
Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Collection and Bacterial Strains Isolation
2.2. Screening for Bacterial Isolates for AHLs Production
2.3. Identification and Phylogenetic Analysis of AHL-Producing Bacteria
2.4. Extraction of AHLs
2.5. Identification of AHLs by TLC and HPLC/MS
2.6. AHLs Quantification by β-Galactosidase Assay
2.7. Biofilm Formation Assay
2.8. Statistical Analysis
3. Result
3.1. Isolation and Identification of AHL-Producing Bacteria
3.2. Identification of AHL-Producing Bacteria
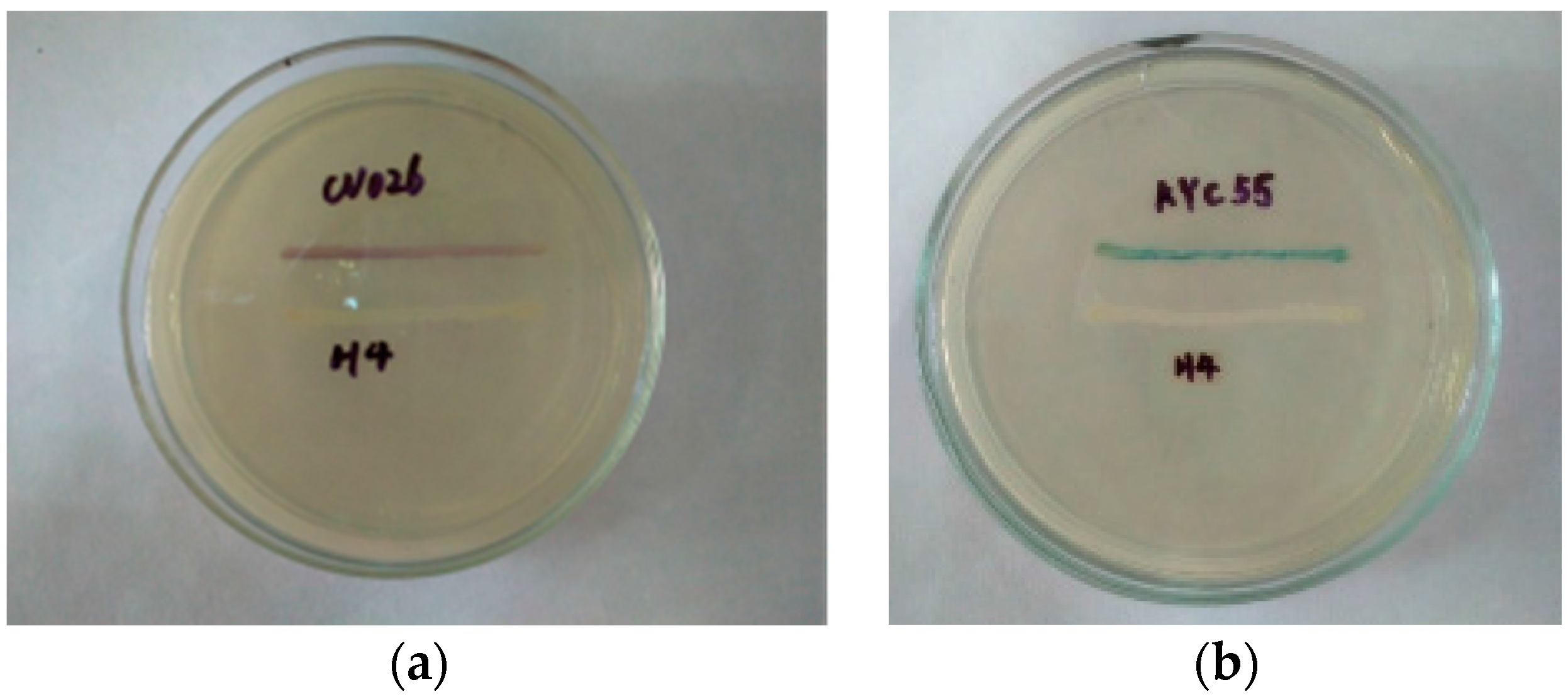
3.3. AHL Profile Analysis by TLC and HPLC-MS/MS
3.4. AHLs Production by H. alvei H4 in Growth Phase
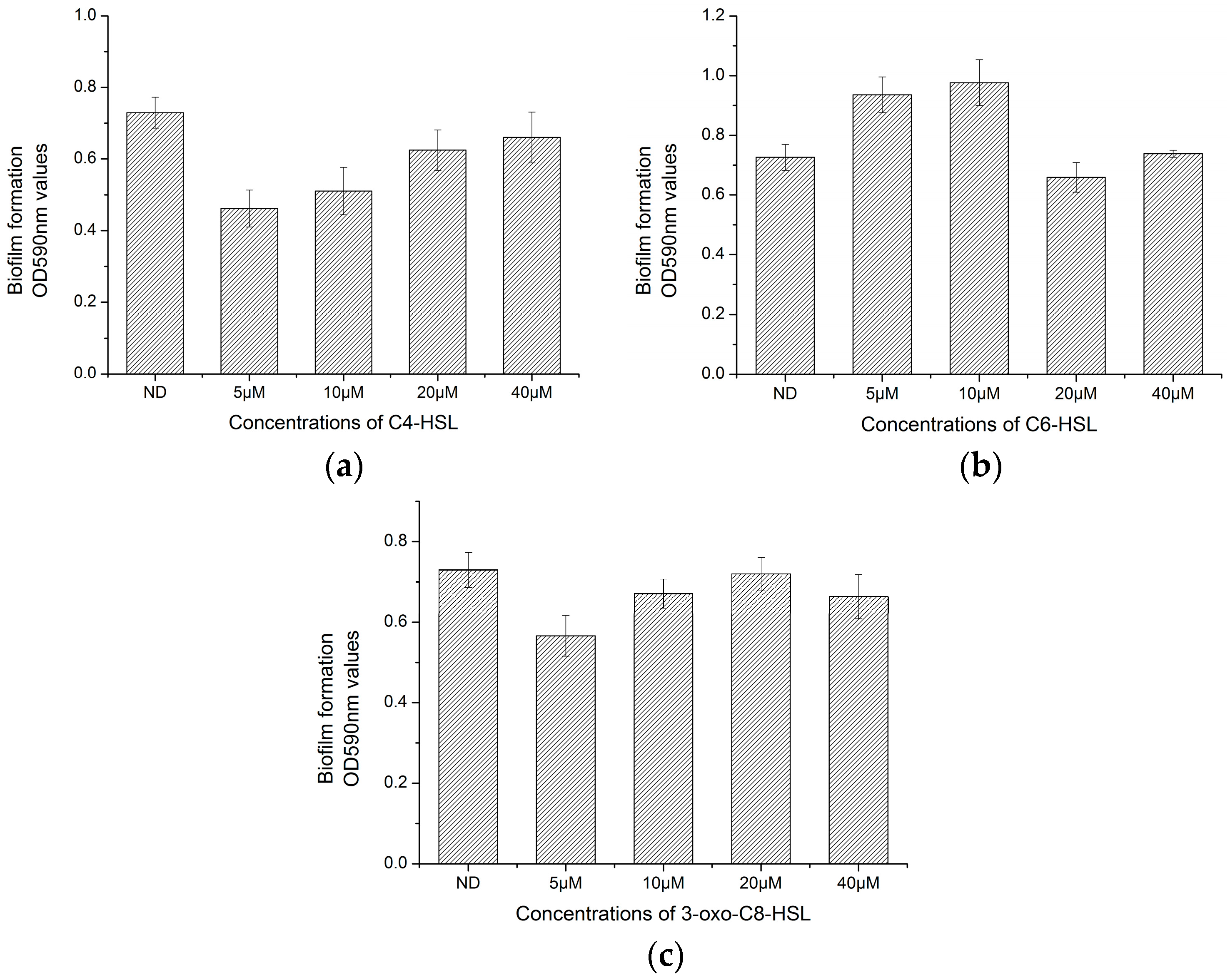
3.5. Effect of AHLs on Biofilm Formation of H. alvei H4
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Skandamis, P.N.; Nychas, G.J. Quorum sensing in the context of food microbiology. Appl. Environ. Microbiol. 2012, 78, 5473–5482. [Google Scholar] [CrossRef] [PubMed]
- Dourou, D.; Ammor, M.S.; Skandamis, P.N.; Nychas, G.J. Growth of Salmonella enteritidis and Salmonella typhimurium in the presence of quorum sensing signalling compounds produced by spoilage and pathogenic bacteria. Food Microbiol. 2011, 28, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.Y.; Yin, W.F.; Chan, K.G. Quorum sensing activity of Hafnia alvei isolated from packed food. Sensors 2014, 14, 6788–6796. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wu, H.; Zeng, M.; Liu, Z.; Wang, Y. The involvement of bacterial quorum sensing in the spoilage of refrigerated Litopenaeus vannamei. Int. J. Food Microbiol. 2015, 192, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gray, J.M.; Griffiths, M.W. Occurrence of proteolytic activity and N-acyl-homoserine lactone signals in the spoilage of aerobically chill-stored proteinaceous raw foods. J. Food Prot. 2006, 69, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Pinto, U.M.; De Souza, V.E.; Martins, M.L.; Vanetti, M.C.D. Detection of acylated homoserine lactones in gram-negative proteolytic psychrotrophic bacteria isolated from cooled raw milk. Food Control 2007, 18, 1322–1327. [Google Scholar] [CrossRef]
