
Molecular Diversity of the Genus Plagiorchis Lühe, 1899 in Snail Hosts of Central Europe with Evidence of New Lineages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Material Processing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site/Country | Coordinates | Sampling Period | Snail Species | Sampled Snails | Infected Snails | Plagiorchis Infected Snails | Plagiorchis Prevalence |
|---|---|---|---|---|---|---|---|
| Barbora | 50°38′35.6″ N, | 2021–2023 | Radix auricularia | 325 | 46 | 8 | 2.5% |
| CZ | 13°45′00.1″ E | Galba truncatula | 1 | — | — | — | |
| Stagnicola sp. | 2 | — | — | — | |||
| Medard | 50°10′44.0″ N, | 2016–2023 | Ampullaceana balthica 1 | 7995 | 877 | 141 | 1.8% |
| CZ | 12°35′50.1″ E | Lymnaea stagnalis | 251 | 72 | 5 | 2.0% | |
| Milada | 50°39′15.2″ N, | 2016–2021 | Ampullaceana lagotis 2 | 279 | 58 | 11 | 3.9% |
| CZ | 13°57′01.2″ E | Lymnaea stagnalis | 102 | 77 | 3 | 2.9% | |
| Galba truncatula | 11 | — | — | — | |||
| Most | 50°32′13.6″ N, | 2016–2023 | Ampullaceana lagotis | 2200 | 927 | 33 | 1.5% |
| CZ | 13°38′40.7″ E | Radix auricularia | 87 | 73 | — | — | |
| Otakar | 50°39′04.8″ N, | 2021–2023 | Radix auricularia | 438 | 80 | 2 | 0.5% |
| CZ | 13°44′23.1″ E | Galba truncatula | 16 | — | — | — | |
| Peregriana peregra | 1 | 1 | — | — | |||
| Stagnicola sp. | 3 | 2 | — | — | |||
| Písník Dubina | 50°11′20.8″ N, | 2023 | Radix auricularia | 326 | 99 | 3 | 0.9% |
| CZ | 15°47′09.7″ E | ||||||
| Skulska Wieś | 52°29′34.7″ N, | 2023 | Ampullaceana balthica 3 | 101 | 13 | 1 | 1.0% |
| PL | 18°18′57.5″ E | ||||||
| Spůle | 49°21′24.3″ N, | 2023 | Radix auricularia | 47 | 5 | 4 | 8.5% |
| CZ | 13°11′44.2″ E | ||||||
| Total | 12,185 | 2330 | 211 | 1.7% |
2.2. Sequence Generation
2.3. Species Identification and Phylogenetic Analyses
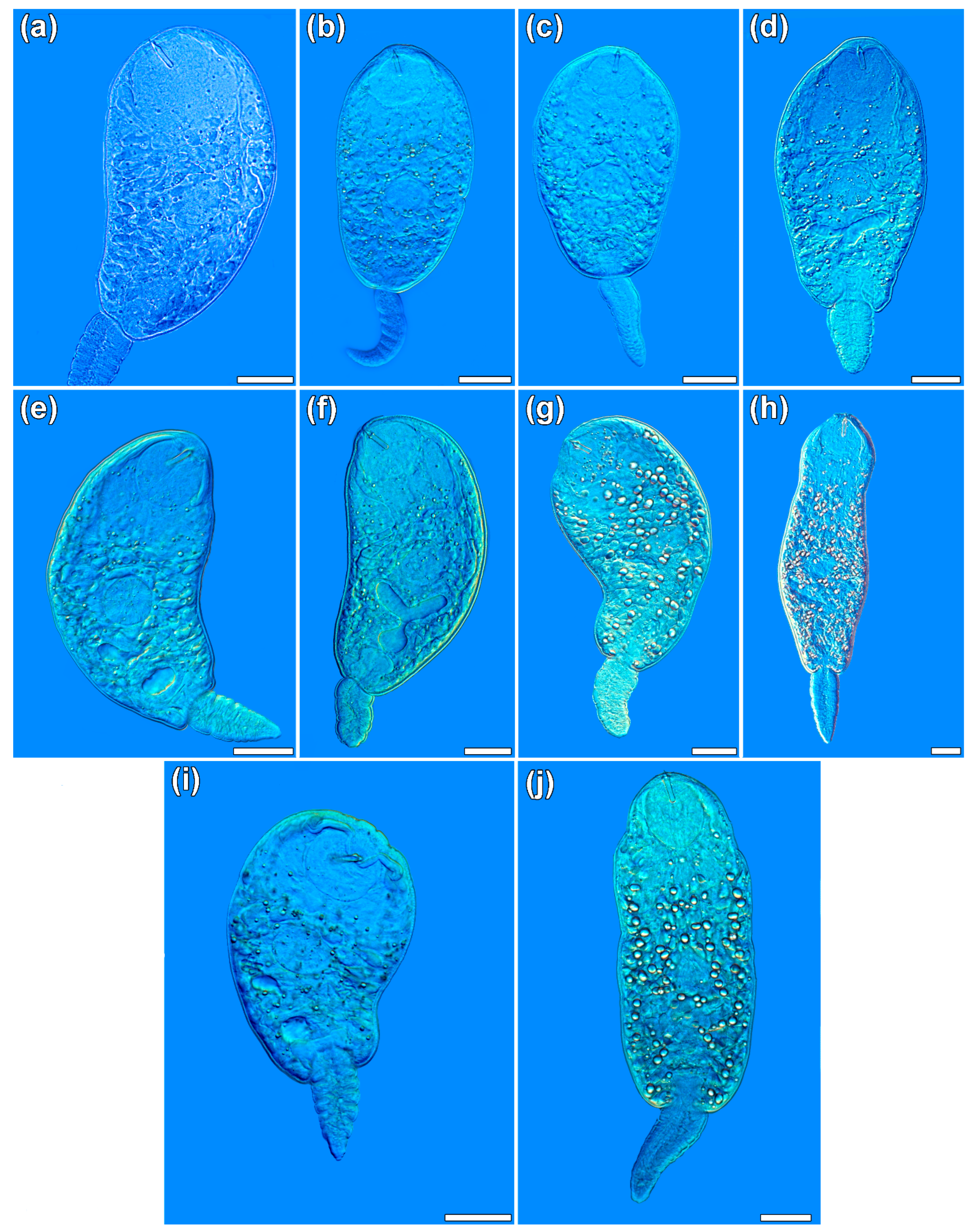
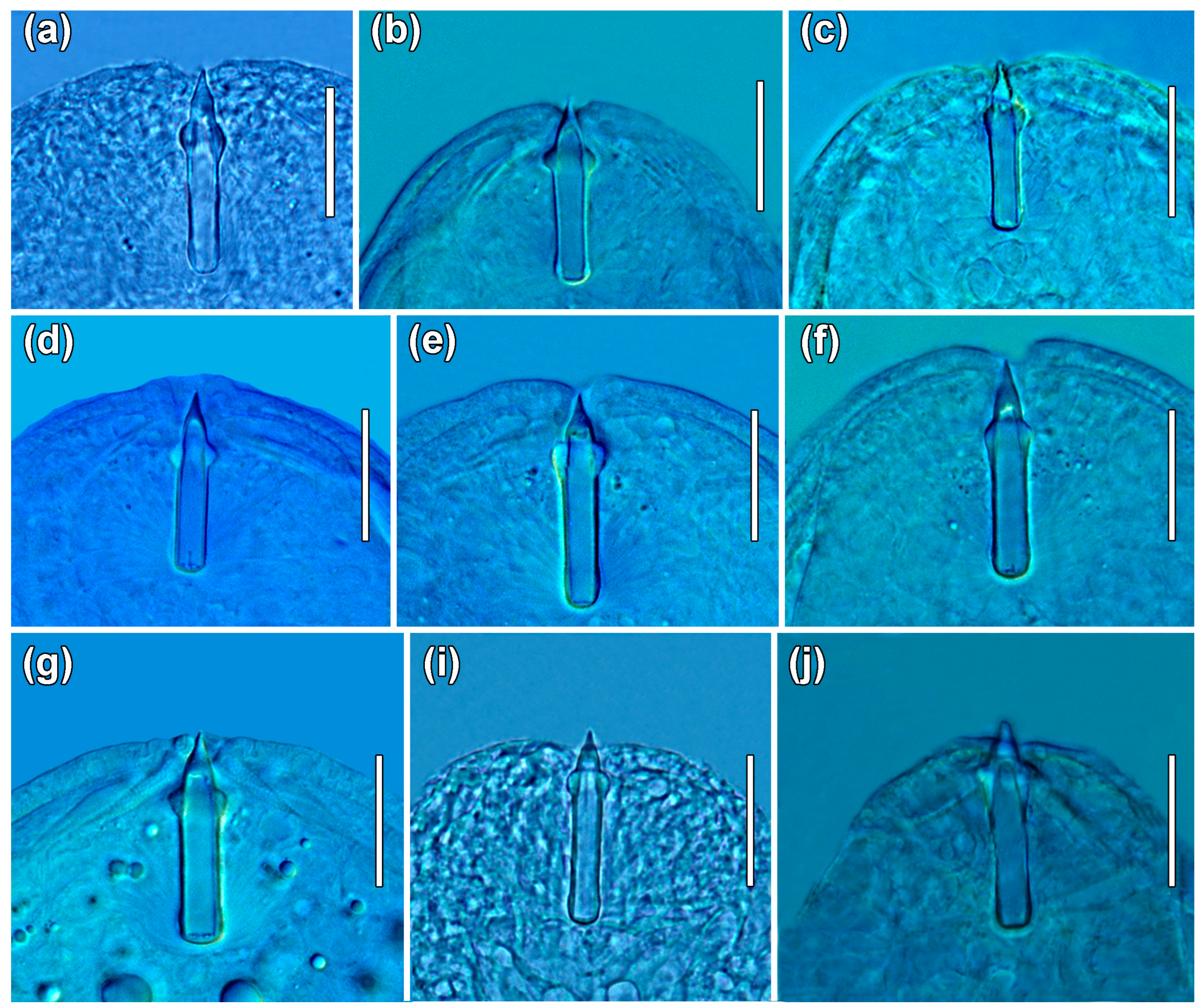
2.4. Morphometric Characterization
2.5. Analyses of Ecological Data
3. Results
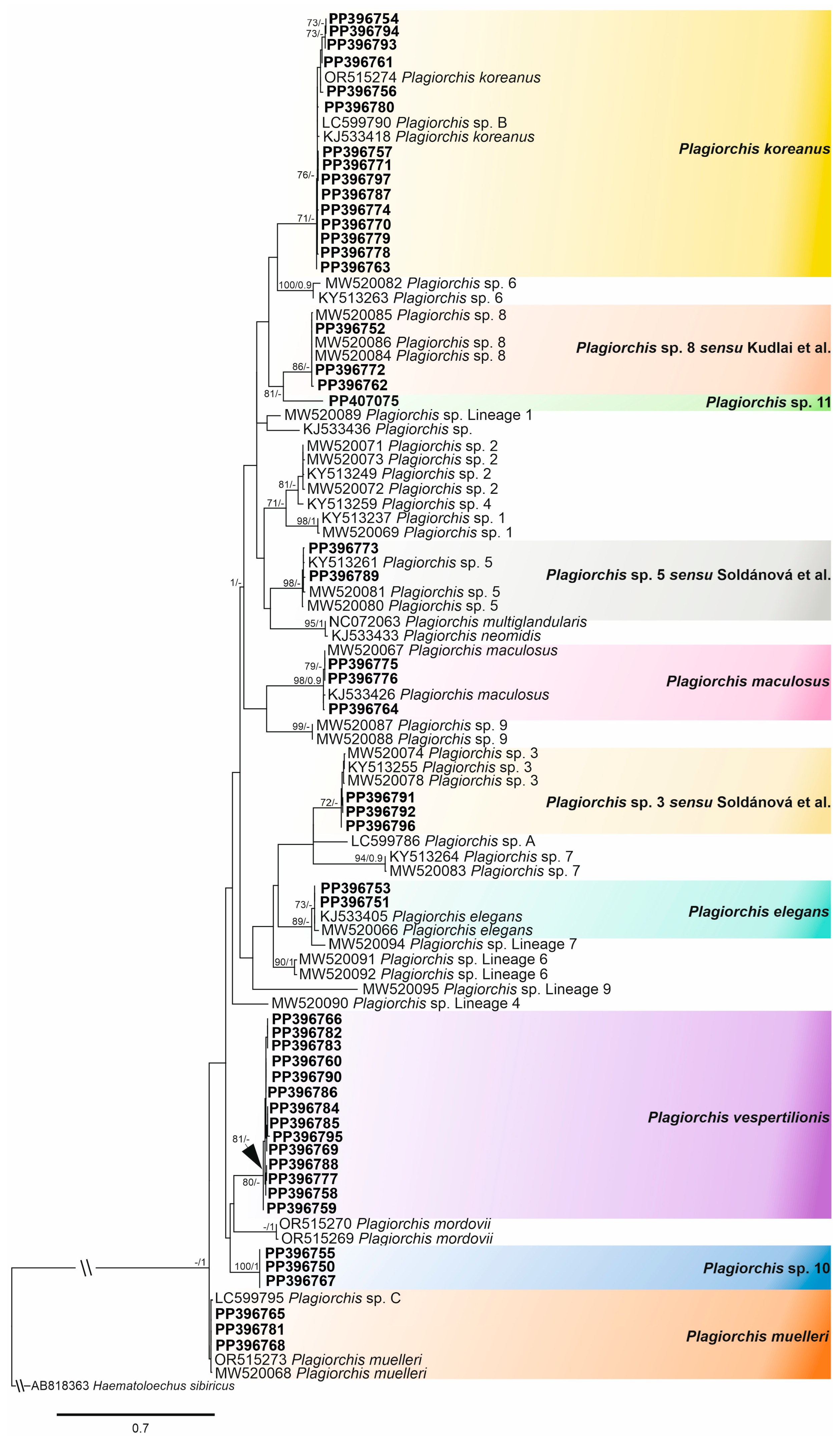
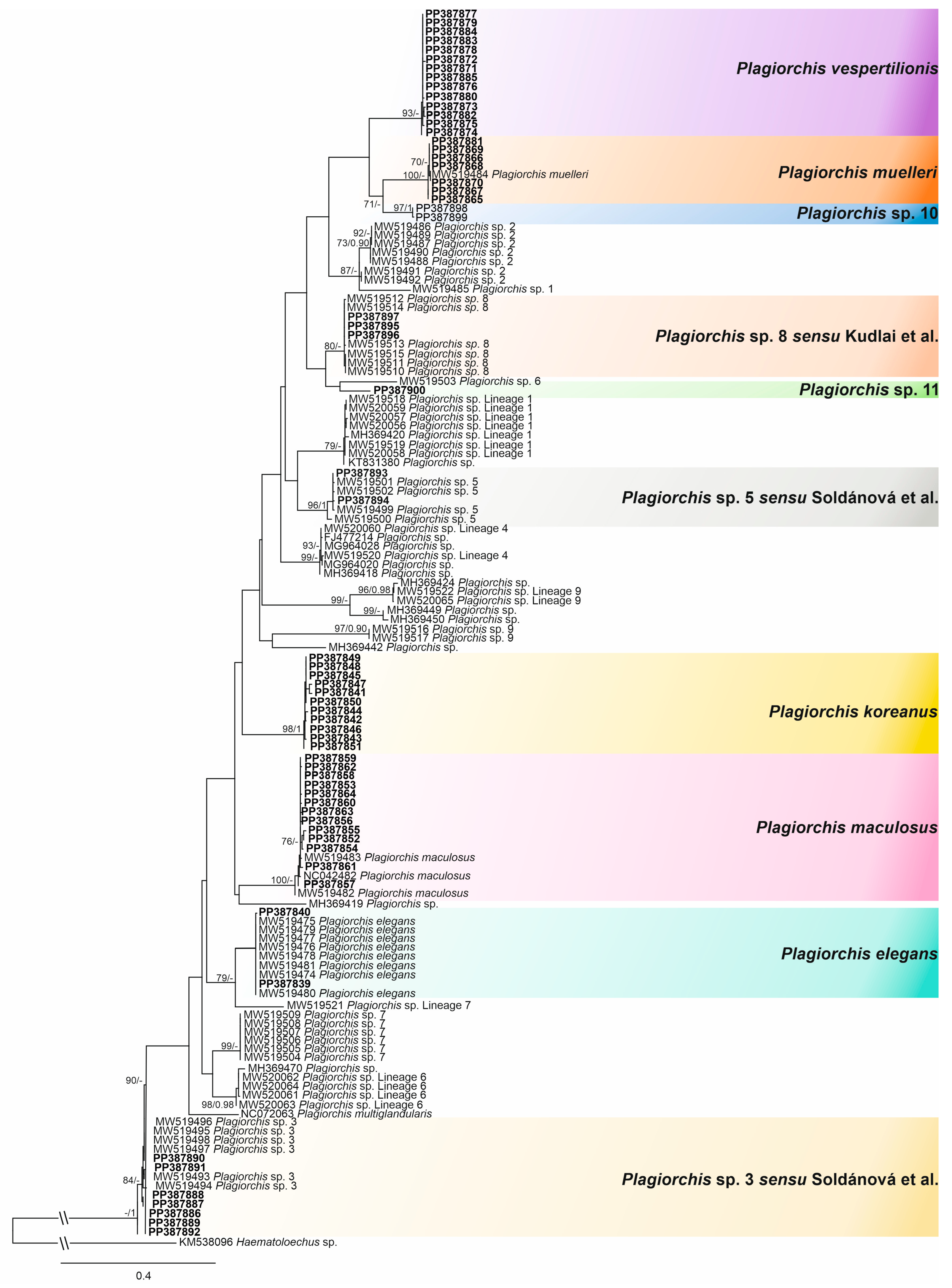
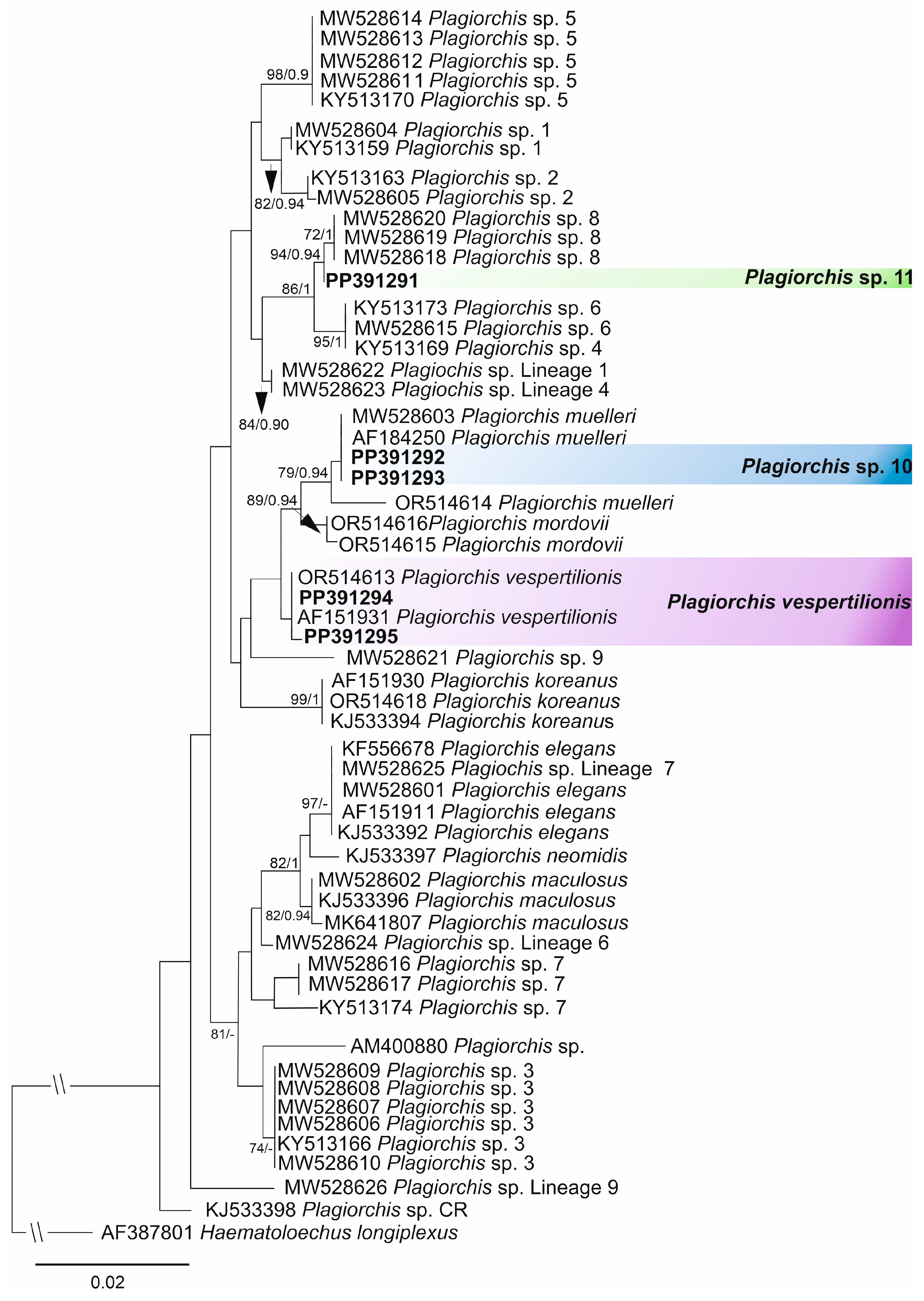
3.1. Sequence-Based Identification
3.2. Morphometric Characterization
- Plagiorchis vespertilionis (Müller, 1780)
- First intermediate hosts: Ampullaceana balthica, Ampullaceana lagotis, Radix auricularia.
- Localities: Lakes Barbora, Medard, Most, Otakar.
- Plagiorchis sp. 10
