temoridae) in aquatic ecosystems of Belarus. Crustaceana 2020, 93, 283–298. [Google Scholar] [CrossRef]
- Sukhikh, N.M.; Lazareva, V.I. First Results of a Molecular Genetic Analysis of the European Invader Eurytemora velox (Crustacea, Calanoida). Inland Water Biol. 2022, 15, 201–203. [Google Scholar] [CrossRef]
- Sukhikh, N.M.; Fefilova, E.B. The first record of Eurytemora velox (Lilljeborg, 1853) (Crustacea, Calanoida) outside of Europe, genetic identification with surprise. Russ. J. Biol. Invasions 2023, 4, 139–140. [Google Scholar] [CrossRef]
- Rymaszewska, A.; Kirczuk, L. Eurytemora carolleeae (Alekseev & Souissi, 2011) as a Non-Native Copepod Invader into Central European Large River. Available online: https://www.ncbi.nlm.nih.gov/nuccore/?term=Eurytemora+velox (accessed on 12 December 2023).
- Sukhikh, N.M.; Lazareva, V.I.; Alekseev, V.R. Copepod Eurytemora caspica Sukhikh et Alekseev, 2013 (Crustacea, Calanoida) in Volga and Kama River Reservoirs. Inland Water Biol. 2020, 13, 198–205. [Google Scholar] [CrossRef]
- Sukhikh, N.M.; Souissi, A.; Souissi, S.; Winkler, G.; Castric, V.; Holl, A.-C.; Alekseev, V. Genetic and morphological heterogeneity among populations of Eurytemora affinis (Crustacea: Copepoda: Temoridae) in European waters. Comptes Rendus Biol. 2016, 339, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Gaviria Melo, S.; Forro, L. Morphological characterization of new populations of the copepod Eurytemora velox (Lilljeborg, 1853) (Calanoida, Temoridae) found in Austria and Hungary. Hydrobiologia 2000, 438, 205–216. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.J.; Gibson, T.J.; Clustal, W. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Res. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Winkler, G.; Souissi, S.; Poux, C.; Castric, V. Genetic heterogeneity among Eurytemora affinis populations in Western Europe. Mar. Biol. 2011, 158, 1841–1856. [Google Scholar] [CrossRef]
- Takezaki, N.; Rzhetsky, A.; Nei, M. Phylogenetic test of the molecular clock and linearized trees. Mol. Biol. Evol. 1995, 12, 823–833. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef]
- Knowlton, N.; Weigt, L.A. New dates and new rates for divergence across the Isthmus of Panama. Proceeding R. Soc. B Biol. Sci. 1998, 265, 2257–2263. [Google Scholar] [CrossRef]
- Zofkova, M.; Timms, B.V. A conflict of morphological and genetic patterns in the australian anostracan Branchinella longirostris. Hydrobiologia 2009, 635, 67–80. [Google Scholar] [CrossRef]
- Marino, I.A.M.; Pujolar, J.M.; Zane, L. Reconciling deep calibration and demographic history: Bayesian inference of post glacial colonization patterns in Carcinus aestuarii (Nardo, 1847) and C. maenas (Linnaeus, 1758). PLoS ONE 2011, 6, e28567. [Google Scholar] [CrossRef] [PubMed]
