
Effects of Invasive Smooth Cordgrass Degradation on Avian Species Diversity in the Dafeng Milu National Nature Reserve, a Ramsar Wetland on the Eastern Coast of China


Abstract
:1. Introduction
2. Materials and Methods
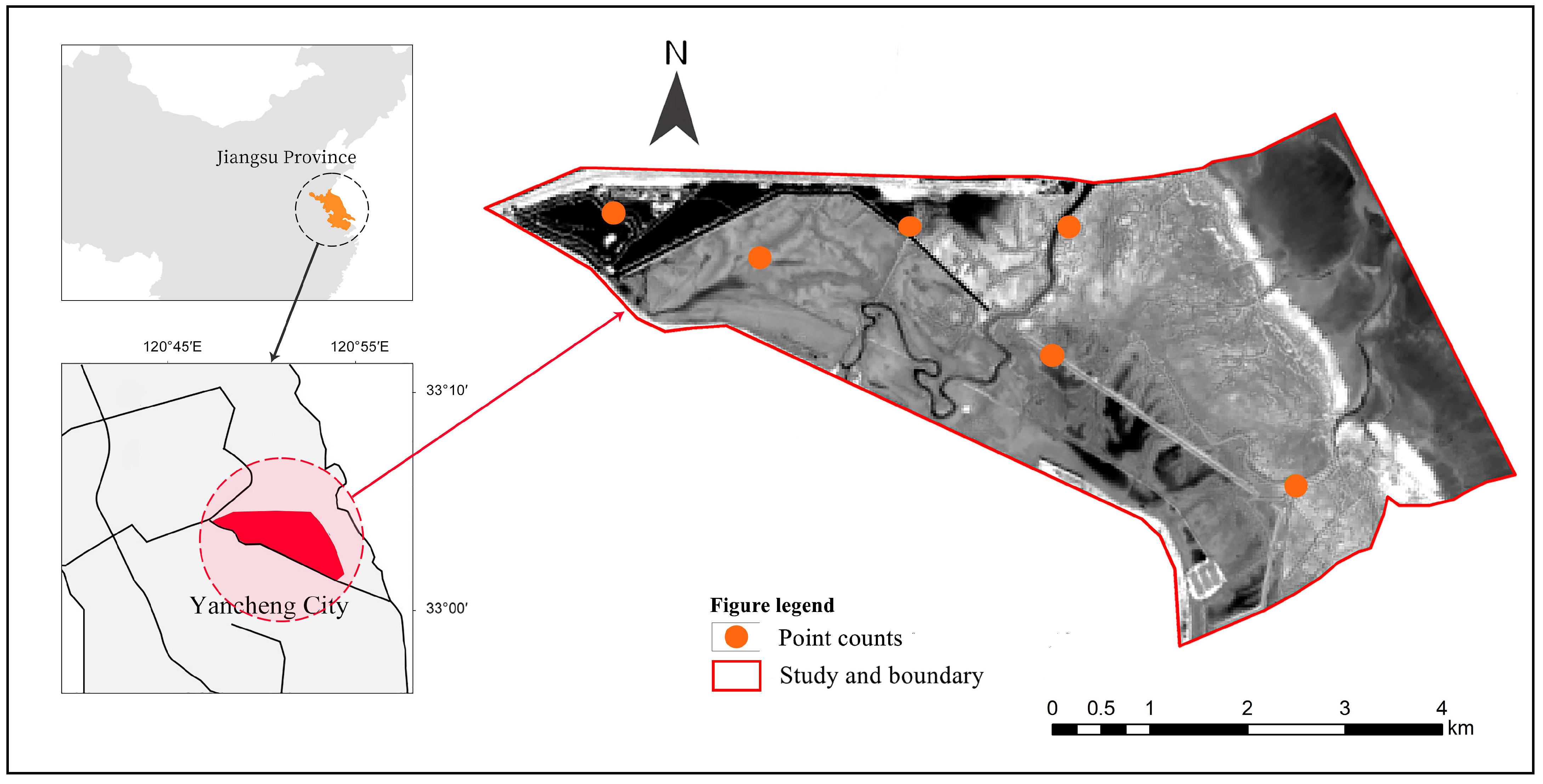
2.1. Study Area
2.2. Field Surveys
2.3. Data Analyses
3. Results
3.1. Changes in Area Covered by Spartina from 2017 to 2021
3.2. Bird Species
3.3. Bird Abundance and Density
3.4. Spatial Variations of Bird Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. The Average Number of Birds during 2017 to 2021 in DMNNR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Scientific Name | IUCN | 2017 (n = 12) | 2018 (n = 12) | 2019 (n = 12) | 2020 (n = 12) | 2021 (n = 12) |
|---|---|---|---|---|---|---|---|
| Little Grebe | Tachybaptus ruficollis | 14.4 | 13.6 | 17.0 | 4.8 | 5.9 | |
| Great Crested Grebe | Podiceps cristatus | 0.3 | 0.2 | ||||
| Great Cormorant | Phalacrocorax carbo | 6.9 | 30.9 | 107.3 | 228.8 | 223.5 | |
| Grey Heron | Ardea cinerea | 34.0 | 46.9 | 69.9 | 112.8 | 103.6 | |
| Great Egret | Egretta alba | 15.2 | 23.9 | 40.9 | 57.5 | 36.5 | |
| Intermediate Egret | Egretta intermedia | 12.2 | 16.5 | 23.7 | 16.9 | 19.8 | |
| Little Egret | Egretta garzetta | 23.2 | 37.2 | 66.8 | 55.3 | 37.9 | |
| Cattle Egret | Bubulcus ibis | 26.5 | 29.4 | 31.3 | 19.8 | 4.4 | |
| Chinese Pond Heron | Ardeola bacchus | 2.0 | 5.0 | 5.1 | 0.8 | ||
| Striated Heron | Butorides striata | 1.8 | 0.8 | 0.3 | |||
| Black-crowned Night Heron | Nycticorax nycticorax | 5.3 | 6.3 | 15.5 | 2.6 | 3.2 | |
| Eurasian Bittern | Botaurus stellaris | 0.8 | 0.3 | 0.3 | |||
| Oriental Stork | Ciconia boyciana | EN | 3.2 | 13.8 | 15.8 | 2.4 | 1.5 |
| Eurasian Spoonbill | Platalea leucorodia | 17.1 | 10.2 | 28.0 | 24.5 | 28.2 | |
| Black-faced Spoonbill | Platalea minor | EN | 2.6 | 1.2 | 1.6 | 3.3 | 2.9 |
| Bean Goose | Anser fabalis | 12.7 | 11.9 | 45.5 | 5.4 | 37.6 | |
| Bean Goose | Anser albifrons | 0.9 | 3.8 | 0.7 | 1.8 | ||
| Lesser White-fronted Goose | Anser erythropus | VU | 0.3 | 0.3 | 0.8 | ||
| Common Shelduck | Tadorna tadorna | 9.2 | |||||
| Gadwall | Anas strepera | 9.2 | 20.1 | 24.5 | 5.8 | 1.6 | |
| Falcated Duck | Anas falcata | NT | 3.7 | 4.7 | 0.8 | ||
| Eurasian Wigeon | Anas penelope | 5.3 | 2.5 | 1.3 | 9.9 | 11.7 | |
