Relationship between Remote Sensing Data, Plant Biomass and Soil Nitrogen Dynamics in Intensively Managed Grasslands under Controlled Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Soil Sampling
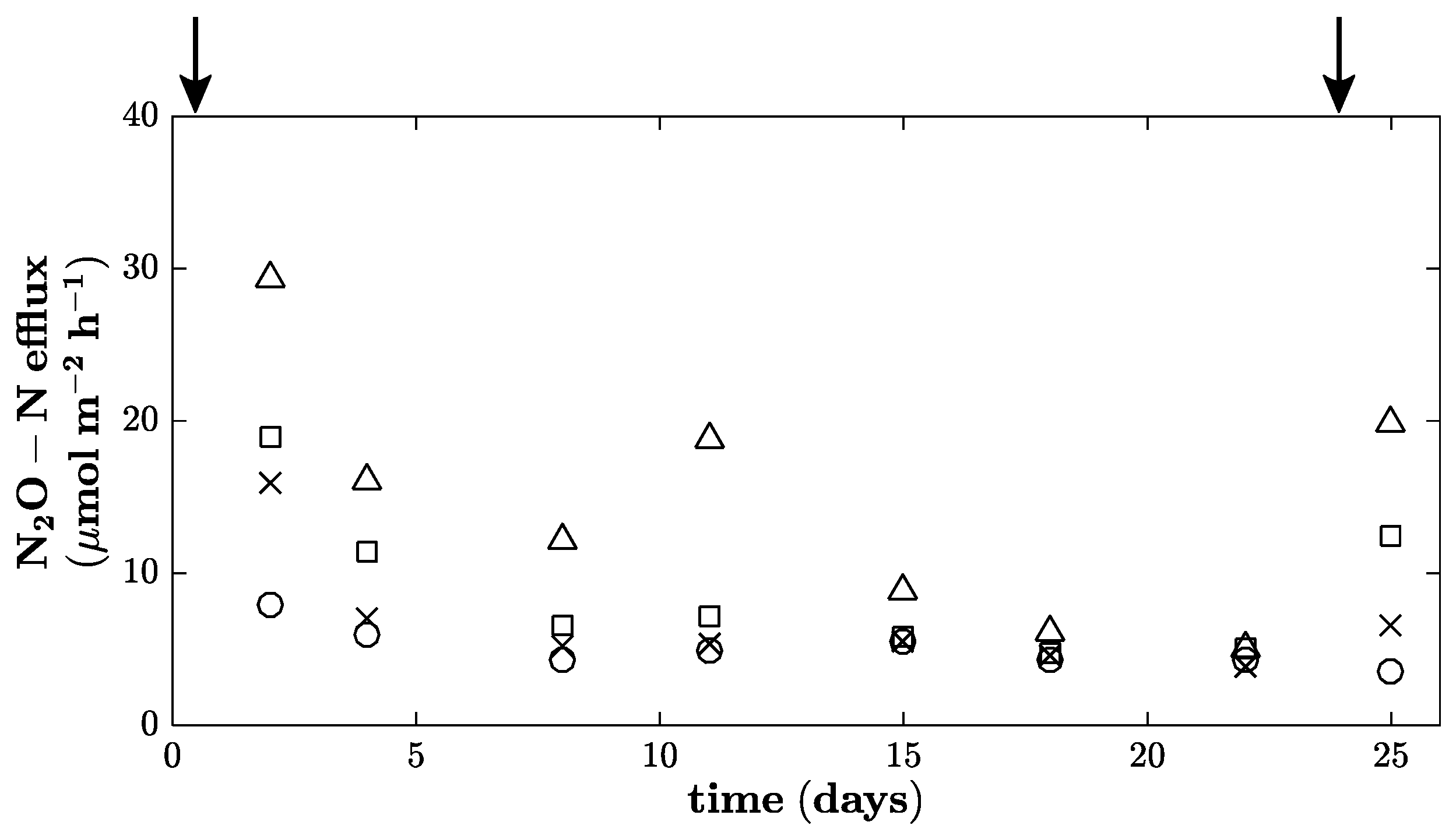
2.2. Gas Flux Measurements
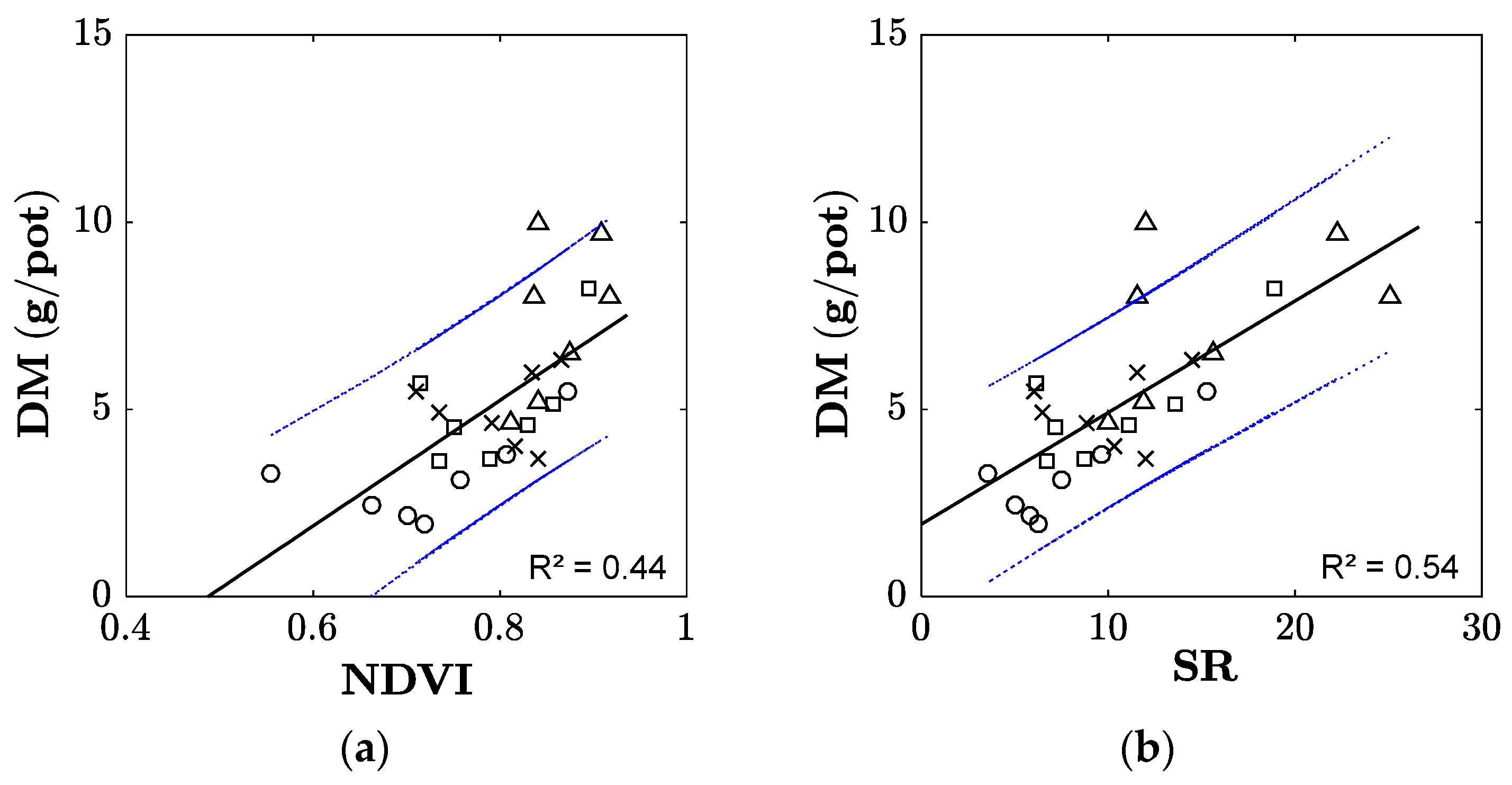
2.3. Narrow-Band Vegetation Indices
2.4. Soil Analysis
2.5. Statistical Analysis
3. Results
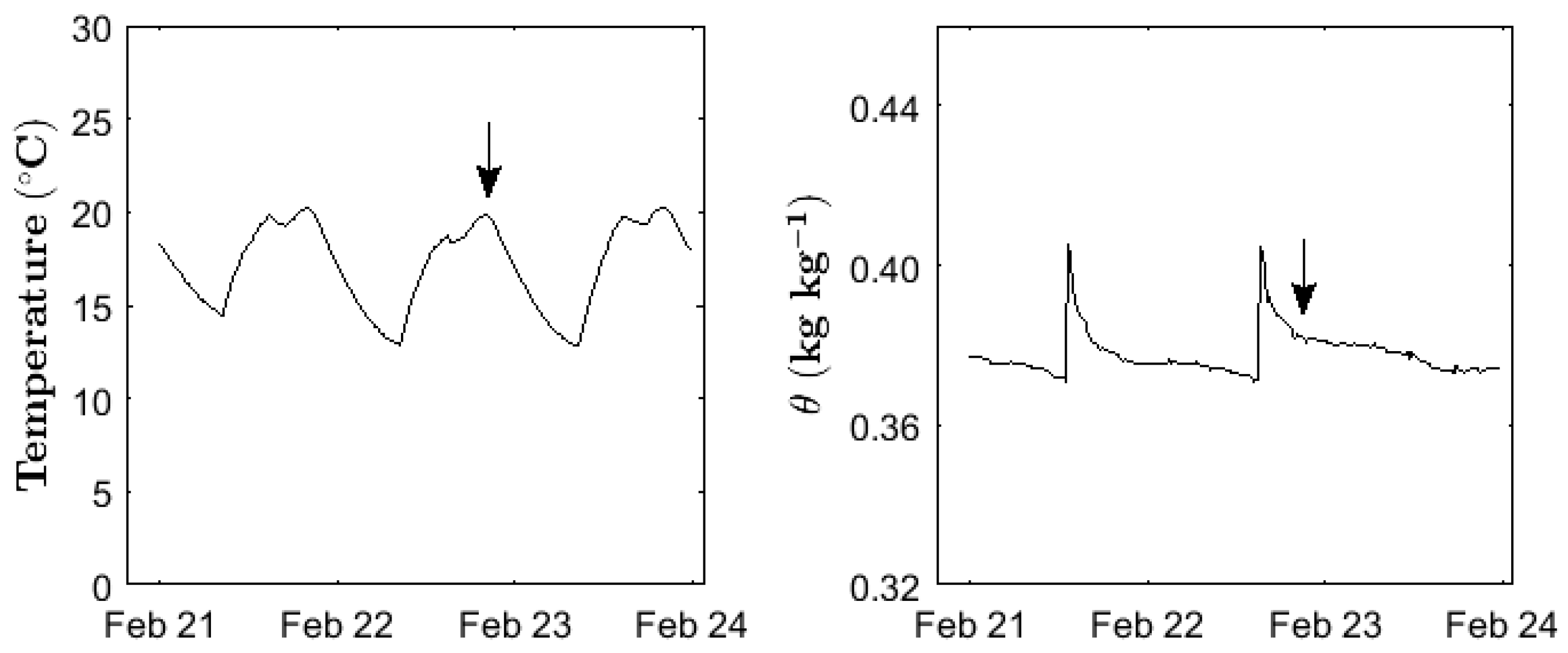
3.1. Temporal Dynamics of Soil Moisture and Temperature
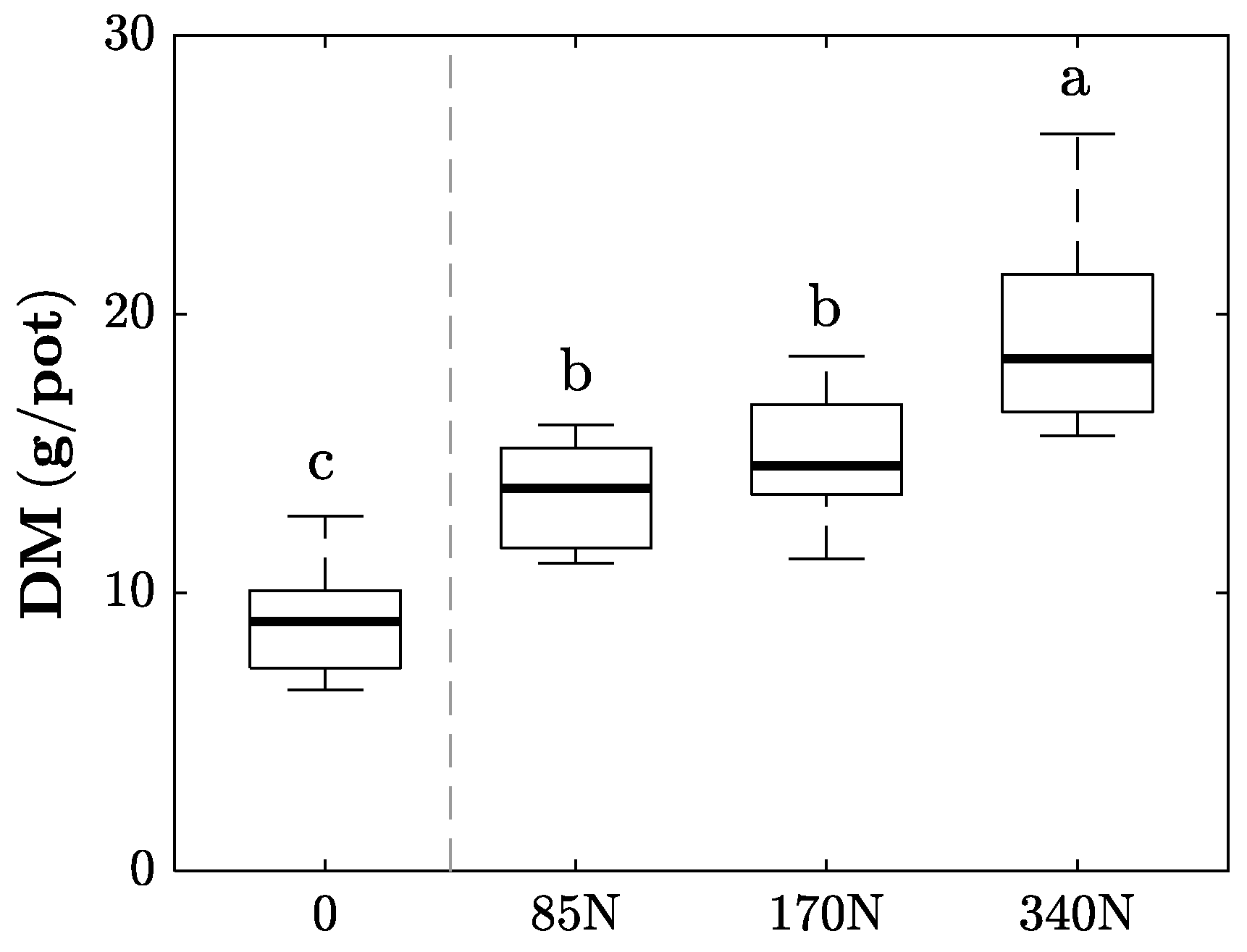
3.2. Plant Biomass Development
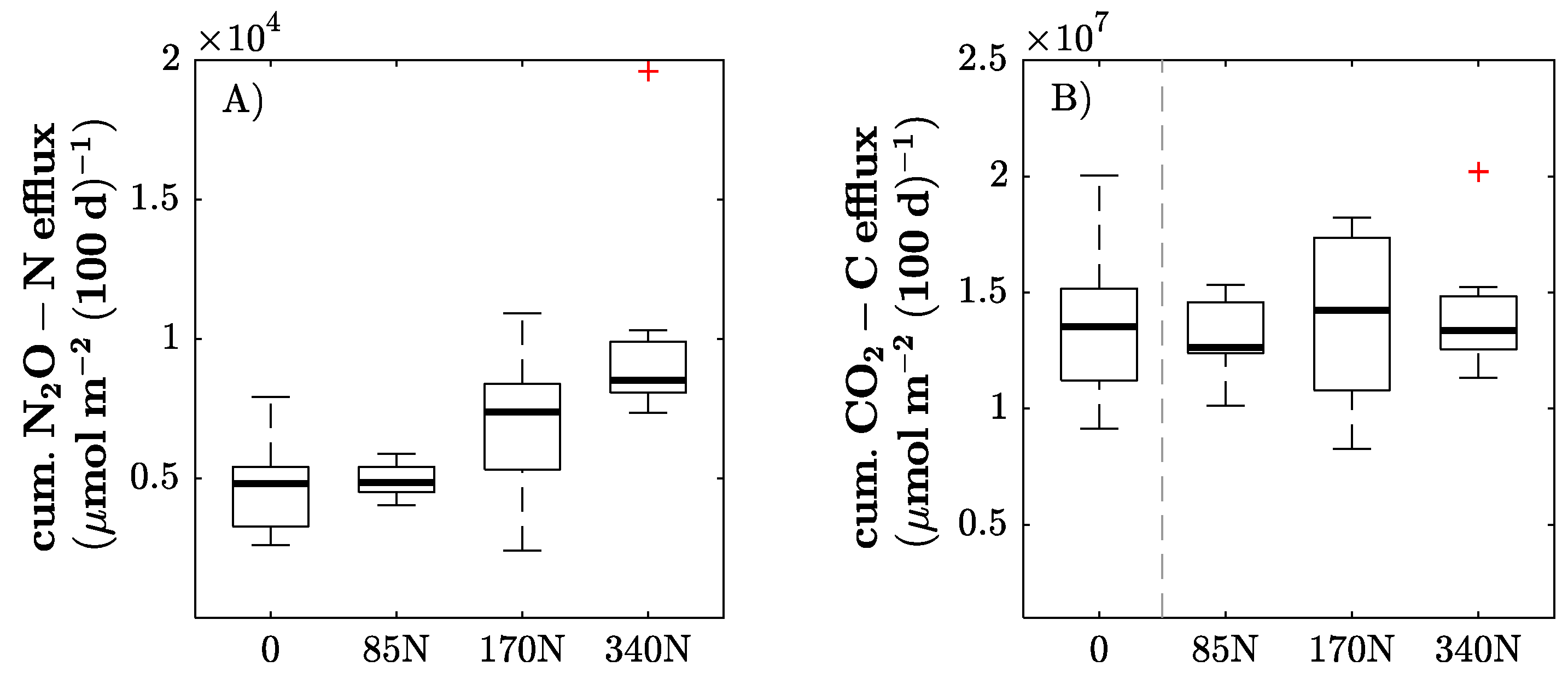
3.3. Soil C and N Dynamics
4. Discussion
4.1. Sensing Plant Traits
4.2. Soil C and N Dynamics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Breustedt, G.; Latacz-Lohmann, U.; Tiedemann, T. Organic or conventional? Optimal dairy