Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains and Culture Conditions
2.3. Biotinylation of Lactoferrin
2.4. Biacore Assay
2.5. Statistical Analysis
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kumar, J.; Weber, W.; Munchau, S.; Yadav, S.; Singh, S.B.; Saravanan, K.; Paramasivam, M.; Sharma, S.; Kaur, P.; Bhushan, A.; et al. Crystal structure of human seminal diferric lactoferrin at 3.4 angstrom resolution. Indian J. Biochem. Biophys. 2003, 40, 14–21. [Google Scholar] [PubMed]
- Severin, S.; Wenshui, X. Milk biologically active components as nutraceuticals: Review. Crit Rev. Food Sci. Nutr. 2005, 45, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Nishimura, T.; Yoshida, S. Presence of a glycan at a potential N-glycosylation site, Asn-281, of bovine lactoferrin. J. Dairy Sci. 2000, 83, 683–689. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M. Molecular structure, binding properties and dynamics of lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.; Colavizza, D.; Benaissa, M.; Maes, P.; Tartar, A.; Montreuil, J.; Spik, G. Molecular cloning and sequence analysis of bovine lactotransferrin. Eur. J. Biochem. 1991, 196, 177–184. [Google Scholar] [CrossRef] [PubMed]
- O'Riordan, N.; Kane, M.; Joshi, L.; Hickey, R.M. Structural and functional characteristics of bovine milk protein glycosylation. Glycobiology 2014, 24, 220–236. [Google Scholar] [CrossRef] [PubMed]
- O'Riordan, N.; Gerlach, J.Q.; Kilcoyne, M.; O’Callaghan, J.; Kane, M.; Hickey, R.M.; Joshi, L. Profiling temporal changes in bovine milk lactoferrin glycosylation using lectin microarrays. Food Chem. 2014, 165, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, S.S.; Schoemaker, R.J.W.; Timmer, C.J.; Kamerling, J.P.; Dijkhuizen, L. Use of Wisteria floribunda agglutinin affinity chromatography in the structural analysis of the bovine lactoferrin N-linked glycosylation. Biochim. Biophys. Acta 2012, 1820, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Chavez, S.A.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin: Structure, function and applications. Int. J. Antimicrob. Agents 2009, 33, 301.e1–301.e8. [Google Scholar] [CrossRef] [PubMed]
- Debbabi, H.; Dubarry, M.; Rautureau, M.; Tome, D. Bovine lactoferrin induces both mucosal and systemic immune response in mice. J. Dairy Res. 1998, 65, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Ohashi, M.; Tanaka, T.; Kumura, H.; Kim, G.Y.; Kwon, I.K.; Goh, J.S.; Shimazaki, K. Growth-promoting effects of lactoferrin on L. acidophilus and Bifidobacterium spp. Biometals 2004, 17, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, W.S.; Ito, T.; Kumura, H.; Shimazaki, K. Growth promotion and cell binding ability of bovine lactoferrin to Bifidobacterium longum. Anaerobe 2009, 15, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Cornish, J.; Callon, K.E.; Naot, D.; Palmano, K.P.; Banovic, T.; Bava, U.; Watson, M.; Lin, J.M.; Tong, P.C.; Chen, Q.; et al. Lactoferrin is a potent regulator of bone cell activity and increases bone formation in vivo. Endocrinology 2004, 145, 4366–4374. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Saito, D.; Akasu, T.; Alexander, D.B.; Futakuchi, M.; Fukamachi, K.; et al. Cancer prevention by bovine lactoferrin: From animal studies to human trial. Biometals 2010, 23, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Sekine, K.; Fujita, K.; Ligo, M. Cancer prevention by bovine lactoferrin and underlying mechanisms a review of experimental and clinical studies. Biochem. Cell Biol. 2002, 80, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Adlerova, L.; Bartoskova, A.; Faldyna, M. Lactoferrin: A review. Vet. Med. 2008, 53, 457–468. [Google Scholar]
- Garcia-Montoya, I.A.; Cendon, T.S.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta 2012, 1820, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ binding to bovine lactoferrin enhances protein stability and influences the release of bacterial lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Mazurier, J.; Colavizza, D.; Montreuil, J.; Spik, G. Properties of the iron-binding site of the N-terminal lobe of human and bovine lactotransferrins—Importance of the glycan moiety and of the noncovalent interactions between the N-terminal and C-terminal lobes in the stability of the iron-binding site. Biochem. J. 1990, 266, 575–581. [Google Scholar] [PubMed]
