
First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea




Abstract
:Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandez-Leborans, G. Epibiosis in Crustacea: An overview. Crustaceana 2010, 83, 549–640. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit crab biocoenoses: A worldwide review of the diversity and natural history of hermit crab associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- Bouillon, J.; Gravili, C.; Pagès, F.; Gili, J.-M.; Boero, F. An introduction to hydrozoa. In Mémoires du Muséum National d’Histoire Naturelle; MarBEF: Aberdeen, Scotland, 2006; Volume 194, pp. 1–598. [Google Scholar]
- Antsulevich, A.E. Hydrozoa (Hydroids and Hydromedusae of Russian Seas); St. Petersburg University: St. Petersburg, Russia, 2015. (In Russian) [Google Scholar]
- Schuchert, P. Hydroids of Greenland (Cnidaria, Hydrozoa). Meddelelser om Gremland. Bioscience; Danish Polar Center: Copenhagen, Denmark, 2001; Volume 53. [Google Scholar]
- Schuchert, P. Survey of the family Corynidae (Cnidaria, Hydrozoa). Rev. Suisse Zool. 2001, 108, 739–878. [Google Scholar] [CrossRef]
- Schuchert, P. North-West European Athecate Hydroids and Their Medusae: Keys and Notes for the Identification of the Species; Dorset Press: Dorchester, UK, 2012. [Google Scholar]
- Rees, W.J. Evolutionary trends in the classification of capitate hydroids and medusae. Bull. Brit. Mus. Nat. Hist. Zool. 1957, 4, 453–534. [Google Scholar]
- Bouillon, J. Essai de classification des Hydropolypes−Hydroméduses (Hydrozoa-Cnidaria). Indo-Malayan Zool. 1985, 1, 29–243. [Google Scholar]
- Rees, J.T. The polyp and immature medusa stages of Cladonema californicum, Hyman, 1947 (Anthomedusae: Cladonemidae) with biological notes and a discussion of the taxonomy of the genus Cladonema. J. Nat. Hist. 1979, 13, 295–302. [Google Scholar] [CrossRef]
- Gili, J.M.; Lopez-Gonzalez, P.J.; Bouillon, J. A new Antarctic association: The case of the hydroid Sarsia medelae (new sp.) associated with gorgonians. Polar Biol. 2006, 29, 624–631. [Google Scholar] [CrossRef]
- Boero, F.; Gravili, C.; Denitto, F.; Miglietta, M.P.; Bouillon, J. The rediscovery of Codonorchis octaedrus (Hydroidomedusae, Anthomedusae, Pandeidae), with an update of the Mediterranean hydroidomedusan biodiversity. Ital. J. Zool. 1997, 64, 359–365. [Google Scholar] [CrossRef]