farming technology under the EU milk quota system and organic subsidies. Food Policy 2011, 36, 223–229. [Google Scholar] [CrossRef]
- Van Zanten, B.T.; Verburg, P.H.; Espinosa, M.; Gomez-Y-Paloma, S.; Galimberti, G.; Kantelhardt, J.; Kapfer, M.; Lefebvre, M.; Manrique, R.; Piorr, A.; et al. European agricultural landscapes, common agricultural policy and ecosystem services: A review. Agron. Sustain. Dev. 2014, 34, 309–325. [Google Scholar] [CrossRef]
- Harrison, R.; Webb, J. A review of the effect of N fertilizer type on gaseous emissions. Adv. Agron. 2001, 73, 65–108. [Google Scholar]
- Bristow, A.W.; Jarvis, S.C. Effects of grazing and nitrogen fertiliser on the soil microbial biomass under permanent pasture. J. Sci. Food Agric. 1991, 54, 9–21. [Google Scholar] [CrossRef]
- Knorr, M.; Frey, S.D.; Curtis, P.S. Nitrogen additions and litter decomposition: A meta-analysis. Ecology 2005, 86, 3252–3257. [Google Scholar] [CrossRef]
- Knoblauch, C.; Watson, C.; Becker, R.; Berendonk, C.; Wichern, F. Change of ergosterol content after inorganic N fertilizer application does not affect short-term C and N mineralization patterns in a grassland soil. Appl. Soil Ecol. 2017, 111, 57–64. [Google Scholar] [CrossRef]
- Capolupo, A.; Kooistra, L.; Berendonk, C.; Boccia, L.; Suomalainen, J. Estimating Plant Traits of Grasslands from UAV-Acquired Hyperspectral Images: A Comparison of Statistical Approaches. ISPRS Int. J. Geoinf. 2015, 4, 2792–2820. [Google Scholar] [CrossRef]
- Fava, F.; Colombo, R.; Bocchi, S.; Meroni, M.; Sitzia, M.; Fois, N.; Zucca, C. Identification of hyperspectral vegetation indices for Mediterranean pasture characterization. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 233–243. [Google Scholar] [CrossRef]