- Ketmaier, V.; Marrone, F.; Alfonso, G.; Paulus, K.; Wiemann, A.; Tiedemann, R.; Mura, G. Mitochondrial DNA regionalism and historical demography in the extant populations of Chirocephalus kerkyrensis (Branchiopoda: Anostraca). PLoS ONE 2012, 7, e30082. [Google Scholar] [CrossRef] [PubMed]
- Sworobowicz, L.; Mamos, T.; Grabowski, M.; Wysocka, A. Lasting through the ice age: The role of the proglacial refugia in the maintenance of genetic diversity, population growth, and high dispersal rate in a widespread freshwater crustacean. Freshw. Biol. 2020, 65, 1028–1046. [Google Scholar] [CrossRef]
- Kotov, A.A.; Taylor, D.J. Mesozoic fossils (>145 Mya) suggest the antiquity of the subgenera of Daphnia and their coevolution with chaoborid predators. BMC Evol. Biol. 2011, 11, 129. [Google Scholar] [CrossRef]
- Wolfe, J.M.; Daley, A.C.; Legg, D.A.; Edgecombe, G. Fossil calibrations for the arthropod Tree of Life. Earth-Sci. Rev. 2016, 160, 43–110. [Google Scholar] [CrossRef]
- Eyun, S. Phylogenomic analysis of Copepoda (Arthropoda, Crustacea) reveals unexpected similarities with earlier proposed morphological phylogenies. BMC Evol. Biol. 2017, 17, 23. [Google Scholar] [CrossRef]
- Lucks, R. Die crustaceen und rotatorien des messinasees. Ber. Westpreuss Bol-Zool. 1937, 59, 59–101. [Google Scholar]
- Petit, G.; Schachter, D. Contributions a l’titude biologique de la Camargue. Sur la presence du genre Eurytemora en Camargue. Bull. Mus. Hist. Nat. Marseille 1947, 7, 103–114. [Google Scholar]
- Forro, L.; Gulyzis, P. Eurytemora velox (Li11jeborg. 1853) (Copepoda. Calanoida). Misc. Zool. Hung. 1992, 7, 53–58. [Google Scholar]
- Smirnov, C.C. On the so-called hybrids in the genus Eurytemora Giesbr. (Copepoda). Rep. Acad. Sci. USSR 1940, 26, 615–618. (In Russian) [Google Scholar]
- Bucklin, A.; LaJeunesse, T.C. Molecular genetic variation of Calanus pacificus (Copepoda: Calanoida): Preliminary evaluation of genetic structure and subspecific differentiation based on mt-DNA sequences. Calif. Coop. Ocean. Fish. Investig. Rep. 1994, 35, 45–51. [Google Scholar]
- Kochanova, E.; Nair, A.; Sukhikh, N.; Väinölä, R.; Husby, A. Patterns of cryptic diversity and phylogeography in four freshwater copepod crustaceans in European lakes. Diversity 2021, 13, 448. [Google Scholar] [CrossRef]
- Kiefer, F. Freilebende Copepoda. Das Zooplankton der Binnengewässer; Teil 2, E. Schweizerbartsche Verlag: Stuttgart, Germany, 1978; p. 380. [Google Scholar]
- Ekmann, S. Uber das crustacea plankton des Ekoln (Mälaren) und überver schiedene Kategorien von marinen Relikten in schwedischen Binnenseen. Zool. Stud. Tillägn. T. Tullberg. Upsala Almqvist Wik-Sells 1907, 42–65. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Drainage Basin | Number of Morphologically Studied Specimens | Number of Genetically Studied Specimens | Date of Sampling | Collector |
|---|---|---|---|---|---|
| Rybinsk Reservoir, Russia | Volga River, Caspian Sea basin | 8 females | 2017 | Valentina Lazareva, IBIV RAS | |
