Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains, Plasmids and Growth Conditions
2.3. Animal Cell Lines and Culture Conditions
2.4. Mucin and Mannose Binding Assays
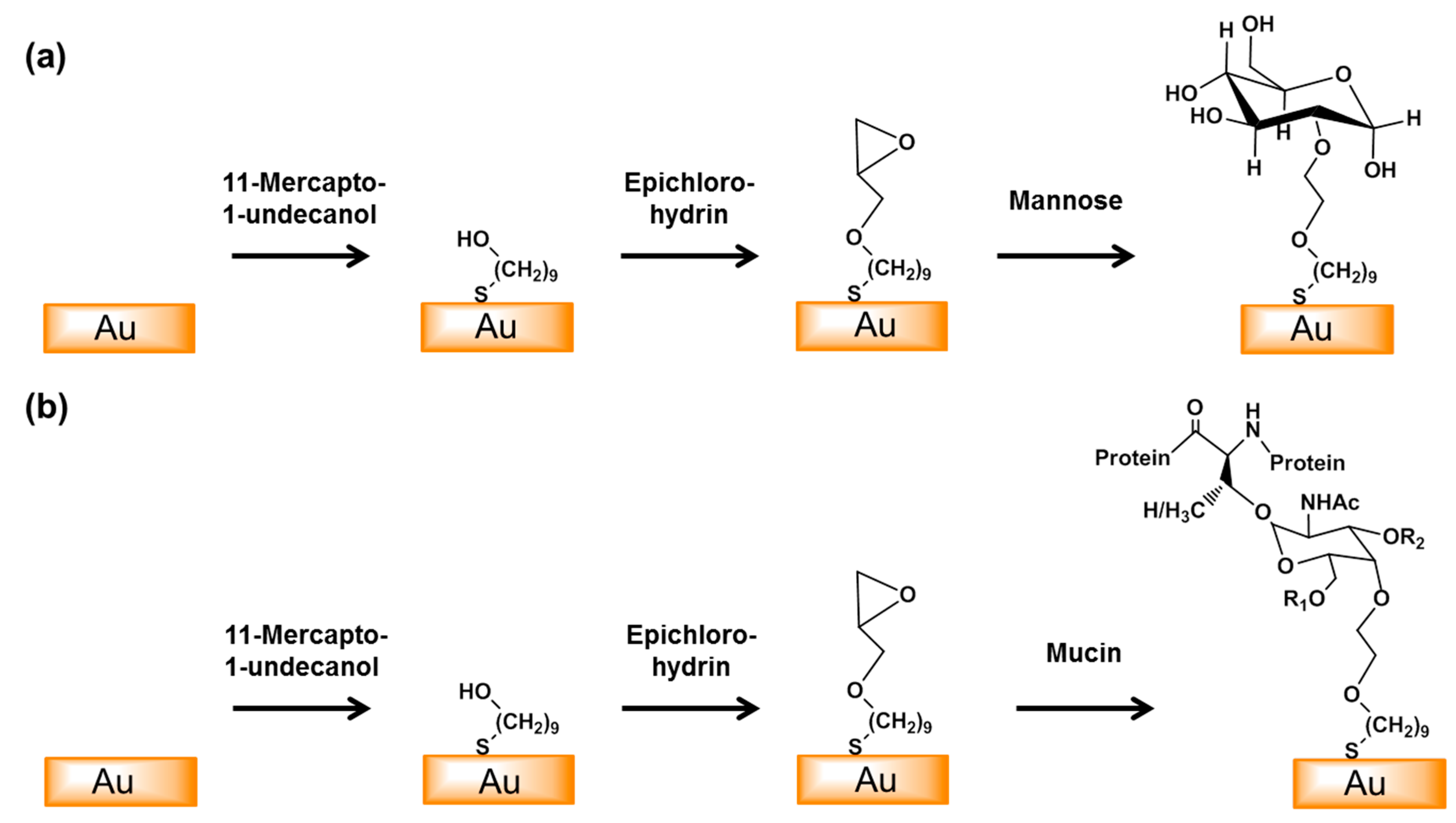
2.5. Functionalization of a Gold Electrode with Mucin and Mannose
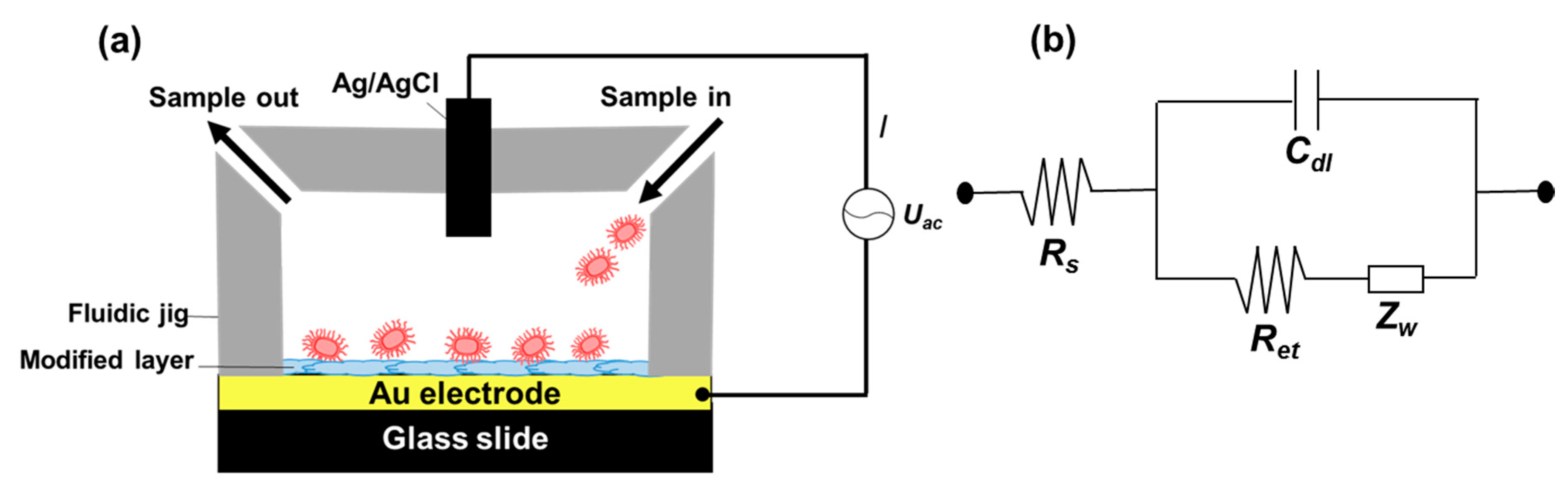
2.6. Electrical Measurement of Bacterial Adhesion to a Model Surface
2.7. Competition Assay
2.8. Statistical Analysis
3. Results and Discussion
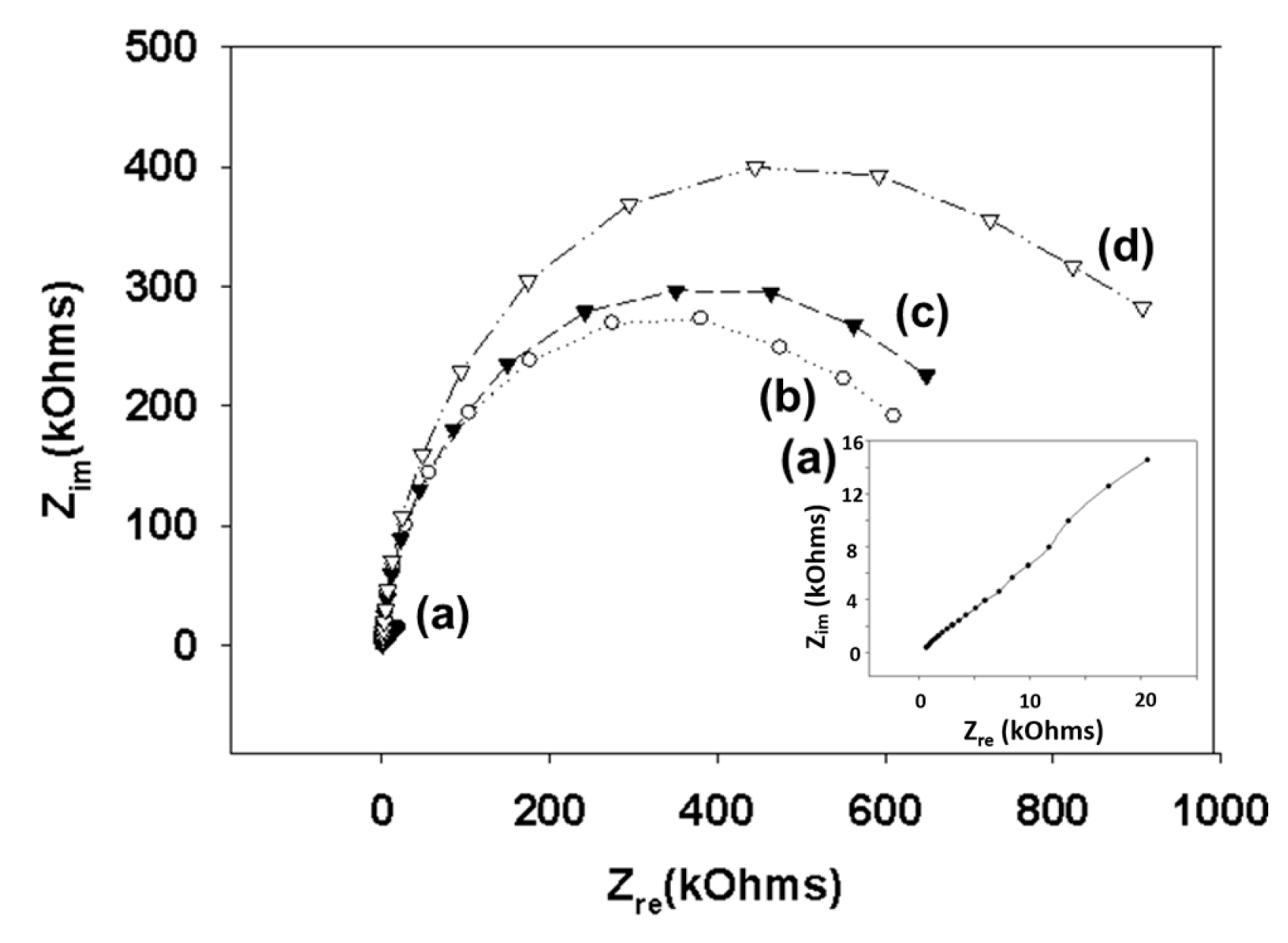
3.1. Characterization of Model Surface
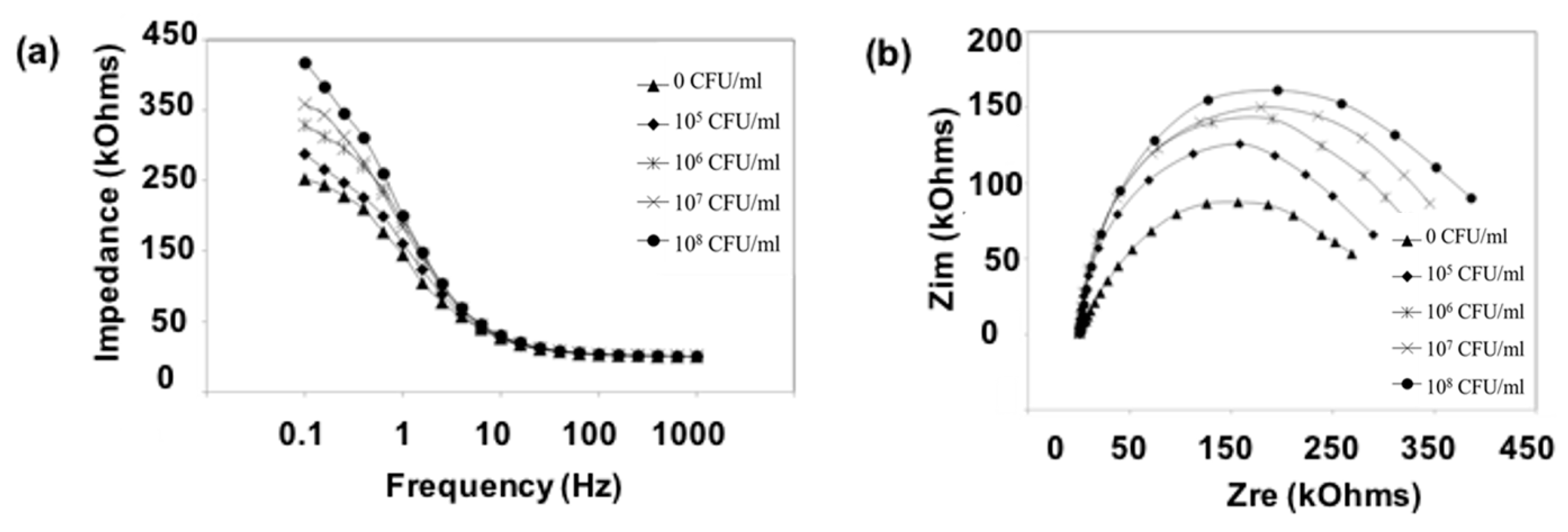
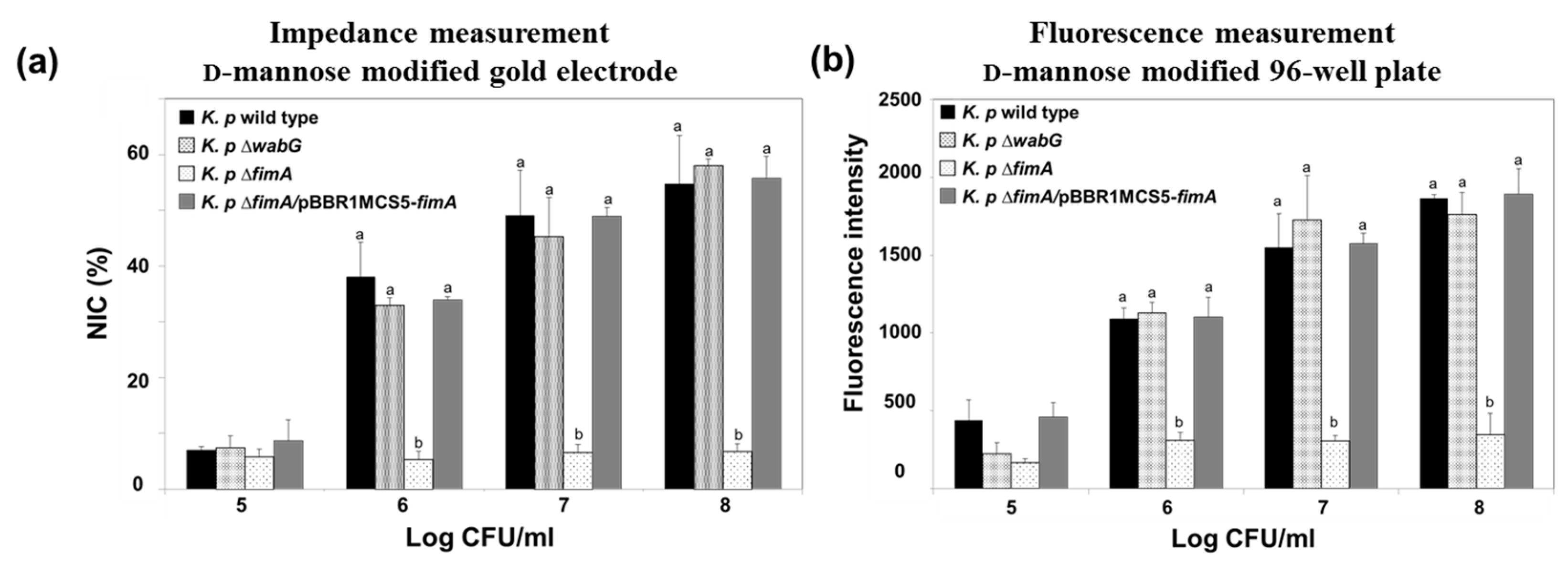
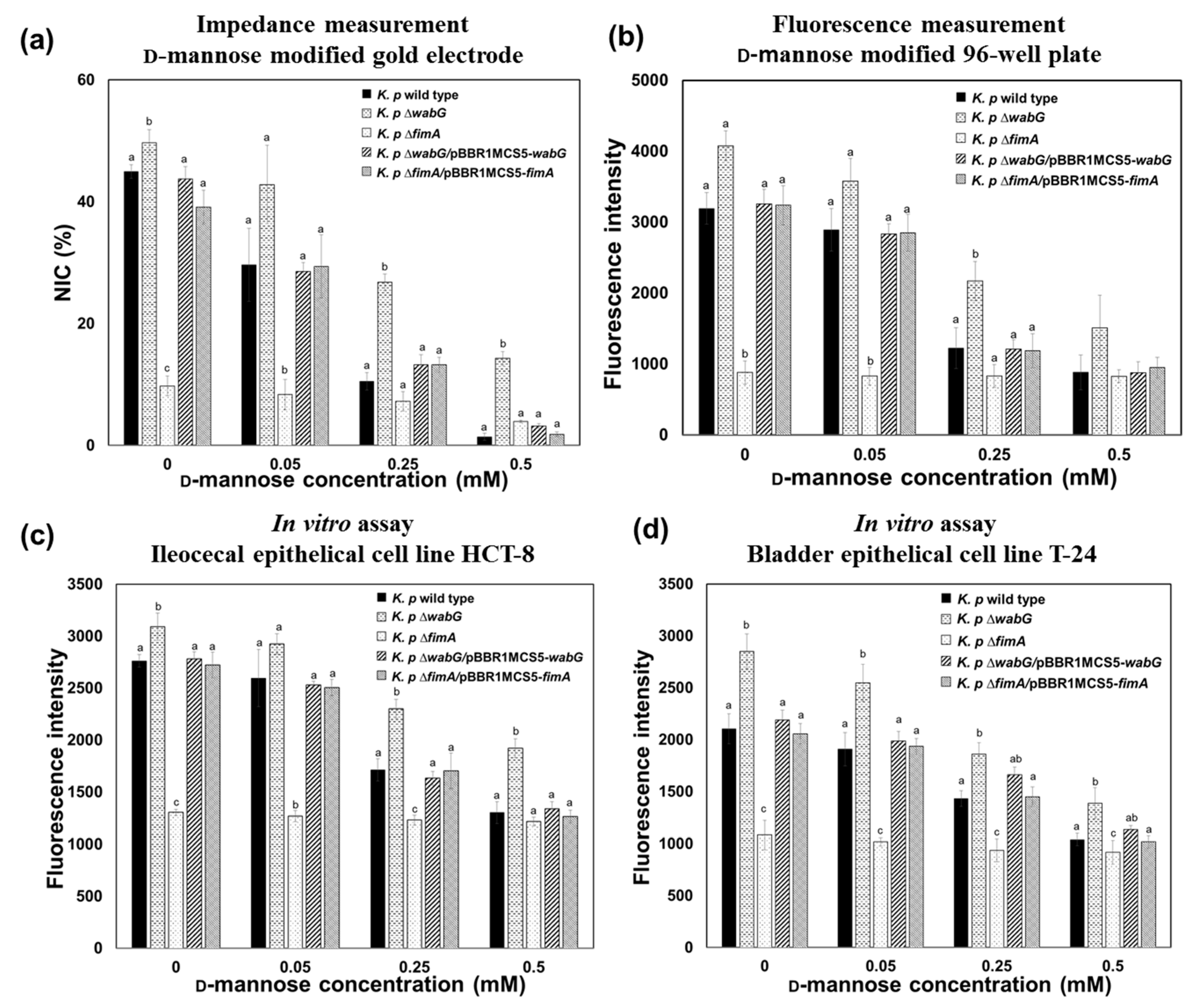
3.2. Adhesion Properties of K. pneumoniae on the Mannose-Modified Surface
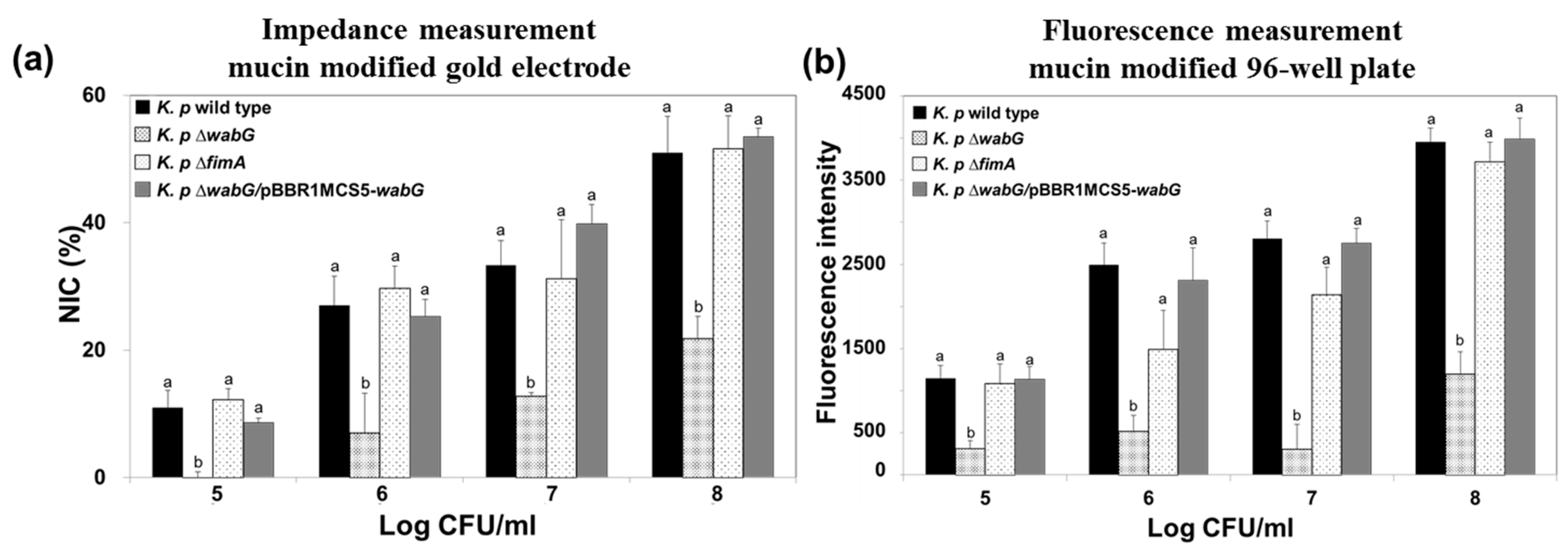
3.3. Adhesion Properties of K. pneumoniae on Mucin Modified Surfaces
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ariffin, H.; Navaratnam, P.; Mohamed, M.; Arasu, A.; Abdullah, W.A.; Lee, C.L.; Peng, L.H. Ceftazidime-resistant Klebsiella pneumoniae bloodstream infection in children with febrile neutropenia. Int. J. Infect. Dis. 2000, 4, 21–25. [Google Scholar] [CrossRef]