- Moore, S.A.; Anderson, B.F.; Groom, C.R.; Haridas, M.; Baker, E.N. Three-dimensional structure of diferric bovine lactoferrin at 2.8 Å resolution. J. Mol. Biol. 1997, 274, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, H.A.; Geerts, M.E.; van Berkel, P.H.; Nuijens, J.H. The role of N-linked glycosylation in the protection of human and bovine lactoferrin against tryptic proteolysis. Eur. J. Biochem. 2004, 271, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hirmo, S.; Willen, R.; Wadstrom, T. Inhibition of Helicobacter pylori infection by bovine milk glycoconjugates in a BALB/cA mouse model. J. Med. Microbiol. 2001, 50, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Teraguchi, S.; Shin, K.; Fukuwatari, Y.; Shimamura, S. Glycans of bovine lactoferrin function as receptors for the type 1 fimbrial lectin of Escherichia coli. Infect. Immun. 1996, 64, 1075–1077. [Google Scholar] [PubMed]
- Yoshida, S.; Wei, Z.; Shinmura, Y.; Fukunaga, N. Separation of lactoferrin-a and -b from bovine colostrum. J. Dairy Sci. 2000, 83, 2211–2215. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Isoda, H.; Shinmoto, H.; Tanimoto, M.; Dosako, S.; Idota, T.; Nakajima, I. Inhibition by k-casein glycomacropeptide and lactoferrin of influenza virus hemmaglutination. Biosci. Biotechnol. Biochem. 1993, 57, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Isoda, H.; Tanimoto, M.; Dosako, S.; Idota, T.; Ahiko, K. Inhibition by lactoferrin and kappa-casein glycomacropeptide of binding of cholera-toxin to its receptor. Biosci. Biotechnol. Biochem. 1992, 56, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Mueller, F. Biomolecular interaction analysis in drug discovery using surface plasmon resonance technology. Curr. Pharm. Des. 2006, 12, 3999–4021. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.A.; Mehra, R.K.; Carrington, S.D.; Hickey, R.M. Development of biosensor-based assays to identify anti-infective oligosaccharides. Anal. Biochem. 2011, 410, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-H.; Chen, R.-H.; Lee, C.-H.; Chang, Y.; Chen, C.-S.; Chen, W.-Y. Studies of the binding mechanism between aptamers and thrombin by circular dichroism, surface plasmon resonance and isothermal titration calorimetry. Colloids Surf. B 2011, 88, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Šípová, H.; Zhang, S.; Dudley, A.M.; Galas, D.; Wang, K.; Homola, J. Surface plasmon resonance biosensor for rapid label-free detection of microribonucleic acid at subfemtomole level. Anal. Chem. 2010, 82, 10110–10115. [Google Scholar] [CrossRef] [PubMed]
- Milkani, E.; Lambert, C.R.; McGimpsey, W.G. Direct detection of acetylcholinesterase inhibitor binding with an enzyme-based surface plasmon resonance sensor. Anal. Biochem. 2011, 408, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Schlick, K.H.; Cloninger, M.J. Inhibition binding studies of glycodendrimer/lectin interactions using surface plasmon resonance. Tetrahedron 2010, 66, 5305–5310. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.E.; Abbas, S.; Bennemo, M.; Larsson, A.; Hämäläinen, M.D.; Frostell-Karlsson, Å. A novel assay for influenza virus quantification using surface plasmon resonance. Vaccine 2010, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Altaye, M.; Jiang, X.; Guerrero, M.L.; Meinzen-Derr, J.K.; Farkas, T.; Chaturvedi, P.; Pickering, L.K.; Newburg, D.S. Human milk oligosaccharide blood group epitopes and innate immune protection against Campylobacter and calicivirus diarrhea in breastfed infants. In Protecting Infants through Human Milk; Pickering, L., Morrow, A., Ruiz-Palacios, G., Schanler, R., Eds.; Kluwer academic/Plenum publishers: New York, NY, USA, 2004; Volume 554, pp. 443–446. [Google Scholar]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.N.; Okawara, T.; Watanabe, M.; Kawai, Y.; Kitazawa, H.; Ohnuma, S.; Shibata, C.; Horii, A.; Kimura, K.; Taketomo, N.; et al. New screening methods for probiotics with adhesion properties to sialic acid and sulphate residues in human colonic mucin using the biacore assay. J. Appl. Microbiol. 2013, 114, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kitazawa, H.; Miura, K.; Shiiba, K.; Horii, A.; Saito, T. Quantitative evaluation of adhesion of lactobacilli isolated from human intestinal tissues to human colonic mucin using surface plasmon resonance (biacore assay). J. Appl. Microbiol. 2007, 102, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.D.; May, K.; Johansson, V.; Markey, F.; Critchley, I.A. Studies on the interaction of Staphylococcus aureus and Staphylococcus epidermidis with fibronectin using surface plasmon resonance (biacore). J. Microbiol. Methods 1997, 28, 77–84. [Google Scholar] [CrossRef]
- Oli, M.W.; McArthur, W.P.; Brady, L.J. A whole cell BIAcore assay to evaluate P1-mediated adherence of Streptococcus mutans to human salivary agglutinin and inhibition by specific antibodies. J. Microbiol. Methods 2006, 65, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Bulard, E.; Bouchet-Spinelli, A.; Chaud, P.; Roget, A.; Calemczuk, R.; Fort, S.; Livache, T. Carbohydrates as new probes for the identification of closely related Escherichia coli strains using surface plasmon resonance imaging. Anal. Chem. 2015, 87, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Naidu, A.S.; Andersson, M.; Miedzobrodzki, J.; Forsgren, A.; Watts, J.L. Bovine lactoferrin receptors in Staphylococcus aureus isolated from bovine mastitis. J. Dairy Sci. 1991, 74, 1218–1226. [Google Scholar] [CrossRef]