- First intermediate hosts: Ampullaceana balthica, Ampullaceana lagotis.
- Localities: Lakes Medard, Most, Skulska Wieś.
- Plagiorchis sp. 11
- First intermediate host: Ampullaceana lagotis.
- Locality: Lake Most.
3.3. Prevalence, Distribution and Diversity of Plagiorchis Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rands, M.R.; Adams, W.M.; Bennun, L.; Butchart, S.H.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Ali, A. Biodiversity-ecosystem functioning research: Brief history, major trends and perspectives. Biol. Conserv. 2023, 285, 110210. [Google Scholar] [CrossRef]
- Beck, H.P.; Blake, D.; Dardé, M.L.; Felger, I.; Pedraza-Díaz, S.; Regidor-Cerrillo, J.; Gómez-Bautista, M.; Ortega-Mora, L.M.; Putignani, L.; Shiels, B.; et al. Molecular approaches to diversity of populations of apicomplexan parasites. Int. J. Parasitol. 2009, 39, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Lacorte, G.A.; Felix, G.M.; Pinheiro, R.R.; Chaves, A.V.; Almeida-Neto, G.; Neves, F.S.; Leite, L.O.; Santos, F.R.; Braga, E.M. Exploring the diversity and distribution of neotropical avian malaria parasites—A molecular survey from Southeast Brazil. PLoS ONE 2013, 8, e57770. [Google Scholar] [CrossRef] [PubMed]
- Morand, S.; Krasnov, B.R.; Littlewood, D.T.J. Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics; Cambridge University Press: Cambridge, UK, 2015; ISBN 110-703-765-4. [Google Scholar] [CrossRef]
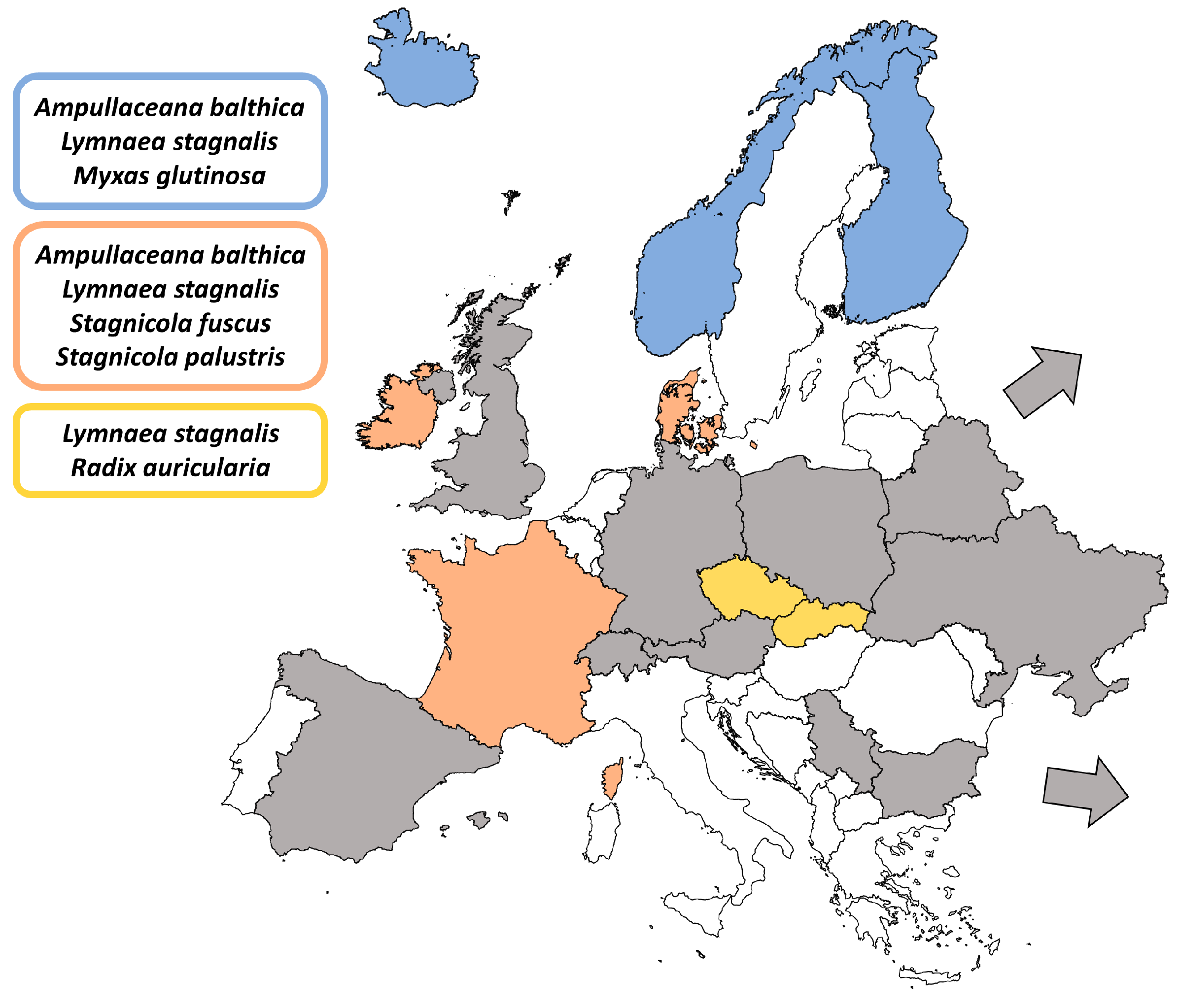
- Kudlai, O.; Pantoja, C.; O’Dwyer, K.; Jouet, D.; Skírnisson, K.; Faltýnková, A. Diversity of Plagiorchis (Trematoda: Digenea) in high latitudes: Species composition and snail host spectrum revealed by integrative taxonomy. J. Zoolog. Syst. Evol. Res. 2021, 59, 937–962. [Google Scholar] [CrossRef]
- Pantoja, C.; Faltýnková, A.; O’Dwyer, K.; Jouet, D.; Skírnisson, K.; Kudlai, O. Diversity of echinostomes (Digenea: Echinostomatidae) in their snail hosts at high latitudes. Parasite 2021, 28, 59. [Google Scholar] [CrossRef]
- Scholz, T.; Kuchta, R. Fish tapeworms (Cestoda) in the molecular era: Achievements, gaps and prospects. Parasitology 2022, 149, 1876–1893. [Google Scholar] [CrossRef]
- Cháves-González, L.E.; Morales-Calvo, F.; Mora, J.; Solano-Barquero, A.; Verocai, G.G.; Rojas, A. What lies behind the curtain: Cryptic diversity in helminth parasites of human and veterinary importance. Curr. Res. Parasitol. Vector. Borne Dis. 2022, 2, 100094. [Google Scholar] [CrossRef]