- Hennequin, C.; Forestier, C. Influence of capsule and extended-spectrum beta-lactamases encoding plasmids upon Klebsiella pneumoniae adhesion. Res. Microbiol. 2007, 158, 339–347. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Munn, V.P.; Highsmith, A.K.; Culver, D.H.; Hughes, J.M. The epidemiology of nosocomial infections caused by Klebsiella pneumoniae. Infect. Control 1985, 6, 68–74. [Google Scholar] [CrossRef]
- De Champs, C.; Sauvant, M.; Chanal, C.; Sirot, D.; Gazuy, N.; Malhuret, R.; Baguet, J.; Sirot, J. Prospective survey of colonization and infection caused by expanded-spectrum-beta-lactamase-producing members of the family Enterobacteriaceae in an intensive care unit. J. Clin. Microbiol. 1989, 27, 2887–2890. [Google Scholar]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar]
- Erridge, C.; Bennett-Guerrero, E.; Poxton, I.R. Structure and function of lipopolysaccharides. Microbes Infect. 2002, 4, 837–851. [Google Scholar] [CrossRef]
- Strettoti, C.W.; Ristuccia, P.A.; Cunha, B.A. Topics in clinical microbiology Klebsiella. Infect. Control 1984, 5, 343–348. [Google Scholar]