| Mallard | Anas platyrhynchos | 25.7 | 2.8 | 45.3 | 28.6 | 6.8 | |
| Eastern Spot-billed Duck | Anas poecilorhyncha | 95.1 | 92.8 | 186.7 | 120.6 | 44.0 | |
| Northern Shoveler | Anas clypeata | 1.8 | 2.4 | 25.8 | 0.5 | 16.8 | |
| Northern Pintail | Anas acuta | 2.6 | 0.9 | 2.3 | 4.5 | ||
| Garganey | Anas querquedula | 1.7 | 0.6 | 3.9 | 1.9 | ||
| Green-winged Teal | Anas crecca | 44.7 | 38.8 | 54.2 | 47.0 | 23.3 | |
| Common Pochard | Aythya ferina | VU | 35.9 | 14.7 | 15.5 | 6.3 | 1.8 |
| Ferruginous Duck | Aythya nyroca | NT | 11.9 | 1.2 | |||
| Tufted Duck | Aythya fuligula | 13.6 | 9.5 | 4.2 | 2.5 | ||
| Greater Scaup | Aythya marila | 2.3 | 1.2 | 0.4 | 2.9 | ||
| Smew | Mergellus albellus | 1.0 | 1.5 | ||||
| Common Pheasant | Phasianus colchicus | 5.5 | 1.7 | 0.8 | 0.3 | ||
| Common Crane | Grus grus | 0.3 | |||||
| Red-crowned Crane | Grus japonensis | EN | 0.9 | 0.3 | 0.3 | 0.3 | |
| Common Moorhen | Gallinula chloropus | 7.3 | 3.8 | 1.5 | 1.9 | 0.4 | |
| Common Coot | Fulica atra | 17.7 | 17.8 | 24.8 | 20.6 | 10.7 | |
| Black-winged Stilt | Himantopus himantopus | 3.3 | 3.0 | 5.5 | 15.5 | 3.7 | |
| Pied Avocet | Himantopus himantopus | 18.0 | 70.4 | 93.2 | 228.3 | 77.5 | |
| Oriental Pratincole | Glareolidae | 6.7 | 8.7 | 3.5 | 1.6 | 0.8 | |
| Northern Lapwing | Vanellus vanellus | NT | 6.2 | 7.3 | 34.2 | 10.9 | 4.8 |
| Grey-headed Lapwing | Vanellus cinereus | 16.9 | 11.2 | 12.7 | 24.5 | 22.7 | |
| Pacific Golden Plover | Pluvialis fulva | 0.3 | 1.0 | 0.5 | 3.2 | ||
| Grey Plover | Pluvialis squatarola | 0.7 | 1.9 | 0.3 | 3.0 | 4.4 | |
| Little Ringed Plover | Charadrius dubius | 3.9 | 3.0 | 5.3 | 3.6 | 3.3 | |
| Kentish Plover | Charadrius dubius | 6.5 | 9.0 | 18.9 | 27.0 | 14.3 | |
| Lesser Sand Plover | Charadrius mongolus | 0.1 | 2.3 | 0.9 | 0.8 | 7.8 | |
| Greater Sand Plover | Charadrius leschenaultii | 0.9 | 2.3 | 11.3 | 1.8 | 7.9 | |
| Common Snipe | Gallinago gallinago | 2.8 | 2.3 | 0.2 | 1.0 | 0.2 | |
| Long-billed Dowitcher | Limnodromus scolopaceus | 0.1 | 0.1 | ||||
| Black-tailed Godwit | Limosa limosa | 2.2 | 61.7 | 30.4 | 16.3 | 12.2 | |
| Bar-tailed Godwit | Limosa lapponica | 4.2 | 5.3 | 7.8 | 2.6 | 6.5 | |
| Eurasian Curlew | Numenius arquata | NT | 9.8 | 2.1 | 3.4 | 27.0 | 13.3 |