- Smith, D.; Wang, J.H.; Swatton, J.E.; Davenport, P.; Price, B.; Mikkelsen, H.; Stickland, H.; Nishikawa, K.; Gardiol, N.; Spring, D.R.; et al. Variations on a theme: diverse N-acyl homoserine lactone-mediated quorum sensing mechanisms in gram-negative bacteria. Sci. Prog. 2006, 89, 167–211. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.A.; Barnard, A.M.; Slater, H.; Simpson, N.J.; Salmond, G.P. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef] [PubMed]
- Popat, R.; Cornforth, D.M.; McNally, L.; Brown, S.P. Collective sensing and collective responses in quorum-sensing bacteria. J. R. Soc. Interface 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Rul, F.; Monnet, V. How microbes communicate in food: A review of signaling molecules and their impact on food quality. Curr. Opin. Food Sci. 2015, 2, 100–105. [Google Scholar] [CrossRef]
- Liu, Q.; Sun, J.; Pang, Y.; Jia, Z. Optimization of Processing Technology of Instant Sea Cucumber with Fuzzy Mathematic Comprehensive Evaluation by Response Surface Methodology and Exploration on Nutritional Value of Instant Sea Cucumber. Food Sci. Technol. Res. 2016, 22, 583–593. [Google Scholar] [CrossRef]
- Bai, A.J.; Rai, V.R. Bacterial quorum sensing and food industry. Compr. Rev. Food Sci. Food Saf. 2011, 10, 183–193. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Deak, E.; Tako, M.; Tserennadmid, R.; Petkovits, T.; Vagvolgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Micrbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Stanford, K.; McAllister, T.A.; Johnson, R.P.; Chen, J.; Hou, H.; Niu, Y.D. Biofilm Formation, Virulence Gene Profiles, and Antimicrobial Resistance of Nine Serogroups of Non-O157 Shiga Toxin—Producing Escherichia coli. Foodborne Pathog. Dis. 2016, 13, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Aye, A.M.; Bonnin-Jusserand, M.; Brian-Jaisson, F.; Ortalo-Magné, A.; Culioli, G.; Nevry, R.K.; Molmeret, M. Modulation of violacein production and phenotypes associated with biofilm by exogenous quorum sensing N-acylhomoserine lactones in the marine bacterium Pseudoalteromonas ulvae TC14. Microbiology 2015, 161, 2039–2051. [Google Scholar]
- Zhao, A.; Zhu, J.; Ye, X.; Ge, Y.; Li, J. Inhibition of biofilm development and spoilage potential of Shewanella baltica by quorum sensing signal in cell-free supernatant from Pseudomonas fluorescens. Int. J. Food Microbiol. 2016, 230, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Savini, V.; Santarelli, A.; Polilli, E.; Astolfi, D.; Pompilio, A.; Di Bonaventura, G.; Fazii, P.; D’Antonio, D. Hafnia alvei from the farm to the delivery room. Vet. Microbiol. 