- Favre-Bonte, S.; Joly, B.; Forestier, C. Consequences of reduction of Klebsiella pneumoniae capsule expression on interactions of this bacterium with epithelial cells. Infect. Immun. 1999, 67, 554–561. [Google Scholar]
- Di Martino, P.; Livrelli, V.; Sirot, D.; Joly, B.; Darfeuille-Michaud, A. A new fimbrial antigen harbored by CAZ-5/SHV-4-producing Klebsiella pneumoniae strains involved in nosocomial infections. Infect. Immun. 1996, 64, 2266–2273. [Google Scholar]
- Duguid, J. Fimbriae and adhesive properties in Klebsiella strains. J. Gen. Microbiol. 1959, 21, 271–286. [Google Scholar] [CrossRef]
- Connell, I.; Agace, W.; Klemm, P.; Schembri, M.; Mărild, S.; Svanborg, C. Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract. Proc. Natl. Acad. Sci. USA 1996, 93, 9827–9832. [Google Scholar] [CrossRef]
- Fader, R.; Gondesen, K.; Tolley, B.; Ritchie, D.; Moller, P. Evidence that in vitro adherence of Klebsiella pneumoniae to ciliated hamster tracheal cells is mediated by type 1 fimbriae. Infect. Immun. 1988, 56, 3011–3013. [Google Scholar]
- Hornick, D.; Dayton, C.; Bedell, G.; Fick, R. Nontuberculous mycobacterial lung disease. Substantiation of a less aggressive approach. CHEST J. 1988, 93, 550–555. [Google Scholar] [CrossRef]