- Hollberg, J.; Schellberg, J. Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices. Remote Sens. 2017, 9, 81. [Google Scholar] [CrossRef]
- Hansen, P.M.; Schjoerring, J.K. Reflectance measurement of canopy biomass and nitrogen status in wheat crops using normalized difference vegetation indices and partial least squares regression. Remote Sens. Environ. 2003, 86, 542–553. [Google Scholar] [CrossRef]
- Carvalho, S.; van der Putten, W.H.; Hol, W.H.G. The Potential of Hyperspectral Patterns of Winter Wheat to Detect Changes in Soil Microbial Community Composition. Front. Plant Sci. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lesschen, J.P.; Velthof, G.L.; De Vries, W.; Kros, J. Differentiation of nitrous oxide emission factors for agricultural soils. Environ. Pollut. 2011, 159, 3215–3222. [Google Scholar] [CrossRef] [PubMed]
- Rafique, R.; Hennessy, D.; Kiely, G. Nitrous Oxide Emission from Grazed Grassland Under Different Management Systems. Ecosystems 2011, 14, 563–582. [Google Scholar] [CrossRef]
- Possoch, M.; Bieker, S.; Hoffmeister, D.; Bolten, A.A.; Schellberg, J.; Bareth, G. Multi-Temporal crop surface models combined with the rgb vegetation index from UAV-based images for forage monitoring in grassland. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, 991–998. [Google Scholar] [CrossRef]
- Wilke, B.-M. Determination of Chemical and Physical Soil Properties. In Monitoring and Assessing Soil Bioremediation; Springer: Berlin/Heidelberg, Germany, 2005; pp. 47–95. [Google Scholar]
- FAO ISSS. World Reference Base for Soil Resources; World Soil Resourse Report; Bundesanstalt für Geowissenschaften und Rohstoffe (BGR): Hannover, Germany, 2007. [Google Scholar]
- Topp, G.C.; Davis, J.L.; Bailey, W.G.; Zebchuk, W.D. The measurement of soil water content using a portable TDR hand probe. Can. J. Soil Sci. 1984, 64, 313–321. [Google Scholar] [CrossRef]
- Rochette, P.; Ellert, B.; Gregorich, E.G.; Desjardins, R.L.; Pattey, E.; Lessard, R.; Johnson, B.G. Description of a dynamic closed chamber for measuring soil respiration and its comparison with other techniques. Can. J. Soil Sci. 1997, 77, 195–203. [Google Scholar] [CrossRef]
- Velthof, G.L. Nitrous oxide fluxes from grassland in the Netherlands: I. Statistical analysis of flux-chamber measurements. Eur. J. Agron. 1995, 46, 533–540. [Google Scholar] [CrossRef]
- Rouse, W.; Haas, R.H.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS; NASA: Washington, DC, USA, 1974.