| Eastern Curlew | Numenius madagascariensis | EN | 1.8 | 0.8 | 0.9 | 2.0 | 1.1 |
| Spotted Redshank | Tringa erythropus | 6.1 | 6.1 | 10.8 | 3.5 | 4.4 | |
| Common Redshank | Tringa totanus | 10.0 | 12.2 | 9.4 | 5.9 | 11.4 | |
| Marsh Sandpiper | Tringa stagnatilis | 2.2 | 1.3 | 1.3 | 16.1 | 2.5 | |
| Common Greenshank | Tringa nebularia | 5.3 | 4.5 | 5.7 | 19.5 | 5.7 | |
| Green Sandpiper | Tringa ochropus | 0.1 | 0.1 | 0.8 | 0.8 | 0.5 | |
| Wood Sandpiper | Tringa glareola | 0.1 | 0.1 | 0.1 | 0.1 | 0.3 | |
| Common Sandpiper | Actitis hypoleucos | 1.4 | 0.4 | 0.3 | 3.3 | 0.7 | |
| Grey-tailed Tattler | Heteroscelus brevipes | 0.5 | 0.8 | 1.0 | |||
| Sanderling | Calidris alba | 33.8 | 12.5 | ||||
| Red-necked Stint | Calidris ruficollis | 5.0 | 165.0 | 38.3 | |||
| Sharp-tailed Sandpiper | Calidris acuminata | 15.3 | 10.7 | 19.8 | 85.0 | 88.3 | |
| Dunlin | Calidris alpina | 8.7 | 19.4 | 45.0 | 67.8 | 152.5 | |
| Curlew Sandpiper | Calidris ferruginea | 0.8 | 0.2 | ||||
| Broad-billed Sandpiper | Limicola falcinellus | 0.5 | 0.3 | ||||
| Eurasian Oystercatcher | Haematopus ostralegus | 0.7 | 0.3 | ||||
| Black-tailed Gull | Larus crassirostris | 0.4 | 0.4 | 0.3 | 0.4 | 0.9 | |
| Caspian Gull | Larus cachinnans | 0.3 | 1.7 | 1.7 | 2.3 | 6.6 | |
| Slaty-backed Gull | Larus schistisagus | 0.1 | 0.6 | 14.0 | 1.4 | 0.5 | |
| Black-headed Gull | Chroicocephalus ridibundus | 18.0 | 18.4 | 10.8 | 67.9 | 21.6 | |
| Saunders’s Gull | Saundersilarus saundersi | VU | 5.0 | 8.6 | 25.9 | 4.4 | 29.7 |
| Gull-billed Tern | Gelochelidon nilotica | 1.7 | 12.5 | 1.0 | |||
| Common Tern | Sterna hirundo | 16.8 | 21.8 | 11.9 | 5.5 | 7.8 | |
| Little Tern | Sternula albifrons | 16.3 | 12.3 | 30.8 | 19.8 | 8.9 | |
| White-winged Tern | Chlidonias leucoptera | 12.8 | 1.7 | ||||
| Common Cuckoo | Cuculus canorus | 1.8 | 1.5 | 1.3 | 1.4 | 0.4 | |
| Lesser Coucal | Centropus toulou | 1.3 | 1.2 | 0.5 | 0.1 | ||
| Common Kingfisher | Alcedo atthis | 2.0 | 1.4 | 0.3 | 0.3 | 0.2 | |
| Pied Kingfisher | Ceryle rudis | 1.1 | 1.8 | 1.0 | 0.5 | 0.2 | |
| Common Hoopoe | Upupa epops | 2.2 | 1.8 | 1.4 | 0.5 | 0.4 | |
| Oriental Skylark | Alauda gulgula | 2.2 | 1.8 | 2.7 | 0.3 | 1.0 | |
| Barn Swallow | Hirundo rustica | 15.4 | 14.8 | 11.0 | 11.1 | 17.3 | |
| Red-rumped Swallow | Cecropis daurica | 7.1 | 7.8 | 9.5 | 9.8 | 10.4 | |