| Cheboksary Reservoir, Russia | Volga River, Caspian Sea basin | 1 female | 4 females | 2020 | Valentina Lazareva, IBIV RAS [31] |
| Kuybyshev Reservoir, Russia | Volga River, Caspian Sea basin | 1 female | 2018 | Valentina Lazareva, IBIV RAS | |
| Feofania ponds, Ukraine | Dnieper River, Black Sea basin | 7 females | 3 females | 2020 | Larysa Samchyshyna, IF NAAS [31] |
| Mukhavets River, Belarus | Western Bug, Baltic Sea basin | 6 females | 2019 | Vasilii Vezhnovets, NAS of Belarus | |
| Ob Bay, Russia | Kara Sea basin | 11 females | 2 females | 2022 | Petr Garibian, IEE RAS |
| Magnitogorsk Lake | Ob River, Kara Sea basin | 14 females | 2 females | 2022 | Alisa Neplyukhina, IEE RAS |
| Lake Smolino, Southern Urals region, Russia | Ob River, Kara Sea basin | 7 females | 2021 | Maria Baturina, IB Komi SC UB RAS [32] | |
| Dziwna Channel, Poland | Baltic Sea basin | 4 specimens | 2020 | [33] |
| Trait | Gulf of Ob | Magnitogorsk | Kyiv | Belarus | Rybinsk Reservoir | Cheboksary Reservoir | Kuibyshev Reservoir |
|---|---|---|---|---|---|---|---|
| Female | |||||||
| Body length, mm mean +/− SD | 1.85 +/− 0.11 | 1.61 +/− 0.06 | 1.77 +/− 0.02 | 1.51 +/− 0.02 | 1.43 +/− 0.06 | 1.48 | 1.52 |
| Body length, mm range | 1.63–1.97 | 1.54–1.72 | 1.68–1.86 | 1.42–1.58 | 1.68–1.87 | ||
| N of studied individuals | 11 | 14 | 7 | 6 | 8 | 1 | 1 |
| Body Length/Weight | 2.89 | 3.16 | 3.9 +/− 0.06 | 3.5 +/− 0.08 | 3.6 +/− 0.09 | 3.32 | 3.31 |
| Thoracic wings | + | + | + | + | + | + | + |
| Abdominal segments 1–3 length ratio | 1.62/1/1.83 | 1.53/1/1.84 | 1.71/1/1.97 | 1.79/1/1.87 | 1.68/1/1.93 | 1.79/1/1.82 | 1.88/1/1.8 |
| Furcal L/W | 5.73 +/− 0.05 | 5.06 +/− 0.07 | 5.83 +/− 0.18 | 5.27 +/− 0.15 | 4.67 +/− 0.2 | ||
| Furcal L/W range | 5.48–5.95 | 4.74–5.62 | 4.84–6.93 | 4.42–6.16 | 4.31–6.17 | ||
| Body L/Furcal L | 7.91 +/− 0.14 | 8.3 +/− 0.20 | 5.91 +/− 0.09 | 6.28 +/− 0.16 | 6.47 +/− 0.08 | ||
| Body L/Furcal L range | 6.79–8.48 | 6.91–9.53 | 5.37–6.58 | 5.57–7.77 | 6.16–6.91 | ||
| Furcal W/Body W | 0.06 +/− 0.00 | 0.08 +/− 0.00 | 0.12 +/− 0.00 | 0.11 +/− 0.00 | 0.12 +/− 0.00 | ||
| Furcal W/Body W range | 0.06–0.07 | 0.06–0.09 | 0.10–0.14 | 0.10–0.13 | 0.10–0.13 | ||
| Furcal L/Body W | 0.37 +/− 0.01 | 0.39 +/− 0.01 | 0.69 +/− 0.15 | 0.58 +/− 0.02 | 0.58 +/− 0.15 | ||
| Furcal L/Body W range | 0.34–0.4 | 0.35–0.46 | 0.58–0.78 | 0.46–0.64 | 0.52–0.65 | ||
| Furcal W/Body L | 0.02 +/− 0.00 | 0.02 +/− 0.00 | 0.03 +/− 0.00 | 0.03 +/− 0.00 | 0.03 +/− 0.00 | ||
| Furcal W/Body L Range | 0.021–0.025 | 0.020–0.028 | 0.025–0.036 | 0.028–0.034 | 0.026–0.034 | ||
| P5th leg. Exo. 1 | |||||||
| P5 right leg/left leg: one spine/one spine | + | + | + | + | + | + | + |
| P5 right leg/left leg: one spine/two spines | + | − | − | − | − | − | − |
| P5 right leg/left leg: two spines/one spine | + | − | − | + | − | − | − |