- Tarkkanen, A.-M.; Virkola, R.; Clegg, S.; Korhonen, T.K. Binding of the type 3 fimbriae of Klebsiella pneumoniae to human endothelial and urinary bladder cells. Infect. Immun. 1997, 65, 1546–1549. [Google Scholar]
- Hultgren, S.J.; Abraham, S.; Caparon, M.; Falk, P.; Geme, J.W.S.; Normark, S. Pilus and nonpilus bacterial adhesins: Assembly and function in cell recognition. Cell 1993, 73, 887–901. [Google Scholar] [CrossRef]
- Disney, M.D.; Seeberger, P.H. The use of carbohydrate microarrays to study carbohydrate-cell interactions and to detect pathogens. Chem. Biol. 2004, 11, 1701–1707. [Google Scholar] [CrossRef]
- Huynh, D.T.N.; Kim, A.-Y.; Seol, I.-H.; Jung, S.; Lim, M.-C.; Lee, J.-A.; Jo, M.-R.; Choi, S.-J.; Kim, B.; Lee, J. Inactivation of the virulence factors from 2,3-butanediol-producing Klebsiella pneumoniae. Appl. Microbiol. Biotechnol. 2015, 99, 9427–9438. [Google Scholar] [CrossRef]
- Sahly, H.; Podschun, R.; Oelschlaeger, T.A.; Greiwe, M.; Parolis, H.; Hasty, D.; Kekow, J.; Ullmann, U.; Ofek, I.; Sela, S. Capsule impedes adhesion to and invasion of epithelial cells by Klebsiella pneumoniae. Infect. Immun. 2000, 68, 6744–6749. [Google Scholar] [CrossRef]