- MacKenzie, K. Parasites as indicators of host populations. Int. J. Parasitol. 1987, 17, 345–352. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Taylor, C.M. Macroecology of a host-parasite relationship. Ecography 2000, 23, 11–20. [Google Scholar] [CrossRef]
- Krasnov, B.R.; Shenbrot, G.I.; Khokhlova, I.S.; Allan Degen, A. Relationship between host diversity and parasite diversity: Flea assemblages on small mammals. J. Biogeogr. 2004, 31, 1857–1866. [Google Scholar] [CrossRef]
- Hechinger, R.F.; Lafferty, K.D. Host diversity begets parasite diversity: Bird final hosts and trematodes in snail intermediate hosts. Proc. R. Soc. B 2005, 272, 1059–1066. [Google Scholar] [CrossRef]
- Hechinger, R.F.; Lafferty, K.D.; Huspeni, T.C.; Brooks, A.J.; Kuris, A.M. Can parasites be indicators of free-living diversity? Relationships between species richness and the abundance of larval trematodes and of local benthos and fishes. Oecologia 2007, 151, 82–92. [Google Scholar] [CrossRef]
- Hechinger, R.F.; Lafferty, K.D.; Kuris, A.M. Trematodes indicate animal biodiversity in the Chilean intertidal and Lake Tanganyika. J. Parasitol. 2008, 94, 966–968. [Google Scholar] [CrossRef]
- Kuris, A.M.; Hechinger, R.F.; Shaw, J.C.; Whitney, K.L.; Aguirre-Macedo, L.; Boch, C.A.; Dobson, A.P.; Dunham, E.J.; Fredensborg, B.L.; Huspeni, T.C.; et al. Ecosystem energetic implications of parasite and free-living biomass in three estuaries. Nature 2008, 454, 515–518. [Google Scholar] [CrossRef]
- Soldánová, M.; Georgieva, S.; Roháčová, J.; Knudsen, R.; Kuhn, J.A.; Henriksen, E.H.; Siwertsson, A.; Shaw, J.C.; Kuris, A.M.; Amundsen, P.-A.; et al. Molecular analyses reveal high species diversity of trematodes in a sub-Arctic lake. Int. J. Parasitol. 2017, 47, 327–345. [Google Scholar] [CrossRef]
- Duan, Y.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Trematode diversity reflecting the community structure of Danish freshwater systems: Molecular clues. Parasit. Vectors 2021, 14, 43. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Gómez, A.; Nichols, E.S.; Perkins, S.L. Parasite conservation, conservation medicine, and ecosystem health. In New Directions in Conservation Medicine: Applied Cases of Ecological Health; Aguirre, A.A., Ostfeld, R., Daszak, P., Eds.; Oxford University Press: Oxford, UK, 2012; pp. 67–81. ISBN 978-019-973-147-3. [Google Scholar]