| White Wagtail | Motacilla alba | 2.1 | 0.8 | 1.0 | 2.0 | 0.5 | |
| Gray Wagtail | Motacilla cinerea | 3.5 | 1.3 | 0.3 | 0.5 | 0.2 | |
| Olive-backed Pipit | Anthus hodgsoni | 1.1 | 0.5 | ||||
| Light-vented Bulbul | Pycnonotus sinensis | 6.3 | 5.5 | 6.7 | 4.1 | 3.5 | |
| Tiger Shrike | Lanius tigrinus | 0.5 | 0.1 | ||||
| Long-tailed Shrike | Lanius schach | 4.1 | 4.2 | 2.6 | 1.5 | 1.2 | |
| Brown Shrike | Lanius cristatus | 2.0 | 1.8 | 2.9 | 0.6 | 0.3 | |
| Black Drongo | Dicrurus macrocercus | 9.1 | 9.5 | 11.1 | 5.5 | 3.6 | |
| White-cheeked Starling | Sturnus cineraceus | 9.6 | 12.5 | 11.3 | 8.8 | 1.9 | |
| Silky Starling | Spodiopsar sericeus | 11.4 | 5.8 | 10.1 | 6.8 | 0.3 | |
| Common Magpie | Pica Pica | 8.6 | 9.8 | 10.3 | 3.4 | 1.7 | |
| Orange-flanked Bluetail | Tarsiger cyanurus | 0.8 | 0.3 | 0.3 | |||
| Daurian Redstart | Phoenicurus auroreus | 3.5 | 1.7 | 1.7 | |||
| Siberian Thrush | Geokichla sibirica | 0.3 | |||||
| Dusky Thrush | Turdus eunomus | 0.9 | 0.7 | 3.0 | |||
| Grey-streaked Flycatcher | Muscicapa griseisticta | 1.3 | 0.8 | 1.3 | |||
| Mugimaki Flycatcher | Ficedula mugimaki | 0.5 | 0.3 | ||||
| Vinous-throated Parrotbill | Paradoxornis webbianus | NT | 43.8 | 52.8 | 40.3 | 10.5 | 3.0 |
| Reed Parrotbill | Paradoxornis heudei | 12.8 | 13.5 | 17.0 | 2.0 | ||
| Zitting Cisticola | Cisticola juncidis | 17.6 | 10.8 | 7.1 | 2.8 | 0.2 | |
| Plain Prinia | Prinia inornata | 14.2 | 11.6 | 9.5 | 3.3 | 0.5 | |
| Oriental Reed Warbler | Acrocephalus orientalis | 2.8 | 4.2 | 4.3 | 1.3 | ||
| Black-browed Reed Warbler | Acrocephalus bistrigiceps | 1.5 | 2.6 | ||||
| Yellow-browed Warbler | Phylloscopus inornatus | 0.3 | 0.2 | 0.7 | |||
| Marsh Grassbird | Locustella pryeri | 14.0 | 5.1 | 1.9 | |||
| Chinese Penduline Tit | Remiz consobrinus | 4.8 | 5.2 | 2.3 | 1.6 | ||
| Cinereous Tit | Parus major | 3.5 | 3.3 | 3.1 | 1.3 | 0.2 | |
| House Sparrow | Passer montanus | 38.8 | 27.8 | 11.5 | 7.1 | 2.9 | |
| Brambling | Fringilla montifringilla | 3.8 | 3.2 | ||||
| Meadow Bunting | Emberiza ioides | 4.6 | 8.7 | 6.7 | 1.3 | ||
| Chestnut-eared Bunting | Emberiza fucata | 3.2 | 1.8 | 0.9 | 0.1 | ||
| Little Bunting | Emberiza pusilla | 7.0 | 8.8 | 8.2 | 2.3 | ||
| Rustic Bunting | Emberiza rustica | 8.2 | 12.6 | 10.8 | 1.3 | 0.8 | |