- Mills, C.E.; Marques, A.C.; Migotto, A.E.; Calder, D.R.; Hand, C.; Rees, J.T.S.; Haddock, H.D.C.; Dunn, W.; Pugh, P.R. Hydrozoa: Polyps, Hydromedusae, and Siphonophora. In The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon; Carlton, J.T., Ed.; University of California Press: Berkeley, CA, USA, 2007; pp. 118–167. [Google Scholar]
- Matishov, G.G. (Ed.) Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; KSC RAS: Apatity, Russia, 2008. (In Russian) [Google Scholar]
- Kuznetsov, V.V. The Biology of Abundant and the Most Common Species of Crustaceans in the Barents and White Seas; Nauka: Moscow, USSR, 1964. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Stevens, B.G. (Ed.) King Crabs of the World: Biology and Fisheries Management; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Panteleeva, N.N.; Dvoretsky, A.G.; Dvoretsky, V.G. New Records of the hydrozoan Coryne hincksi Bonnevie, 1898 on red king crabs in the Barents Sea. Diversity 2023, 15, 100. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef] [PubMed]
- Naumov, D.V. Hydroids and Hydromedusae of Marine, Brackish and Freshwater Basins of the USSR. In Guide to the Fauna of the USSR; Academy of Sciences: Moscow–Leningrad, USSR, 1960. (In Russian) [Google Scholar]
- Arai, M.N. Behavior of the planktonic coelenterates, Sarsia tubulosa, Phialidium gregarium, and Pleurobrachia pileus in salinity discontinuity layers. J. Fish. Board Can. 1973, 30, 1105–1110. [Google Scholar] [CrossRef]
- Singla, C.L.; Weber, C. Fine structure of the ocellus of Sarsia tubulosa (Hydrozoa, Anthomedusae). Zoomorphology 1982, 100, 11–22. [Google Scholar] [CrossRef]
- Leonard, J.L. The effect of environmental factors on swimming activity in Sarsia tubulosa M. Sars (Hydrozoa). Mar. Freshw. Behav. Physiol. 1983, 9, 99–110. [Google Scholar] [CrossRef]
- Daan, R. Food intake and growth of Sarsia tubulosa (Sars, 1835), with quantitative estimates of predation on copepod populations. Neth. J. Sea Res. 1986, 20, 67–74. [Google Scholar] [CrossRef]