- Gagne, R.B.; Crooks, K.; Craft, M.E.; Chiu, E.S.; Fountain-Jones, N.M.; Malmberg, J.L.; Carver, S.; Funk, C.; VandeWoude, S. Parasites as conservation tools. Conserv. Biol. 2021, 36, e13719. [Google Scholar] [CrossRef]
- Poulin, R.; Morand, S. Parasite Biodiversity; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2005; ISBN 158-834-170-4. [Google Scholar]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How many parasites? How many hosts? Proc. Natl. Acad. Sci. USA 2008, 105, 11482–11489. [Google Scholar] [CrossRef]
- Carlson, C.J.; Dallas, T.A.; Alexander, L.W.; Phelan, A.L.; Phillips, A.J. What would it take to describe the global diversity of parasites? Proc. R. Soc. B 2020, 287, 20201841. [Google Scholar] [CrossRef]
- Bray, R.A.; Gibson, D.I.; Jones, A. Keys to the Trematoda, Volume 3; CABI: Wallingford, UK, 2008; ISBN 978-0-85199-588-5. [Google Scholar] [CrossRef]
- Nadler, S.A.; Pérez-Ponce de León, G. Integrating molecular and morphological approaches for characterizing parasite cryptic species: Implications for parasitology. Parasitology 2011, 138, 1688–1709. [Google Scholar] [CrossRef]
- Poulin, R. Uneven distribution of cryptic diversity among higher taxa of parasitic worms. Biol. Lett. 2011, 7, 241–244. [Google Scholar] [CrossRef]
- Georgieva, S.; Faltýnková, A.; Brown, R.; Blasco-Costa, I.; Soldánová, M.; Sitko, J.; Scholz, T.; Kostadinova, A. Echinostoma ‘revolutum’ (Digenea: Echinostomatidae) species complex revisited: Species delimitation based on novel molecular and morphological data gathered in Europe. Parasit. Vectors 2014, 7, 520. [Google Scholar] [CrossRef]
- Selbach, C.; Soldánová, M.; Georgieva, S.; Kostadinova, A.; Sures, B. Integrative taxonomic approach to the cryptic diversity of Diplostomum spp. in lymnaeid snails from Europe with a focus on the ‘Diplostomum mergi’ species complex. Parasit. Vectors 2015, 8, 300. [Google Scholar] [CrossRef]
- Pérez-Ponce de León, G.; Poulin, R. An updated look at the uneven distribution of cryptic diversity among parasitic helminths. J. Helminthol. 2018, 92, 197–202. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Poulin, R. Parasite life-cycle studies: A plea to resurrect an old parasitological tradition. J. Helminthol. 2017, 91, 647–656. [Google Scholar] [CrossRef]
- EUR-Lex. Available online: https://eur-lex.europa.eu/eli/dir/2010/63/oj (accessed on 21 December 2023).