2013, 163, 202–203. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, J.B.; Christensen, A.B.; Flodgaard, L.R.; Nielsen, K.F.; Larsen, T.O.; Givskov, M.; Gram, L. Presence of acylated homoserine lactones (AHLs) and AHL-producing bacteria in meat and potential role of AHL in spoilage of meat. Appl. Environ. Microbiol. 2004, 70, 4293–4302. [Google Scholar] [CrossRef] [PubMed]
- Viana, E.S.; Campos, M.E.; Poce, A.R.; Mantovani, H.C.; Vanetti, M.C. Biofilm formation and acyl homoserine lactone production in Hafnia alvei isolated from raw milk. Biol. Res. 2009, 42, 427–436. [Google Scholar] [PubMed]
- Zhang, C.; Zhu, S.; Jatt, A.N.; Zeng, M. Characterization of N-acyl homoserine lactones (AHLs) producing bacteria isolated from vacuum-packaged refrigerated turbot (Scophthalmus maximus) and possible influence of exogenous AHLs on bacterial phenotype. J. Gen. Appl. Microbiol. 2016, 62, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Anbazhagan, D.; Mansor, M.; Yan, G.O.S.; Yusof, M.Y.M.; Hassan, H.; Sekaran, S.D. Detection of quorum sensing signal molecules and identification of an autoinducer synthase gene among biofilm forming clinical isolates of Acinetobacter spp. PLoS ONE 2012, 7, e36696. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.Y.; Khan, S.A.; Tee, K.K.; Kasim, N.H.A.; Yin, W.F.; Chan, K.G. Quorum sensing activity of Citrobacter amalonaticus L8A, a bacterium isolated from dental plaque. Sci. Rep. 2016, 6, 20702. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.W.; Koh, C.L.; Sam, C.K.; Yin, W.F.; Chan, K.G. Short chain N-acyl homoserine lactone production by soil isolate Burkholderia sp. strain A9. Sensors 2013, 13, 13217–13227. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; George, J.E.; McCarty, C.L.; Wendelken, S.C. Compliance analysis of phenylurea and related compounds in drinking water by liquid chromatography/electrospray ionization/mass spectrometry coupled with solid-phase extraction. J. Chromatogr. A 2006, 1134, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Okutsu, N.; Morohoshi, T.; Xie, X.; Kato, N.; Ikeda, T. Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems. Sensors 2015, 16, E44. [Google Scholar] [CrossRef] [PubMed]