- Linden, S.; Sutton, P.; Karlsson, N.; Korolik, V.; McGuckin, M. Mucins in the mucosal barrier to infection. Mucosal. Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef]
- Dreux, N.; Denizot, J.; Martinez-Medina, M.; Mellmann, A.; Billig, M.; Kisiela, D.; Chattopadhyay, S.; Sokurenko, E.; Neut, C.; Gower-Rousseau, C. Point mutations in FimH adhesin of Crohn’s disease-associated adherent-invasive Escherichia coli enhance intestinal inflammatory response. PLoS Pathog. 2013, 9, e1003141. [Google Scholar] [CrossRef]
- Jung, S.-G.; Jang, J.-H.; Kim, A.-Y.; Lim, M.-C.; Kim, B.; Lee, J.; Kim, Y.-R. Removal of pathogenic factors from 2,3-butanediol-producing Klebsiella species by inactivating virulence-related wabG gene. Appl. Microbiol. Biotechnol. 2013, 97, 1997–2007. [Google Scholar] [CrossRef]
- Ryan, P.A.; Pancholi, V.; Fischetti, V.A. Group A streptococci bind to mucin and human pharyngeal cells through sialic acid-containing receptors. Infect. Immun. 2001, 69, 7402–7412. [Google Scholar] [CrossRef]
- Vishwanath, S.; Ramphal, R. Adherence of Pseudomonas aeruginosa to human tracheobronchial mucin. Infect. Immun. 1984, 45, 197–202. [Google Scholar]