- Barmeier, G.; Schmidhalter, U. High-throughput phenotyping of wheat and barley plants grown in single or few rows in small plots using active and passive spectral proximal sensing. Sensors 2016, 16, 1860. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. Microbial biomass measurements in forest soils: The use of the chloroform fumigation-incubation method in strongly acid soils. Soil Biol. Biochem. 1987, 19, 697–702. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Dusseux, P.; Hubert-Moy, L.; Corpetti, T.; Vertès, F. Evaluation of SPOT imagery for the estimation of grassland biomass. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 72–77. [Google Scholar] [CrossRef]
- Poorter, H.; Fiorani, F.; Stitt, M.; Schurr, U.; Finck, A.; Gibon, Y.; Usadel, B.; Munns, R.; Atkin, O.K.; Tardieu, F.; et al. The art of growing plants for experimental purposes: A practical guide for the plant biologist. Funct. Plant Biol. 2012, 39, 821–838. [Google Scholar] [CrossRef]
- Shcherbak, I.; Millar, N.; Robertson, G.P. Global metaanalysis of the nonlinear response of soil nitrous oxide (N2O) emissions to fertilizer nitrogen. Proc. Natl. Acad. Sci. USA 2014, 111, 9199–9204. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Dobbie, K.E.; Thorman, R.; Watson, C.J.; Chadwick, D.R.; Yamulki, S.; Ball, B.C. The effect of N fertilizer forms on nitrous oxide emissions from UK arable land and grassland. Nutr. Cycl. Agroecosyst. 2012, 93, 127–149. [Google Scholar] [CrossRef]
- Dobbie, K.; Smith, K. Nitrous oxide emission factors for agricultural soils in Great Britain: The impact of soil water filled pore space and other controlling variables. Glob. Chang. Biol. 2003, 99, 204–218. [Google Scholar] [CrossRef]
- Glatzel, S.; Stahr, K. Methane and nitrous oxide exchange in differently fertilised grassland in southern Germany. Plant Soil 2001, 231, 21–35. [Google Scholar] [CrossRef]
- Hoffmann, C.; Anger, M.; Kühbauch, W. N2O-Freisetzung auf gemähtem Dauergrünland in Abhängigkeit von Standort und N-Düngung. J. Agron. Crop Sci. 2001, 187, 153–159. [Google Scholar] [CrossRef]
- Del Grosso, S.J.; Wirth, T.; Ogle, S.; Parton, W.J. Estimating Agricultural Nitrous Oxide Emissions. EOS Trans. Am. Geophys. Union 2008, 89, 529–540. [Google Scholar] [CrossRef]
- Jungkunst, H.F.; Freibauer, A.; Neufeldt, H.; Bareth, G. Nitrous oxide emissions from agricultural land use in Germany—A synthesis of available annual field data. J. Plant Nutr. Soil Sci. 2006, 169, 341–351. [Google Scholar] [CrossRef]
- Venterea, R.T.; Halvorson, A.D.; Kitchen, N.; Liebig, M.A.; Cavigelli, M.A.; Del Grosso, S.J.; Motavalli, P.P.; Nelson, K.A.; Spokas, K.A.; Singh, B.P.; et al. Challenges and opportunities for mitigating nitrous oxide emissions from fertilized cropping systems. Front. Ecol. Environ. 2012, 10, 562–570. [Google Scholar] [CrossRef]
- Joergensen, R.G. Ergosterol and microbial biomass in the rhizosphere of grassland soils. Soil Biol. Biochem. 2000, 32, 647–652. [Google Scholar] [CrossRef]