| P5 right leg/left leg: two spines/two spines | + | − | − | − | − | − | − |
| Male | |||||||
| Body length mean +/− SD | 1.69 +/− 0.11 | 1.35 +/− 0.1 | 1.09 +/− 0.17 | 1.14 +/− 0.01 | |||
| Body length range | 1.54–1.86 | 1.23–1.51 | 1.06–1.13 | 1.12–1.15 | |||
| N of studied individuals | 11 | 11 | 4 | 2 | |||
| Right 1st antennular spine 10 | + | + | +/− | − | |||
| Right 1st antennular spine 11 | + | + | +/− | − | |||
| Furcal L/W | 7.24 +/− 0.07 | 5.92 +/− 0.13 | 5.86 +/− 0.16 | 5.43 +/− 0.46 | |||
| Furcal L/W range | 6.88–7.65 | 4.85–6.43 | 5.24–9.46 | 5.17–6.04 | |||
| Body L/Furcal L | 6.67 +/− 0.27 | 7.85 +/− 0.29 | 6.00 +/− 0.09 | 7.54 +/− 0.26 | |||
| Body L/Furcal L range | 5.74–8.45 | 6.75–9.44 | 5.64–6.36 | 6.86–8.05 | |||
| Furcal W/Body L | 0.021 +/− 0.001 | 0.022 +/− 0.001 | 0.029 +/− 0.001 | 0.025 +/− 0.002 | |||
| Furcal W/Body L range | 0.017–0.024 | 0.018–0.026 | 0.026–0.032 | 0.021–0.028 | |||
| Left 5th leg length ratio Cox:Bas.:Exo1:Exo2 | 1/1.46/1.7/2.25 | 1/1.6/1.8/2.2 | 1/1.15/1.38/1.73 | 1/1.2/1.4/1.9 | |||
| Right 5th leg length ratio Cox:Bas:Exo1:Exo2:Exo3 | 1/2.3/2.4/2.16/1.55 | 1/2.3/2.8/2.3/1.5 | 1/2.1/2.3/2.0/1.83 | 1/2.0/2.3/1.9/1.7 |
| Sampling Place | CO1 | nITS | 18SrRNA | Source |
|---|---|---|---|---|
| Cheboksary Reservoir, Russia | 2 sequences MZ373321-MZ373322 | 3 sequences MZ400502-MZ400504 | 1 sequence MZ373302 | [31] |
| Feofania ponds in Kyiv, Ukraine | 3 sequences MZ373318-Z373320 | 3 sequences MZ400499-MZ400501 | OR921659 | [31] |
| Gulf of Ob, RF | 1 sequence OR578622 | 1 sequence OR583034 | 2 sequences OR578728-OR578729 | This paper |
| Magnitogorsk Lake, Russia | 2 sequences OR583032-OR583033 | 2 sequences OR921657- OR921658 | This paper | |
| Lake Smolino, Southern Urals region, Russia | 3 sequences OR578619-OR578621 | 7 sequences OR583025-OR583031 | 3 sequences OR578725-OR578727 | [32] |
| Dziwna Channel, Poland | 2 sequences MT146446-MT146445 | 3 sequences MT787212-MT787214 | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukhikh, N.; Garibian, P.; Chertoprud, E. Resettlement of Eurytemora velox (Crustacea: Copepoda) in Europe, the Urals and Western Siberia. Diversity 2024, 16, 47. https://doi.org/10.3390/d16010047
Sukhikh N, Garibian P, Chertoprud E. Resettlement of Eurytemora velox (Crustacea: Copepoda) in Europe, the Urals and Western Siberia. Diversity. 2024; 16(1):47. https://doi.org/10.3390/d16010047
Chicago/Turabian StyleSukhikh, Natalia, Petr Garibian, and Elena Chertoprud. 2024. "Resettlement of Eurytemora velox (Crustacea: Copepoda) in Europe, the Urals and Western Siberia" Diversity 16, no. 1: 47. https://doi.org/10.3390/d16010047
APA StyleSukhikh, N., Garibian, P., & Chertoprud, E. (2024). Resettlement of Eurytemora velox (Crustacea: Copepoda) in Europe, the Urals and Western Siberia. Diversity, 16(1), 47. https://doi.org/10.3390/d16010047