- Tkach, V.V.; Pawlowski, J.; Sharpilo, V.P. Molecular and morphological differentiation between species of the Plagiorchis vespertilionis group (Digenea, Plagiorchiidae) occurring in European bats, with a re-description of P. vespertilionis (Müller, 1780). Syst. Parasitol. 2000, 47, 9–22. [Google Scholar] [CrossRef]
- Tkach, V.V. Family Plagiorchiidae Lühe, 1901. In Keys to the Trematoda, Volume 3; Bray, R.A., Gibson, D.I., Jones, A., Eds.; CABI: Wallingford, DC, UK, 2008; pp. 295–325. [Google Scholar] [CrossRef]
- Greani, S.; Quilichini, Y.; Foata, J.; Greiman, S.E.; Ndiaye, P.I.; Tkach, V.V.; Marchand, B. Vitellogenesis of the digenean Plagiorchis elegans (Rudolphi, 1802) (Plagiorchioidea, Plagiorchiidae). Parasitol. Int. 2014, 63, 537–543. [Google Scholar] [CrossRef]
- Horsák, M.; Juřičková, L.; Beran, L.; Čejka, T.; Dvořák, L. Annotated list of mollusc species recorded outdoors in the Czech and Slovak Republics. Malacol. Bohemoslov. 2010, 1, 1–37. (In Czech) [Google Scholar] [CrossRef]
- Aksenova, O.V.; Bolotov, I.N.; Gofarov, M.Y.; Kondakov, A.V.; Vinarski, M.V.; Bespalaya, Y.V.; Kolosova, Y.S.; Palatov, D.M.; Sokolova, S.E.; Spitsyn, V.M.; et al. Species richness, molecular taxonomy and biogeography of the Radicine pond snails (Gastropoda: Lymnaeidae) in the Old World. Sci. Rep. 2018, 8, 11199. [Google Scholar] [CrossRef]
- Cichy, A.; Faltýnková, A.; Żbikowska, E. Cercariae (Trematoda, Digenea) in European freshwater snails—A checklist of records from over one hundred years. Folia Malacol. 2011, 19, 165–189. [Google Scholar] [CrossRef]
- Faltýnková, A.; Sures, B.; Kostadinova, A. Biodiversity of trematodes in their intermediate mollusc and fish hosts in the freshwater ecosystems of Europe. Syst. Parasitol. 2016, 93, 283–293. [Google Scholar] [CrossRef]
- Zikmundová, J.; Georgieva, S.; Faltýnková, A.; Soldánová, M.; Kostadinova, A. Species diversity of Plagiorchis Lühe, 1899 (Digenea: Plagiorchiidae) in lymnaeid snails from freshwater ecosystems in central Europe revealed by molecules and morphology. Syst. Parasitol. 2014, 88, 37–54. [Google Scholar] [CrossRef]
- Krasnolobova, T.A. Trematodes of the Fauna of USSR. Genus Plagiorchis; Nauka: Moscow, Russia, 1987. (In Russian) [Google Scholar]
- Guk, S.M.; Kim, J.L.; Park, J.H.; Chai, J.Y. A human case of Plagiorchis vespertilionis (Digenea: Plagiorchiidae) infection in the Republic of Korea. J. Parasitol. 2007, 93, 1225–1227. [Google Scholar] [CrossRef]
- Boyce, K.; Hide, G.; Craig, P.S.; Reynolds, C.; Hussain, M.; Bodell, A.J.; Bradshaw, H.; Pickles, A.; Rogan, M.T. A molecular and ecological analysis of the trematode Plagiorchis elegans in the wood mouse Apodemus sylvaticus from a periaquatic ecosystem in the UK. J. Helminthol. 2014, 88, 310–320. [Google Scholar] [CrossRef]
- Chai, J.Y. Plagiorchids. In Human Intestinal Flukes; Springer: Dordrecht, The Netherlands, 2019; pp. 463–489. ISBN 978-94-024-1702-9. [Google Scholar]
- Suleman, M.J.; Khan, M.S.; Tkach, V.V.; Muhammad, N.; Zhang, D.; Zhu, X.Q. Characterization of the complete mitochondrial genome of Plagiorchis maculosus (Digenea, Plagiorchiidae) representative of a taxonomically complex digenean family. Parasitol. Int. 2019, 71, 99–105. [Google Scholar] [CrossRef]
- Našincová, V. Trematode Developmental Stages in Czech Aquatic Snails and Life-Cycles of Selected Species of the Family Omphalometridae and Echinostomatidae. Ph.D. Thesis, Institute of Parasitology, Czechoslovak Academy of Sciences, České Budějovice, Czech Republic, 1992. (In Czech). [Google Scholar]
- Faltýnková, A.; Našincová, V.; Kablásková, L. Larval trematodes (Digenea) of the great pond snail, Lymnaea stagnalis (L.), (Gastropoda, Pulmonata) in Central Europe: A survey of species and key to their identification. Parasite 2007, 14, 39–51. [Google Scholar] [CrossRef]
- Kirillova, N.Y.; Kirillov, A.A.; Shchenkov, S.V.; Knyazev, A.E.; Vekhnik, V.A. Morphological and molecular characterization of plagiorchiid trematodes (Plagiorchis: Plagiorchiidae, Digenea) from bats with redescription of Plagiorchis mordovii Shaldybin, 1958. J. Helminthol. 2024, 98, e2. [Google Scholar] [CrossRef]
- Pinto, H.A. Describing formally larval trematodes: Some reflections in the taxonomic integrative era. Trends Parasitol. 2023, 39, 889–890. [Google Scholar] [CrossRef]
- Gibson, D.I.; Bray, R.A.; Harris, E.A. (Compiler) Host-Fresent Study Are Highlighted in bMuseum. 2005. Available online: https://www.nhm.ac.uk/research-curation/scientific-resources/taxonomy-systematics/host-parasites/index.html (accessed on 22 January 2024).
- Huguenin, A.; Depaquit, J.; Villena, I.; Ferté, H. MALDI-TOF mass spectrometry: A new tool for rapid identification of cercariae (Trematoda, Digenea). Parasite 2019, 26, 11. [Google Scholar] [CrossRef]
- Li, R.; Wang, H.M.; Liu, G.H.; Tu, Y.; Deng, Y.P. Characterization of the complete mitochondrial genome of the fluke of turdus, Plagiorchis elegans, and phylogenetic implications. Exp. Parasitol. 2022, 242, 108387. [Google Scholar] [CrossRef]
- Gacad, J.L.J.; Yurlova, N.I.; Ponomareva, N.M.; Urabe, M. Characterization of the complete mitochondrial genome of Plagiorchis multiglandularis (Digenea, Plagiorchiidae): Comparison with the members of Xiphidiatan species and phylogenetic implications. Parasitol. Res. 2023, 122, 1545–1556. [Google Scholar] [CrossRef]
- Soldánová, M.; Selbach, C.; Sures, B.; Kostadinova, A.; Pérez-del-Olmo, A. Larval trematode communities in Radix auricularia and Lymnaea stagnalis in a reservoir system of the Ruhr River. Parasit. Vectors 2010, 3, 56. [Google Scholar] [CrossRef]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10, 2694. [Google Scholar] [CrossRef]
- Żbikowska, E.; Nowak, A. One hundred years of research on the natural infection of freshwater snails by trematode larvae in Europe. Parasitol. Res. 2009, 105, 301–311. [Google Scholar] [CrossRef]
- Přikryl, I.; Kabrna, M. Findings from flooding residual pits remaining after coal mining in the Czech Republic. In Mining Meets Water—Conflicts and Solutions, Proceedings of the International Mine Water Association Symposium, Leipzig, Germany, 11–15 July 2016; Drebenstedt, C., Paul, M., Eds.; Technische Universität Bergakademie: Freiberg, Germany, 2016; pp. 201–208. [Google Scholar]
- Žižka, L.; Burda, J. Lake Most. In Post Exploitation Lakes, Risk Assessment of Final pits during Flooding; Burda, J., Bajcar, A., Eds.; Zpravodaj Hnědé Uhlí: Most, Czech Republic, 2020; pp. 11–46. [Google Scholar]
- Žižka, L.; Valvoda, P.; Burda, J. Lake Medard. In Post Exploitation Lakes, Risk Assessment of Final pits during Flooding; Burda, J., Bajcar, A., Eds.; Zpravodaj Hnědé Uhlí: Most, Czech Republic, 2020; pp. 47–59. [Google Scholar]
- Bažant, J. Waterbirds on lake Most 2013–2015. Sbor. Obl. Muz. Most. ř. př. 2015, 37, 61–80. (In Czech) [Google Scholar]
- Bažant, J. Interesting ornithological observations in the Most district (Northwestern Bohemia). Sbor. Obl. Muz. Most. ř. př. 2018, 39, 143–153. (In Czech) [Google Scholar]
- Bažant, J. Birds of the sandpit in Polerady village (Most county, Northwestern Bohemia). Sbor. Obl. Muz. Most. ř. př. 2020, 40, 122–134. (In Czech) [Google Scholar]
- Pozorování Ptáků. Available online: https://pozorovaniptaku.cz/ (accessed on 22 January 2024). (In Czech).