| Chestnut Bunting | Emberiza rutila | 0.5 | 0.3 | 0.3 | |||
| Black-faced Bunting | Emberiza spodocephala | 19.4 | 29.2 | 15.8 | 4.8 | 0.3 | |
| Pallas’s Bunting | Emberiza pallasi | 17.5 | 10.3 | 6.8 | 1.5 | 0.9 | |
| Reed Bunting | Emberiza schoeniclus | 4.7 | 0.8 | 0.3 |
Appendix B. The Average Bird Number ≥ 100 Individuals during 2017 to 2021 in Different Groups
| Group | Common Name | Abbreviation | River | Open Water | Mudflat | Common Reed | Spartina | Sum |
|---|---|---|---|---|---|---|---|---|
| Swan, goose, and duck | Bean Goose | BG | 17 | 55 | 175 | 247 | ||
| Gadwall | G | 42 | 90 | 10 | 142 | |||
| Mallard | M | 105 | 120 | 7 | 2 | 5 | 239 | |
| Eastern Spot-billed Duck | ESD | 300 | 380 | 200 | 100 | 17 | 997 | |
| Northern Shoveler | NS | 20 | 80 | 13 | 113 | |||
| Green-winged Teal | G-wT | 200 | 150 | 50 | 21 | 40 | 461 | |
| Common Pochard | CP | 73 | 100 | 173 | ||||
| Shorebird | Pied Avocet | PA | 266 | 439 | 252 | 30 | 987 | |
| Northern Lapwing | NL | 23 | 7 | 101 | 12 | 143 | ||
| Grey-headed Lapwing | G-hL | 44 | 12 | 129 | 6 | 191 | ||
| Black-tailed Godwit | B-tG | 70 | 204 | 5 | 2 | 281 | ||
| Common Redshank | CR | 9 | 103 | 1 | 113 | |||
| Sharp-tailed Sandpiper | S-tS | 27 | 120 | 105 | 30 | 180 | 458 | |
| Dunlin | D | 183 | 200 | 175 | 50 | 550 | ||
| Black-headed Gull | B-hG | 90 | 100 | 74 | 10 | 274 | ||
| Saunders’s Gull | SG | 30 | 73 | 60 | 10 | 173 | ||
| Common Tern | CT | 40 | 90 | 19 | 149 | |||
| Little Tern | LT | 66 | 110 | 20 | 196 | |||
| Songbird | Barn Swallow | BS | 45 | 50 | 18 | 25 | 20 | 158 |
| Vinous-throated Parrotbill | V-tP | 13 | 200 | 100 | 313 | |||
| Reed Parrotbill | RP | 105 | 2 | 107 | ||||
| House Sparrow | HS | 28 | 26 | 30 | 40 | 22 | 146 | |
| Black-faced Bunting | B-fB | 7 | 33 | 90 | 12 | 142 | ||
| Others | Little Grebe | LG | 34 | 82 | 116 | |||
| Great Cormorant | GC | 174 | 556 | 401 | 1131 | |||
| Grey Heron | GH | 268 | 100 | 300 | 42 | 27 | 737 | |
| Great Egret | GE | 113 | 155 | 48 | 52 | 4 | 372 | |
| Little Egret | LE | 77 | 163 | 59 | 37 | 22 | 484 | |
| Cattle Egret | CE | 40 | 33 | 101 | 28 | 5 | 207 | |
| Eurasian Spoonbill | ES | 88 | 105 | 12 | 34 | 219 | ||
| Common Coot | CC | 79 | 120 | 4 | 203 |
References