- Joung, C.-K.; Kim, H.-N.; Im, H.-C.; Kim, H.-Y.; Oh, M.-H.; Kim, Y.-R. Ultra-sensitive detection of pathogenic microorganism using surface-engineered impedimetric immunosensor. Sens. Actuators B Chem. 2012, 161, 824–831. [Google Scholar] [CrossRef]
- Shi, L.; Ardehali, R.; Caldwell, K.D.; Valint, P. Mucin coating on polymeric material surfaces to suppress bacterial adhesion. Colloids Surf. B Biointerfaces 2000, 17, 229–239. [Google Scholar] [CrossRef]
- Shen, Z.; Huang, M.; Xiao, C.; Zhang, Y.; Zeng, X.; Wang, P.G. Nonlabeled quartz crystal microbalance biosensor for bacterial detection using carbohydrate and lectin recognitions. Anal. Chem. 2007, 79, 2312–2319. [Google Scholar] [CrossRef]
- Lofas, S. Dextran modified self-assembled monolayer surfaces for use in biointeraction analysis with surface plasmon resonance. Pure Appl. Chem. 1995, 67, 829–834. [Google Scholar] [CrossRef]
- Yang, L.; Bashir, R. Electrical/electrochemical impedance for rapid detection of foodborne pathogenic bacteria. Biotechnol. Adv. 2008, 26, 135–150. [Google Scholar] [CrossRef]
- Bard, A.J.; Faulkner, L.R.; Leddy, J.; Zoski, C.G. Electrochemical Methods: Fundamentals and Applications; Wiley: New York, NY, USA, 1980; Volume 2. [Google Scholar]