- Ptasie Wyspy 22. Available online: https://alauda.org.pl/ptasie-wyspy-22/obszary-objete-projektem/ (accessed on 8 January 2024). (In Polish).
- Glöer, P. Die Süßwassergastropoden Nord- und Mitteleuropas. Bestimmungschlüssel; Lebensweise, Verbreitung; ConchBooks: Harxheim, Germany, 2002; ISBN 392-591-960-0. [Google Scholar]
- Glöer, P. The Freshwater Gastropods of the West-Palaearctis, Volume 1: Fresh- and Brackish Waters except Spring and Subterranean Snails; Biodiversity Research Lab: Hetlingen, Germany, 2019. [Google Scholar]
- Schniebs, K.; Glöer, P.; Vinarski, M.V.; Hundsdoerfer, A.K. Intraspecific morphological and genetic variability in Radix balthica (Linnaeus 1758) (Gastropoda: Basommatophora: Lymnaeidae) with morphological comparison to other European Radix species. J. Conchol. 2011, 40, 657–677. [Google Scholar]
- Huňová, K.; Kašný, M.; Hampl, V.; Leontovyč, R.; Kuběna, A.; Mikeš, L.; Horák, P. Radix spp.: Identification of trematode intermediate hosts in the Czech Republic. Acta Parasitol. 2012, 57, 273–284. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D.P. A molecular phylogeny of the human schistosomes. Mol. Phylogenet. Evol. 1995, 4, 103–109. [Google Scholar] [CrossRef]
- Koehler, A.V.; Springer, Y.P.; Keeney, D.B.; Poulin, R. Intra-and interclonal phenotypic and genetic variability of the trematode Maritrema novaezealandensis. Biol. J. Linn. Soc. 2011, 103, 106–116. [Google Scholar] [CrossRef]
- Van Steenkiste, N.; Locke, S.A.; Castelin, M.; Marcogliese, D.J.; Abbott, C.L. New primers for DNA barcoding of digeneans and cestodes (Platyhelminthes). Mol. Ecol. Resour. 2015, 15, 945–952. [Google Scholar] [CrossRef]
- Tkach, V.V.; Littlewood, D.T.J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z.P. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Syst. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef]
- Almeyda-Artigas, R.J.; Bargues, M.D.; Mas-Coma, S. ITS-2 rDNA sequencing of Gnathostoma species (Nematoda) and elucidation of the species causing human gnathostomiasis in the Americas. J. Parasitol. 2000, 86, 537–544. [Google Scholar] [CrossRef]
- Bargues, M.D.; Vigo, M.; Horák, P.; Dvořák, J.; Patzner, R.A.; Pointier, J.P.; Jackiewicz, M.; Meier-Brook, C.; Mas-Coma, S. European Lymnaeidae (Mollusca: Gastropoda), intermediate hosts of trematodiases, based on nuclear ribosomal DNA ITS-2 sequences. Infect. Genet. Evol. 2001, 1, 85–107. [Google Scholar] [CrossRef]
- Littlewood, D.T.; Rohde, K.; Clough, K.A. Parasite speciation within or between host species? Phylogenetic evidence from site-specific polystome monogeneans. Int. J. Parasitol. 1997, 27, 1289–1297. [Google Scholar] [CrossRef]
- Littlewood, D.T.; Curini-Galletti, M.; Herniou, E.A. The interrelationships of Proseriata (Platyhelminthes: Seriata) tested with molecules and morphology. Mol. Phylogenet. Evol. 2000, 16, 449–466. [Google Scholar] [CrossRef]
- Garey, J.R.; Wolstenholme, D.R. Platyhelminth mitochondrial DNA: Evidence for early evolutionary origin of a tRNA ser AGN that contains a dihydrouridine arm replacement loop, and of serine-specifying AGA and AGG codons. J. Mol. Evol. 1989, 28, 374–387. [Google Scholar] [CrossRef]
- Ohama, T.; Osawa, S.; Watanabe, K.; Jukes, T.H. Evolution of the mitochondrial genetic code IV. AAA as an asparagine codon in some animal mitochondria. J. Mol. Evol. 1990, 30, 329–332. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.; Xie, W.; Drummond, A. Tracer v. 1.6. University of Edinburgh, Institute of Evolutionary Biology. Available online: https://beast.bio.ed.ac.uk/Tracer (accessed on 25 January 2024).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Gordy, M.A.; Hanington, P.C. A fine-scale phylogenetic assessment of digenean trematodes in central Alberta reveals we have yet to uncover their total diversity. Ecol. Evol. 2019, 9, 3153–3238. [Google Scholar] [CrossRef]
- Bušta, J.; Našincová, V. Record of Plagiorchis neomidis Brendow, 1970 (Trematoda: Plagiorchidae) in Czechoslovakia and studies on its life cycle. Folia Parasitol. 1986, 33, 123–129. [Google Scholar]
- Schwelm, J.; Selbach, C.; Kremers, J.; Sures, B. Rare inventory of trematode diversity in a protected natural reserve. Sci. Rep. 2021, 11, 22066. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Cutmore, S.C.; Miller, T.L.; Nolan, M.J. Molecular approaches to trematode systematics: ‘best practice’ and implications for future study. Syst. Parasitol. 2016, 93, 295–306. [Google Scholar] [CrossRef]
- Gordy, M.A.; Kish, L.; Tarrabain, M.; Hanington, P.C. A comprehensive survey of larval digenean trematodes and their snail hosts in central Alberta, Canada. Parasitol. Res. 2016, 115, 3867–3880. [Google Scholar] [CrossRef]
- Cramp, S.; Simmons, K.E.L. Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic (Vol. 3). Waders to Gulls; Oxford University Press: Oxford, UK, 1983; ISBN 978-0-19-857506-1. [Google Scholar]
- Smit, C.J.; Piersma, T. Numbers, midwinter distribution, and migration of wader populations using the East Atlantic flyway. In Flyways and Reserve Networks for Water Birds; Boyd, H., Pirot, J.-Y., Eds.; Canadian Wildlife Service: Ottawa, ON, Canada, 1989; pp. 24–63. ISBN 978-066-217-104-1. [Google Scholar]
- Bairlein, F.; Norris, D.R.; Nagel, R.; Bulte, M.; Voigt, C.C.; Fox, J.W.; Hussell, D.J.T.; Schmaljohann, H. Cross-hemisphere migration of a 25 g songbird. Biol. Lett. 2012, 8, 505–507. [Google Scholar] [CrossRef]
- Esch, G.W.; Fernandez, J.C. Snail-trematode interactions and parasite community dynamics in aquatic aystems: A review. Am. Midl. Nat. 1994, 131, 209–237. [Google Scholar] [CrossRef]
- Smith, N.F. Spatial heterogeneity in recruitment of larval trematodes to snail intermediate hosts. Oecologia 2001, 127, 115–122. [Google Scholar] [CrossRef]
- Byers, J.E.; Blakeslee, A.M.; Linder, E.; Cooper, A.B.; Maguire, T.J. Controls of spatial variation in the prevalence of trematode parasites infecting a marine snail. Ecology 2008, 89, 439–451. [Google Scholar] [CrossRef]
- Fredensborg, B.L.; Mouritsen, K.N.; Poulin, R. Relating bird host distribution and spatial heterogeneity in trematode infections in an intertidal snail—From small to large scale. Mar. Biol. 2006, 149, 275–283. [Google Scholar] [CrossRef]