- Sahin, M.; Mohseni, K.; Colin, S.P. The numerical comparison of flow patterns and propulsive performances for the hydromedusae Sarsia tubulosa and Aequorea victoria. J. Exp. Biol. 2009, 212, 2656–2667. [Google Scholar] [CrossRef]
- Edwards, C. The hydroids and medusae Sarsia occulta sp.nov., Sarsia tubulosa and Sarsia loveni. J. Mar. Biol. Assoc. UK 1978, 58, 291–311. [Google Scholar] [CrossRef]
- Bouillon, J.; Medel, M.D.; Pagès, F.; Gili, J.M.; Boero, F.; Gravili, C. Fauna of the Mediterranean hydrozoa. Sci. Mar. 2004, 68 (Suppl. S2), 5–438. [Google Scholar] [CrossRef]
- Il’in, G.V.; Moiseev, D.V.; Shirokolobov, D.V.; Deryabin, A.A.; Pavlova, L.G. Long-term dynamics of hydrological conditions of the Zelenetskaya Bay, East Murman. Vestn. MSTU 2016, 19, 268–277. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks; Alaska Sea Grant College Program; University of Alaska: Fairbanks, AL, USA, 2005. [Google Scholar]





{kind=link}
{kind=link}
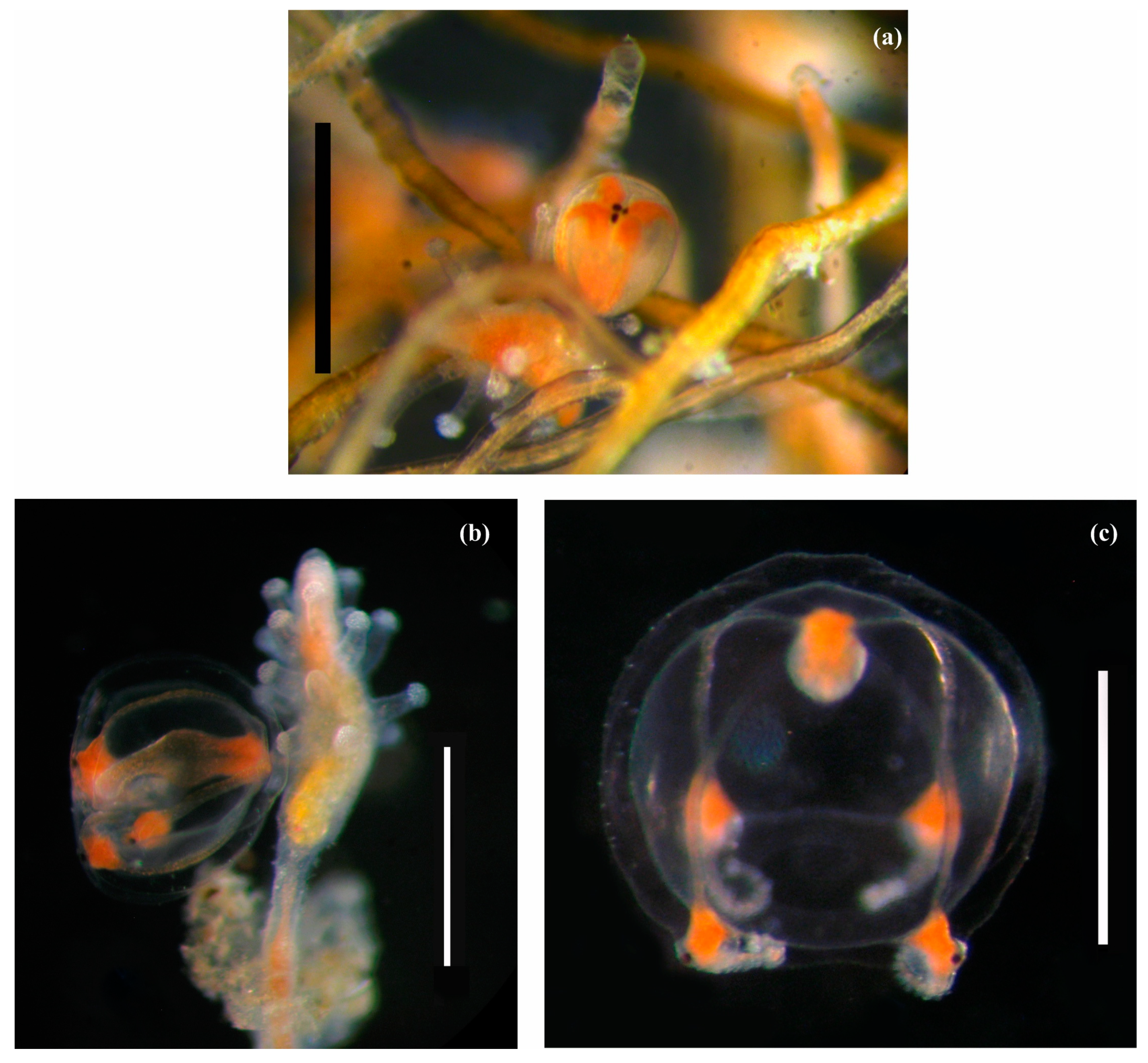{kind=link}
{kind=link}
| No. | Date | Latitude, N | Longitude, E | Depth | T, °C | CL, mm | Wet Weight, g | Shell Condition | Localization |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 6 July 2022 | 69°07′10″ | 36°05′42″ | 18 | 4.0 | 156.0 | 3262 | Very old | Limbs |
| 2 | 12 July 2022 | 69°07′07″ | 36°04′24″ | 31 | 5.0 | 175.0 | 4676 | Old | Carapace |
| 3 | 13 July 2022 | 69°07′30″ | 36°04′48″ | 16 | 6.0 | 156.3 | 2643 | Old | Abdomen |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panteleeva, N.N.; Dvoretsky, A.G.; Dvoretsky, V.G. First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea. Diversity 2024, 16, 72. https://doi.org/10.3390/d16010072
Panteleeva NN, Dvoretsky AG, Dvoretsky VG. First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea. Diversity. 2024; 16(1):72. https://doi.org/10.3390/d16010072
Chicago/Turabian StylePanteleeva, Ninel N., Alexander G. Dvoretsky, and Vladimir G. Dvoretsky. 2024. "First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea" Diversity 16, no. 1: 72. https://doi.org/10.3390/d16010072
APA StylePanteleeva, N. N., Dvoretsky, A. G., & Dvoretsky, V. G. (2024). First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea. Diversity, 16(1), 72. https://doi.org/10.3390/d16010072