- Wright, C.J.; Coleman, D.C. Cross-site comparison of soil microbial biomass, soil nutrient status, and nematode trophic groups. Pedobiologia 2000, 44, 2–23. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Schwenke, G.D.; Van Zwieten, L. Impact of agricultural inputs on soil organisms—A review. Aust. J. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef]
- Geisseler, D.; Lazicki, P.A.; Scow, K.M. Mineral nitrogen input decreases microbial biomass in soils under grasslands but not annual crops. Appl. Soil Ecol. 2016, 106, 1–10. [Google Scholar] [CrossRef]
- De Vries, F.T.; Bloem, J.; van Eekeren, N.; Brusaard, L.; Hoffland, E. Fungal biomass in pastures increases with age and reduced N input. Soil Biol. Biochem. 2007, 39, 1620–1630. [Google Scholar] [CrossRef]
- Jones, D.L.; Shannon, D.; Murphy, D.V.; Farrar, J. Role of dissolved organic nitrogen (DON) in soil N cycling in grassland soils. Soil Biol. Biochem. 2004, 36, 749–756. [Google Scholar] [CrossRef]
- Wichern, F.; Lobe, I.; Amelung, W.; Müller, T.; Joergensen, R.G.; Buerkert, A. Changes in amino acid enantiomers and microbial performance in soils from a subtropical mountain oasis in Oman abandoned for different periods. Biol. Fertil. Soils 2004, 39, 398–406. [Google Scholar] [CrossRef]
- Cookson, W.R.; Murphy, D.V. Quantifying the contribution of dissolved organic matter to soil nitrogen cycling using 15 N isotopic pool dilution. Soil Biol. Biochem. 2004, 36, 2097–2100. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Index | Formula | Source |
|---|---|---|
| NDVI | [20] | |
| SR | [21] |
| Treatment | MBC (µg (g−1 soil)) | MBN (µg (g−1 soil)) | Inorg. N (µg (g−1 soil)) | EOC (µg (g−1 soil)) | EON (µg (g−1 soil)) |
|---|---|---|---|---|---|
| Control | 602 ± 82 a | 121 ± 19 a | 8.9 ± 1.9 b | 71.8 ± 6.6 a | 22.8 ± 4.3 b |
| 85 N | 608 ± 83 a | 124 ± 17 a | 10.0 ± 1.2 ab | 76.6 ± 5.8 a | 25.8 ± 4.2 ab |
| 170 N | 559 ± 71 a | 109 ± 19 a | 9.6 ± 2.1 ab | 74.0 ± 13.7 a | 25.6 ± 4.7 ab |
| 340 N | 547 ± 80 a | 106 ± 16 a | 12.8 ± 3.1 a | 74.9 ± 11.7 a | 34.5 ± 9.4 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knoblauch, C.; Watson, C.; Berendonk, C.; Becker, R.; Wrage-Mönnig, N.; Wichern, F. Relationship between Remote Sensing Data, Plant Biomass and Soil Nitrogen Dynamics in Intensively Managed Grasslands under Controlled Conditions. Sensors 2017, 17, 1483. https://doi.org/10.3390/s17071483
Knoblauch C, Watson C, Berendonk C, Becker R, Wrage-Mönnig N, Wichern F. Relationship between Remote Sensing Data, Plant Biomass and Soil Nitrogen Dynamics in Intensively Managed Grasslands under Controlled Conditions. Sensors. 2017; 17(7):1483. https://doi.org/10.3390/s17071483
Chicago/Turabian StyleKnoblauch, Christoph, Conor Watson, Clara Berendonk, Rolf Becker, Nicole Wrage-Mönnig, and Florian Wichern. 2017. "Relationship between Remote Sensing Data, Plant Biomass and Soil Nitrogen Dynamics in Intensively Managed Grasslands under Controlled Conditions" Sensors 17, no. 7: 1483. https://doi.org/10.3390/s17071483
APA StyleKnoblauch, C., Watson, C., Berendonk, C., Becker, R., Wrage-Mönnig, N., & Wichern, F. (2017). Relationship between Remote Sensing Data, Plant Biomass and Soil Nitrogen Dynamics in Intensively Managed Grasslands under Controlled Conditions. Sensors, 17(7), 1483. https://doi.org/10.3390/s17071483