- Denk, T.; Grímsson, F.; Zetter, R.; Símonarson, L.A. Introduction to the Nature and Geology of Iceland. In Late Cainozoic Floras of Iceland: 15 Million Years of Vegetation and Climate History in the Northern North Atlantic. Topics in Geobiology; Springer: Dordrecht, The Netherlands, 2011; Volume 35, pp. 1–29. ISBN 978-94-007-0371-1. [Google Scholar] [CrossRef]
- Frafjord, K. Influence of night length on home range size in the northern bat Eptesicus nilssonii. Mamm. Biol. 2013, 78, 205–211. [Google Scholar] [CrossRef]
- Petersen, A.; Jensen, J.K.; Jenkins, P.; Bloch, D.; Ingmarsson, F. A Review of the occurrence of bats (Chiroptera) on Islands in the North East Atlantic and on North Sea installations. Acta Chiropt. 2014, 19, 169–195. [Google Scholar] [CrossRef]
- Krasnolobova, T.A. Validity of the species Plagiorchis fastuosus Szidat, 1924 and its life-cycle. Trudy Gel’mintol. Labor. 1973, 23, 86–96. (In Russian) [Google Scholar]
- Schniebs, K.; Glöer, P.; Vinarski, M.V.; Hundsdoerfer, A.K. Intraspecific morphological and genetic variability in the European freshwater snail Radix labiata (Rossmaessler, 1835) (Gastropoda: Basommatophora: Lymnaeidae). Contrib. Zool. 2013, 82, 55–68. [Google Scholar] [CrossRef]
- Schniebs, K.; Georgiev, D.; Glöer, P.; Hundsdoerfer, A.K. A molecular genetic evidence of the occurrence of the freshwater snail Radix lagotis (Schrank, 1803) (Gastropoda: Lymnaeidae) in Bulgaria. Ecol. Monten. 2015, 3, 29–39. [Google Scholar] [CrossRef]
- Akimova, L.N.; Shimalov, V.V.; Bychkova, E.I. Species diversity of trematode larvae of gastropods in water bodies of Belarus. Parazitologiya 2011, 45, 287–305. (In Russian) [Google Scholar]
- Hanák, V.; Anděra, M.; Uhrin, M.; Danko, Š.; Horáček, I. Bats of the Czech Republic and Slovakia: Distributional status of individual species. In A Tribute to Bats; Horáček, I., Uhrin, M., Eds.; Lesnická práce: Kostelec nad Černými lesy, Czech Republic, 2010; pp. 143–254. ISBN 978-80-87154-44-1. [Google Scholar]
- Ancillotto, L.; Bosso, L.; Salinas-Ramos, V.B.; Russo, D. The importance of ponds for the conservation of bats in urban landscapes. Landsc. Urban Plan. 2019, 190, 103607. [Google Scholar] [CrossRef]
- Pešout, P.; Porteš, M.; Pixová, K.Č.; Hendrychová, M.; Kříž, P.; Lacina, D. Ecosystem restoration of brown coal open-pit mines. Nat. Conserv. J. 2022, 77, 34–39. [Google Scholar]
- Gorman, A.M. Studies on the Biology of Plagiorchis elegans (Rudolphi, 1802), (Trematoda: Digenea) in Its Mammalian and Molluscan Hosts. Doctoral Thesis, University of Leeds, Leeds, UK, 1980. [Google Scholar]
- Ponomareva, N.M.; Popova, O.N.; Yurlova, N.I. Odonata (Insecta) larvae as the second intermediate hosts of the trematodes of genus Plagiorchis in the Basin of Chany Lake, Western Siberia. Contemp. Probl. Ecol. 2022, 15, 631–641. [Google Scholar] [CrossRef]
- Macy, R.W. The Life Cycle of Plagiorchis vespertilionis parorchis, n. ssp., (Trematoda: Plagiorchiidae), and observations on the effects of light on the emergence of the cercaria. J. Parasitol. 1960, 46, 337–345. [Google Scholar] [CrossRef]
- Chervy, L. Manual for the study of tapeworms (Cestoda) parasitic in ray-finned fish, amphibians and reptiles. Folia Parasitol. 2024, 71, 001. [Google Scholar] [CrossRef]






| Species | Snail Host | Locality | Life-Cycle Stage | Isolate | GenBank Accession Number | ||
|---|---|---|---|---|---|---|---|
| cox1 1 | cox1 2 | 28S | |||||
| Plagiorchis elegans | Lymnaea stagnalis | Milada | C | U144 | PP396753 | PP387839 | — |
| (Rudolphi, 1802) | Lymnaea stagnalis | Milada | C | U149 | PP396751 | PP387840 | — |
| Plagiorchis koreanus | Ampullaceana balthica | Medard | C | PK10 | PP396763 | — | — |
| Ogata, 1938 | Ampullaceana balthica | Medard | C | PK14 | PP396761 | — | — |
| Ampullaceana balthica | Medard | C | PK33 | PP396756 | PP387841 | — | |
| Ampullaceana balthica | Medard | C | KJ3 | PP396794 | PP387842 | — | |
| Ampullaceana balthica | Medard | C | KJ5 | PP396793 | PP387843 | — | |
| Ampullaceana balthica | Medard | C | KJ28 | PP396778 | — | — | |
| Ampullaceana balthica | Medard | C | KJ41 | PP396774 | PP387844 | — | |
| Ampullaceana balthica | Medard | C | KJ48 | PP396771 | PP387845 | — | |
| Ampullaceana balthica | Medard | C | KJ49 | PP396770 | PP387846 | — | |
| Ampullaceana lagotis | Milada | C | U142 | PP396754 | — | — | |
| Ampullaceana lagotis | Most | C | KJ14 | PP396787 | PP387847 | — | |
| Radix auricularia | Barbora | C | PK27 | PP396757 | PP387848 | — | |
| Radix auricularia | Barbora | C | E21 | PP396797 | PP387849 | — | |
| Radix auricularia | Písník Dubina | C | KJ23 | PP396780 | PP387850 | — | |
| Radix auricularia | Písník Dubina | C | KJ24 | PP396779 | PP387851 | — | |
| Plagiorchis maculosus | Ampullaceana balthica | Medard | C | PK18 | — | PP387852 | — |
| (Rudolphi, 1802) | Ampullaceana balthica | Medard | C | PK24 | — | PP387853 | — |
| Ampullaceana balthica | Medard | C | KJ26 | — | PP387854 | — | |
| Ampullaceana balthica | Medard | C | KJ4 | — | PP387855 | — | |
| Ampullaceana balthica | Medard | C | U147 | — | PP387856 | — | |
| Ampullaceana balthica | Medard | C | KJ55 | — | PP387857 | — | |
| Radix auricularia | Barbora | C | E20 | — | PP387858 | — | |
| Radix auricularia | Barbora | C | PK7 | PP396764 | PP387859 | — | |
| Radix auricularia | Barbora | C | PK15 | — | PP387860 | — | |
| Radix auricularia | Písník Dubina | S | KJ25 | — | PP387861 | — | |
| Radix auricularia | Spůle | C | KJ38 | PP396776 | PP387862 | — | |
| Radix auricularia | Spůle | C | KJ39 | — | PP387863 | — | |
| Radix auricularia | Spůle | C | KJ40 | PP396775 | PP387864 | — | |
| Plagiorchis muelleri | Ampullaceana balthica | Medard | C | PK2 | PP396765 | PP387865 | — |
| Tkach and Sharpilo, 1990 | Ampullaceana balthica | Medard | C | PK8 | — | PP387866 | — |
| Ampullaceana balthica | Medard | C | PK19 | — | PP387867 | — | |
| Ampullaceana balthica | Medard | C | PK47 | — | PP387868 | — | |
| Radix auricularia | Barbora | C | KJ51 | PP396768 | PP387869 | — | |
| Radix auricularia | Otakar | C | KJ22 | PP396781 | PP387870 | — | |
| Plagiorchis vespertilionis | Ampullaceana balthica | Medard | C | KJ2 | PP396795 | PP387871 | PP391294 |
| (Müller, 1780) | Ampullaceana balthica | Medard | C | KJ31 | PP396777 | PP387872 | — |
| Ampullaceana balthica | Medard | C | KJ53 | PP396766 | PP387873 | — | |