- Packiavathy, I.A.; Sasikumar, P.; Pandian, S.K.; Veera Ravi, A. Prevention of quorum-sensing-mediated biofilm development and virulence factors production in Vibrio spp. by curcumin. Appl. Microbiol. Biotechnol. 2013, 97, 10177–10187. [Google Scholar] [CrossRef] [PubMed]
- Alhede, M.; Bjarnsholt, T.; Givskov, M.; Givskov, M. Pseudomonas aeruginosa biofilms: Mechanisms of immune evasion. Adv. Appl. Microbiol. 2014, 86, 1–40. [Google Scholar] [PubMed]
- Ammor, M.S.; Michaelidis, C.; Nychas, G.J.E. Insights into the role of quorum sensing in food spoilage. J. Food Prot. 2008, 71, 1510–1525. [Google Scholar] [CrossRef] [PubMed]
- Jay, J.M.; Vilai, J.P.; Hughes, M.E. Profile and activity of the bacterial biota of ground beef held from freshness to spoilage at 5–7 °C. Int. J. Food Microbiol. 2003, 81, 105–111. [Google Scholar] [CrossRef]
- Chung, M.S.; Lee, J.H.; Min, D.B. Effects of Pseudomonas putrifaciens and Acinetobacter spp. on the flavor quality of raw ground Beef. J. Food Sci. 2002, 67, 77–83. [Google Scholar] [CrossRef]
- Møretrø, T.; Sharifzadeh, S.; Langsrud, S.; Heir, E.; Rickard, A.H. Coaggregation between Rhodococcus and Acinetobacter strains isolated from the food industry. Can. J. Microbiol. 2015, 61, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Ortori, C.A.; Dubern, J.F.; Chhabra, S.R.; Cámara, M.; Hardie, K.; Williams, P.; Barrett, D.A. Simultaneous quantitative profiling of N-acyl-l-homoserine lactone and 2-alkyl-4 (1H)-quinolone families of quorum-sensing signaling molecules using LC-MS/MS. Anal. Bioanal. Chem. 2011, 399, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.B.; Riedel, K.; Eberl, L.; Flodgaard, L.R.; Molin, S.; Gram, L.; Givskov, M. Quorum-sensing-directed protein expression in Serratia proteamaculans B5a. Microbiology 2003, 149, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Khajanchi, B.K.; Sha, J.; Kozlova, E.V.; Erova, T.E.; Suarez, G.; Sierra, J.C.; Chopra, A.K. N-acylhomoserine lactones involved in quorum sensing control the type VI secretion system, biofilm formation, protease production, and in vivo virulence in a clinical isolate of Aeromonas hydrophila. Microbiology 2009, 155, 3518–3531. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Li, X.; Qi, Z.; Zhang, X.H.; Bossier, P. Detection of different quorum-sensing signal molecules in a virulent Edwardsiella tarda strain LTB-4. J. Appl. Microbiol. 2010, 108, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Lazazzera, B.A. Quorum sensing and starvation: Signals for entry into stationary phase. Curr. Opin. Microbiol. 2000, 3, 177–182. [Google Scholar] [CrossRef]