- Gawad, S.; Cheung, K.; Seger, U.; Bertsch, A.; Renaud, P. Dielectric spectroscopy in a micromachined flow cytometer: Theoretical and practical considerations. Lab Chip 2004, 4, 241–251. [Google Scholar] [CrossRef]
- Schroll, C.; Barken, K.B.; Krogfelt, K.A.; Struve, C. Role of type 1 and type 3 fimbriae in Klebsiella pneumoniae biofilm formation. BMC Microbiol. 2010, 10, 179. [Google Scholar] [CrossRef]
- Neeser, J.-R.; Koellreutter, B.; Wuersch, P. Oligomannoside-type glycopeptides inhibiting adhesion of Escherichia coli strains mediated by type 1 pili: Preparation of potent inhibitors from plant glycoproteins. Infect. Immun. 1986, 52, 428–436. [Google Scholar]
- Struve, C.; Krogfelt, K.A. Role of capsule in Klebsiella pneumoniae virulence: Lack of correlation between in vitro and in vivo studies. FEMS Microbiol. Lett. 2003, 218, 149–154. [Google Scholar] [CrossRef]
- Matatov, R.; Goldhar, J.; Skutelsky, E.; Sechter, I.; Perry, R.; Podschun, R.; Sahly, H.; Thankavel, K.; Abraham, S.N.; Ofek, I. Inability of encapsulated Klebsiella pneumoniae to assemble functional type 1 fimbriae on their surface. FEMS Microbiol. Lett. 1999, 179, 123–130. [Google Scholar] [CrossRef]