| Ampullaceana balthica | Medard | C | KJ9 | PP396790 | PP387874 | — | |
| Ampullaceana balthica | Medard | C | PK22 | PP396759 | PP387875 | — | |
| Ampullaceana lagotis | Most | C | KJ13 | PP396788 | PP387876 | PP391295 | |
| Ampullaceana lagotis | Most | C | KJ16 | PP396786 | PP387877 | — | |
| Ampullaceana lagotis | Most | C | KJ18 | PP396784 | PP387878 | — | |
| Ampullaceana lagotis | Most | C | KJ19 | PP396783 | PP387879 | — | |
| Ampullaceana lagotis | Most | C | KJ21 | PP396782 | PP387880 | — | |
| Ampullaceana lagotis | Most | C | KJ36 | - | PP387881 | — | |
| Ampullaceana lagotis | Most | C | KJ50 | PP396769 | PP387882 | — | |
| Ampullaceana lagotis | Most | S | KJ17 | PP396785 | PP387883 | — | |
| Radix auricularia | Barbora | C | PK26 | PP396758 | PP387884 | — | |
| Radix auricularia | Otakar | C | PK17 | PP396760 | PP387885 | — | |
| Plagiorchis sp. 3 sensu | Ampullaceana balthica | Medard | C | KJ1 | PP396796 | PP387886 | — |
| Soldánová et al. [17] | Ampullaceana balthica | Medard | C | KJ7 | PP396792 | PP387887 | — |
| Ampullaceana balthica | Medard | C | KJ8 | PP396791 | PP387888 | — | |
| Ampullaceana balthica | Medard | C | KJ10 | — | PP387889 | — | |
| Ampullaceana balthica | Medard | C | KJ33 | — | PP387890 | — | |
| Ampullaceana balthica | Medard | C | KJ43 | — | PP387891 | — | |
| Ampullaceana balthica | Medard | C | KJ45 | — | PP387892 | — | |
| Plagiorchis sp. 5 sensu | Ampullaceana balthica | Medard | C | KJ11 | PP396789 | PP387893 | |
| Soldánová et al. [17] | Ampullaceana balthica | Medard | C | KJ44 | PP396773 | PP387894 | — |
| Plagiorchis sp. 8 sensu | Ampullaceana balthica | Medard | C | U145 | PP396752 | PP387895 | — |
| Kudlai et al. [6] | Ampullaceana balthica | Medard | C | PK11 | PP396762 | PP387896 | — |
| Ampullaceana balthica | Medard | C | KJ46 | PP396772 | PP387897 | — | |
| Plagiorchis sp. 10 | Ampullaceana balthica | Medard | C | U165 | PP396750 | — | — |
| Ampullaceana balthica | Skulska Wieś | C | KJ52 | PP396767 | PP387898 | PP391292 | |
| Ampullaceana lagotis | Most | C | U140 | PP396755 | PP387899 | PP391293 | |
| Plagiorchis sp. 11 | Ampullaceana lagotis | Most | C | PK13 | PP407075 | PP387900 | PP391291 |
| Species | Plagiorchis vespertilionis 1 | Plagiorchis sp. 10 2 | Plagiorchis sp. 11 3 | |||||
|---|---|---|---|---|---|---|---|---|
| N snails | 9 | 14 | 2 | 2 | 3 | 1 | 1 | 1 |
| N cercariae | 11 | 113 | 29 | 5 | 49 | 19 | 1 | 5 |
| Fixation method | Live | Ethanol | Formalin | Live | Ethanol | Formalin | Live | Ethanol |
| ToL | 332–425 (379) | 233–446 (360) | 263–387 (328) | 353–513 (417) | 244–356 (317) | 225–316 (266) | 440 | 348–400 (368) |
| BL | 190–320 (254) | 153–290 (211) | 171–217 (190) | 238–372 (298) | 145–227 (191) | 124–165 (147) | 302 | 221–247 (231) |
| BW | 124–202 (159) | 70–145 (115) | 103–135 (119) | 132–156 (148) | 71–102 (88) | 77–101 (92) | 158 | 120–137 (129) |
| TL | 99–156 (125) | 96–200 (150) | 77–195 (138) | 87–141 (119) | 99–153 (125) | 73–169 (119) | 138 | 121–154 (137) |
| TW | 25–57 (38) | 19–38 (29) | 22–48 (32) | 39–53 (45) | 20–30 (25) | 18–29 (23) | 44 | 36–40 (39) |
| OSL | 48–80 (67) | 39–64 (50) | 39–57 (48) | 59–69 (63) | 35–52 (42) | 36–42 (38) | 62 | 44–58 (50) |
| OSW | 50–73 (64) | 41–67 (54) | 48–63 (57) | 57–69 (65) | 36–55 (48) | 39–45 (42) | 67 | 50–59 (56) |
| VSL | 35–57 (46) | 28–49 (37) | 32–43 (38) | 43–50 (46) | 27–45 (32) | 24–33 (30) | 39 | 30–41 (35) |
| VSW | 37–57 (47) | 34–58 (44) | 40–52 (45) | 45–58 (51) | 32–46 (38) | 31–37 (35) | 38 | 34–47 (39) |
| OSW/VSW | 1.3–1.4 (1.4) | 0.9–1.7 (1.2) | 1.1–1.6 (1.3) | 1.0–1.5 (1.3) | 1.0–1.6 (1.3) | 1.1–1.3 (1.2) | 1.7 | 1.2–1.7 (1.4) |
| TL/BL (%) | 33.0–74.9 (51.0) | 47.8–95.7 (71.0) | 41.4–110.9 (73.5) | 30.3–48.7 (40.5) | 52.8–92.1 (66.1) | 44.2–134.7 (82.5) | 45.7 | 53.3–69.7 (59.5) |
| SL | 30.0–31.8 (31.0) | 27–33 (30) | – | 28.8–30.3 (29.5) | 26–29 (27) | – | 31.1 | 29–32 (31) |
| SWantt | 7.1–9.0 (7.8) | – | – | 6.7–6.8 (6.7) | – | – | 8.9 | – |
| SWabt | 4.2–5.3 (4.8) | – | – | 4.3–4.5 (4.4) | – | – | 5.3 | – |
| SWbt | 4.2–5.7 (5.0) | – | – | 5.1–5.2 (5.1) | – | – | 5.3 | – |
| SWantt/SL (%) | 22.8–28.7 (25.2) | – | – | 22.1–23.6 (22.8) | – | – | 28.6 | – |
| Region | ||||
|---|---|---|---|---|
| Plagiorchis Species | Central Europe (Present Study) | Central Europe 1 | Sub-Arctic Europe 2,3 | Western Europe 3,4,5 |
| Plagiorchis elegans | ||||
| Plagiorchis koreanus | ||||
| Plagiorchis maculosus | ||||
| Plagiorchis muelleri | ||||
| Plagiorchis neomidis | ||||
| Plagiorchis vespertilionis * | ||||
| Plagiorchis sp. 1 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 2 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 3 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 4 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 5 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 6 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 7 sensu Soldánová et al. [17] | ||||
| Plagiorchis sp. 8 sensu Kudlai et al. [6] | ||||
| Plagiorchis sp. 9 sensu Kudlai et al. [6] | ||||
| Plagiorchis sp. CR sensu Zikmundová et al. [40] | ||||
| Plagiorchis sp. sensu Duan et al. [18] | ||||
| Plagiorchis sp. sensu Huguenin et al. [51] | ||||
| Plagiorchis sp. 10 * | ||||
| Plagiorchis sp. 11 * | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundid, P.; Pantoja, C.; Janovcová, K.; Soldánová, M. Molecular Diversity of the Genus Plagiorchis Lühe, 1899 in Snail Hosts of Central Europe with Evidence of New Lineages. Diversity 2024, 16, 158. https://doi.org/10.3390/d16030158
Kundid P, Pantoja C, Janovcová K, Soldánová M. Molecular Diversity of the Genus Plagiorchis Lühe, 1899 in Snail Hosts of Central Europe with Evidence of New Lineages. Diversity. 2024; 16(3):158. https://doi.org/10.3390/d16030158
Chicago/Turabian StyleKundid, Petra, Camila Pantoja, Kristýna Janovcová, and Miroslava Soldánová. 2024. "Molecular Diversity of the Genus Plagiorchis Lühe, 1899 in Snail Hosts of Central Europe with Evidence of New Lineages" Diversity 16, no. 3: 158. https://doi.org/10.3390/d16030158
APA StyleKundid, P., Pantoja, C., Janovcová, K., & Soldánová, M. (2024). Molecular Diversity of the Genus Plagiorchis Lühe, 1899 in Snail Hosts of Central Europe with Evidence of New Lineages. Diversity, 16(3), 158. https://doi.org/10.3390/d16030158