- Nackerdien, Z.E.; Keynan, A.; Bassler, B.L.; Lederberg, J.; Thaler, D.S. Quorum sensing influences Vibrio harveyi growth rates in a manner not fully accounted for by the marker effect of bioluminescence. PLoS ONE 2008, 3, e1671. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Cui, F.; Bai, F.; Zhao, G.; Li, J. Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.). Sensors 2016, 16, 1083. [Google Scholar] [CrossRef] [PubMed]
- Nievas, F.; Bogino, P.; Sorroche, F.; Giordano, W. Detection, characterization, and biological effect of quorum-sensing signaling molecules in peanut-nodulating bradyrhizobia. Sensors 2012, 12, 2851–2873. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.M.; Wu, L.J.; Xiao, Y.L.; Zhao, D.; Chen, Z.X.; Kang, M.; Xie, Y. Enhancing pili assembly and biofilm formation in Acinetobacter baumannii ATCC19606 using non-native acyl-homoserine lactones. BMC Microbiol. 2015, 15, 62. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, Q.; Guo, F.; Zhang, S.; Jiang, Q.; Yu, X. Responses of bacterial strains isolated from drinking water environments to N-acyl-l-homoserine lactones and their analogs during biofilm formation. Front. Environ. Sci. Eng. 2014, 8, 205–214. [Google Scholar] [CrossRef]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, H.-M.; Zhu, Y.-L.; Wang, J.-Y.; Jiang, F.; Qu, W.-Y.; Zhang, G.-L.; Hao, H.-S. Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber. Sensors 2017, 17, 772. https://doi.org/10.3390/s17040772
Hou H-M, Zhu Y-L, Wang J-Y, Jiang F, Qu W-Y, Zhang G-L, Hao H-S. Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber. Sensors. 2017; 17(4):772. https://doi.org/10.3390/s17040772
Chicago/Turabian StyleHou, Hong-Man, Yao-Lei Zhu, Jia-Ying Wang, Feng Jiang, Wen-Yan Qu, Gong-Liang Zhang, and Hong-Shun Hao. 2017. "Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber" Sensors 17, no. 4: 772. https://doi.org/10.3390/s17040772
APA StyleHou, H. -M., Zhu, Y. -L., Wang, J. -Y., Jiang, F., Qu, W. -Y., Zhang, G. -L., & Hao, H. -S. (2017). Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber. Sensors, 17(4), 772. https://doi.org/10.3390/s17040772