- Schild, S.; Lamprecht, A.-K.; Fourestier, C.; Lauriano, C.M.; Klose, K.E.; Reidl, J. Characterizing lipopolysaccharide and core lipid A mutant O1 and O139 Vibrio cholerae strains for adherence properties on mucus-producing cell line HT29-Rev MTX and virulence in mice. Int. J. Med. Microbiol. 2005, 295, 243–251. [Google Scholar] [CrossRef]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huynh, D.T.N.; Kim, A.-Y.; Kim, Y.-R. Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces. Sensors 2017, 17, 1406. https://doi.org/10.3390/s17061406
Huynh DTN, Kim A-Y, Kim Y-R. Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces. Sensors. 2017; 17(6):1406. https://doi.org/10.3390/s17061406
Chicago/Turabian StyleHuynh, Duyen Thi Ngoc, Ah-Young Kim, and Young-Rok Kim. 2017. "Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces" Sensors 17, no. 6: 1406. https://doi.org/10.3390/s17061406
APA StyleHuynh, D. T. N., Kim, A. -Y., & Kim, Y. -R. (2017). Identification of Pathogenic Factors in Klebsiella pneumoniae Using Impedimetric Sensor Equipped with Biomimetic Surfaces. Sensors, 17(6), 1406. https://doi.org/10.3390/s17061406