- Davidsdottir, B.; Gunnarsson, T.G.; Halldorsson, G.; Sigurdsson, B.D. Avian abundance and communities in areas revegetated with exotic versus native plant species. Icel. Agric. Sci. 2016, 29, 21–37. [Google Scholar] [CrossRef]
- Jackson, M.V.; Fuller, R.A.; Gan, X.; Li, J.; Mao, D.; Melville, D.S.; Murray, N.J.; Wang, Z.; Choi, C.-Y. Dual threat of tidal flat loss and invasive Spartina alterniflora endanger important shorebird habitat in coastal mainland China. J. Environ. Manag. 2021, 278, 111549. [Google Scholar] [CrossRef]
- Gan, X.; Choi, C.; Wang, Y.; Ma, Z.; Chen, J.; Li, B. Alteration of Habitat Structure and Food Resources by Invasive Smooth Cordgrass Affects Habitat Use by Wintering Saltmarsh Birds at Chongming Dongtan, East China. Auk 2010, 127, 317–327. [Google Scholar] [CrossRef]
- Gan, X.; Cai, Y.; Choi, C.; Ma, Z.; Chen, J.; Li, B. Potential impacts of invasive Spartina alterniflora on spring bird communities at Chongming Dongtan, a Chinese wetland of international importance. Estuar. Coast. Shelf Sci. 2009, 83, 211–218. [Google Scholar] [CrossRef]
- Ma, Z.; Melville, D.S.; Liu, J.; Chen, Y.; Yang, H.; Ren, W.; Zhang, Z.; Piersma, T.; Li, B. Rethinking China’s new great wall. Sci. Am. Assoc. Adv. Sci. 2014, 346, 912–914. [Google Scholar] [CrossRef]
- Chen, P.; Chen, Y.; Chen, H.; Chen, T.; Liu, B.; Zhang, M.; Wang, S.; Lu, C. Vinous-throated parrotbills breed in invasive smooth cordgrass habitat: Can native birds avoid the potential ecological trap? Avian Res. 2023, 14, 100119. [Google Scholar] [CrossRef]
- Tang, L.; Gao, Y.; Wang, C.-H.; Li, B.; Chen, J.-K.; Zhao, B. Habitat heterogeneity influences restoration efficacy: Implications of a habitat-specific management regime for an invaded marsh. Estuar. Coast. Shelf Sci. 2013, 125, 20–26. [Google Scholar] [CrossRef]
- Shimeta, J.; Saint, L.; Verspaandonk, E.R.; Nugegoda, D.; Howe, S. Long-term ecological consequences of herbicide treatment to control the invasive grass, Spartina anglica, in an Australian saltmarsh. Estuar. Coast. Shelf Sci. 2016, 176, 58–66. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, J.; Yang, X.; Zhang, M.; Tang, C.; Wang, T. Impact of coastal wetland restoration strategies in the chongming dongtan wetlands, China: Waterbird community composition as an indicator. Acta Zool. Hung. 2014, 60, 185–198. [Google Scholar]
- Lampert, A.; Hastings, A.; Grosholz, E.D.; Jardine, S.L.; Sanchirico, J.N. Optimal approaches for balancing invasive species eradication and endangered species management. Science 2014, 344, 1028–1031. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Y.; Wang, G.; Xu, P.; Lu, C. Differences in the crab community structure between pristine and degraded Suaeda marshes after Spartina invasion. Reg. Stud. Mar. Sci. 2020, 34, 101001. [Google Scholar] [CrossRef]
- Duan, H.; Xia, S.; Jackson, M.V.; Zhao, N.; Liu, Y.; Teng, J.; Meng, Z.; Yu, X.; Shi, J. Identifying new sites of significance to waterbirds conservation and their habitat modification in the Yellow and Bohai Seas in China. Glob. Ecol. Conserv. 2020, 22, e01031. [Google Scholar] [CrossRef]
- Chen, P.; Chen, T.; Liu, B.; Zhang, M.; Lu, C.; Chen, Y. Snakes are the principal nest predators of the threatened reed parrotbill in a coastal wetland of eastern China. Glob. Ecol. Conserv. 2020, 23, e01055. [Google Scholar] [CrossRef]
- Yan, D.; Li, J.; Yao, X.; Luan, Z. Quantifying the Long-Term Expansion and Dieback ofSpartina Alterniflora Using Google Earth Engine and Object-Based Hierarchical Random Forest Classification. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 9781–9793. [Google Scholar] [CrossRef]
- Mao, D.; Liu, M.; Wang, Z.; Li, L.; Man, W.; Jia, M.; Zhang, Y. Rapid Invasion of Spartina Alterniflora in the Coastal Zone of Mainland China: Spatiotemporal Patterns and Human Prevention. Sensors 2019, 19, 2308. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, S.; Jia, Y.; Zeng, Q.; Feng, D.; Lei, G. Spatial and temporal variations in waterbird communities and its implications for ecosystem management in a large temperate arid wetland of northwest P.R. China. Avian Biol. Res. 2017, 2, 119–128. [Google Scholar] [CrossRef]
- Xu, H.F.; Zhao, Y.L. Comprehensive Surveys in Chongming Dongtan Nature Reserve for Migratory Birds, Shanghai; Chinese Forestry Publishing House: Beijing, China, 2005; p. 250. [Google Scholar]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1994, 345, 101–118. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of SPECIES Richness and Shared Species from Samples, Version 8.0; University of Connecticut: Storrs, CT, USA, 2006.
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 663. [Google Scholar]
- Zhang, Y.; Jia, Y.; Jiao, S.; Qing, Z.; Feng, G.; Guo, Y.; Lei, G. Wuliangsuhai Wetlands: A Critical Habitat for Migratory Water Birds. J. Resour. Ecol. 2012, 3, 316–323. [Google Scholar]
- Kang, W.; Zhang, Y.; Bai, J.; Zhu, A.; Chen, R.; Xue, D.; Zhong, Z.; Cheng, Z. Reversing extinction in China’s Père David’s deer. Science 2021, 371, 685. [Google Scholar]
- Sun, J.; Wang, X.; Chen, A.; Ma, Y.; Cui, M.; Piao, S. NDVI indicated characteristics of vegetation cover change in China’s metropolises over the last three decades. Environ. Monit. Assess. 2011, 179, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Chen, S.S.; Yan, Y.; Yu, C. Effects of Urbanization on Vegetation Degradation in the Yangtze River Delta of China: Assessment Based on SPOT-VGT NDVI. Am. Soc. Civ. Eng. 2014, 11, 0733–9488. [Google Scholar] [CrossRef]
- Li, S.; Ge, Z.-M.; Xie, L.-N.; Chen, W.; Yuan, L.; Wang, D.-Q.; Li, X.-Z.; Zhang, L.-Q. Ecophysiological response of native and exotic salt marsh vegetation to waterlogging and salinity: Implications for the effects of sea-level rise. Sci. Rep. 2018, 8, 2441. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-A.; Tang, C.-D.; Niu, J.-Y.; Wang, T.-H.; Xie, Y.-H.; Guo, H. Migratory Waterbirds Response to Coastal Habitat Changes: Conservation Implications from Long-term Detection in the Chongming Dongtan Wetlands, China. In Estuaries and Coasts, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 39, pp. 273–286. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Chen, P.; Liu, B.; Wu, D.; Lu, C. Effects of Invasive Smooth Cordgrass Degradation on Avian Species Diversity in the Dafeng Milu National Nature Reserve, a Ramsar Wetland on the Eastern Coast of China. Diversity 2024, 16, 176. https://doi.org/10.3390/d16030176
Chen T, Chen P, Liu B, Wu D, Lu C. Effects of Invasive Smooth Cordgrass Degradation on Avian Species Diversity in the Dafeng Milu National Nature Reserve, a Ramsar Wetland on the Eastern Coast of China. Diversity. 2024; 16(3):176. https://doi.org/10.3390/d16030176
Chicago/Turabian StyleChen, Taiyu, Pan Chen, Bing Liu, Dawei Wu, and Changhu Lu. 2024. "Effects of Invasive Smooth Cordgrass Degradation on Avian Species Diversity in the Dafeng Milu National Nature Reserve, a Ramsar Wetland on the Eastern Coast of China" Diversity 16, no. 3: 176. https://doi.org/10.3390/d16030176
APA StyleChen, T., Chen, P., Liu, B., Wu, D., & Lu, C. (2024). Effects of Invasive Smooth Cordgrass Degradation on Avian Species Diversity in the Dafeng Milu National Nature Reserve, a Ramsar Wetland on the Eastern Coast of China. Diversity, 16(3), 176. https://doi.org/10.3390/d16030176