- Roe, A.J.; Currie, C.; Smith, D.G.; Gally, D.L. Analysis of type 1 fimbriae expression in verotoxigenic Escherichia coli: A comparison between serotypes O157 and O26. Microbiology 2001, 147, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.N.; Sun, D.X.; Dale, J.B.; Beachey, E.H. Conservation of the D-mannose-adhesion protein among type-1 fimbriated members of the family enterobacteriaceae. Nature 1988, 336, 682–684. [Google Scholar] [CrossRef] [PubMed]
- Jaques, L.W.; Brown, E.B.; Barrett, J.M.; Brey, W.S., Jr.; Weltner, W., Jr. Sialic acid. A calcium-binding carbohydrate. J. Biol. Chem. 1977, 252, 4533–4538. [Google Scholar] [PubMed]
- O’Flaherty, S.; Flynn, J.; Coffey, A.; Fitzgerald, G.; Meaney, B.; Ross, P. Molecular Characterisation of Bacteriophage K towards Applications for the Biocontrol of Pathogenic Staphylococci. Ph.D. Thesis, University College, Cork, Ireland, 2005. [Google Scholar]
- Hagiwara, S.; Kawai, K.; Anri, A.; Nagahata, H. Lactoferrin concentrations in milk from normal and subclinical mastitic cows. J. Vet. Med. Sci. 2003, 65, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Szunerits, S.; Zagorodko, O.; Cogez, V.; Dumych, T.; Chalopin, T.; Alvarez Dorta, D.; Sivignon, A.; Barnich, N.; Harduin-Lepers, A.; Larroulet, I.; et al. Differentiation of crohn’s disease-associated isolates from other pathogenic Escherichia coli by fimbrial adhesion under shear force. Biology 2016, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Erdei, J.; Forsgren, A.; Naidu, A.S. Lactoferrin binds to porins OmpF and OmpC in Escherichia coli. Infect. Immun. 1994, 62, 1236–1240. [Google Scholar] [PubMed]
- Naidu, S.S.; Erdei, J.; Czirok, E.; Kalfas, S.; Gado, I.; Thoren, A.; Forsgren, A.; Naidu, A.S. Specific binding of lactoferrin to Escherichia coli isolated from human intestinal infections. APMIS 1991, 99, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Yamauchi, K.; Takase, M. Inhibitory effects of bovine lactoferrin and lactoferricin b on Enterobacter sakazakii. Biocontrol Sci. 2008, 13, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Harouna, S.; Carramiñana, J.J.; Navarro, F.; Pérez, M.D.; Calvo, M.; Sánchez, L. Antibacterial activity of bovine milk lactoferrin on the emerging foodborne pathogen Cronobacter sakazakii: Effect of media and heat treatment. Food Control 2015, 47, 520–525. [Google Scholar] [CrossRef]
- Quintero-Villegas, M.; Wittke, A.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by lactoferrin. Curr. Microbiol. 2014, 69, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Quintero, M.; Maldonado, M.; Perez-Munoz, M.; Jimenez, R.; Fangman, T.; Rupnow, J.; Wittke, A.; Russell, M.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by prebiotic oligosaccharides. Curr. Microbiol. 2011, 62, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Halpin, R.; Brady, D.; O’Riordan, E.; O’Sullivan, M. Untreated and enzyme-modified bovine whey products reduce association of Salmonella typhimurium, Escherichia coli O157: H7 and Cronobacter malonaticus (formerly Enterobacter sakazakii) to Caco-2 cells. J. Appl. Microbiol. 2010, 108, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Bethe, G.; Remane, P.H.; Chhatwal, G.S. Identification of pneumococcal surface protein A as a lactoferrin-binding protein of Streptococcus pneumoniae. Infect. Immun. 1999, 67, 1683–1687. [Google Scholar] [PubMed]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kawase, K.; Tomita, M. Antibacterial spectrum of lactoferricin b, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Andersson, B.; Dahmen, J.; Frejd, T.; Leffler, H.; Magnusson, G.; Noori, G.; Eden, C.S. Identification of an active disaccharide unit of a glycoconjugate receptor for pneumococci attaching to human pharyngeal epithelial cells. J. Exp. Med. 1983, 158, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Lesman-Movshovich, E.; Lerrer, B.; Gilboa-Garber, N. Blocking of Pseudomonas aeruginosa lectins by human milk glycans. Can. J. Microbiol. 2003, 49, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Giannasca, K.T.; Giannasca, P.J.; Neutra, M.R. Adherence of Salmonella typhimurium to Caco-2 cells: Identification of a glycoconjugate receptor. Infect. Immun. 1996, 64, 135–145. [Google Scholar] [PubMed]
- Robertsson, J.A.; Lindberg, A.A.; Hoiseth, S.; Stocker, B.A. Salmonella typhimurium infection in calves: Protection and survival of virulent challenge bacteria after immunization with live or inactivated vaccines. Infect. Immun. 1983, 41, 742–750. [Google Scholar] [PubMed]



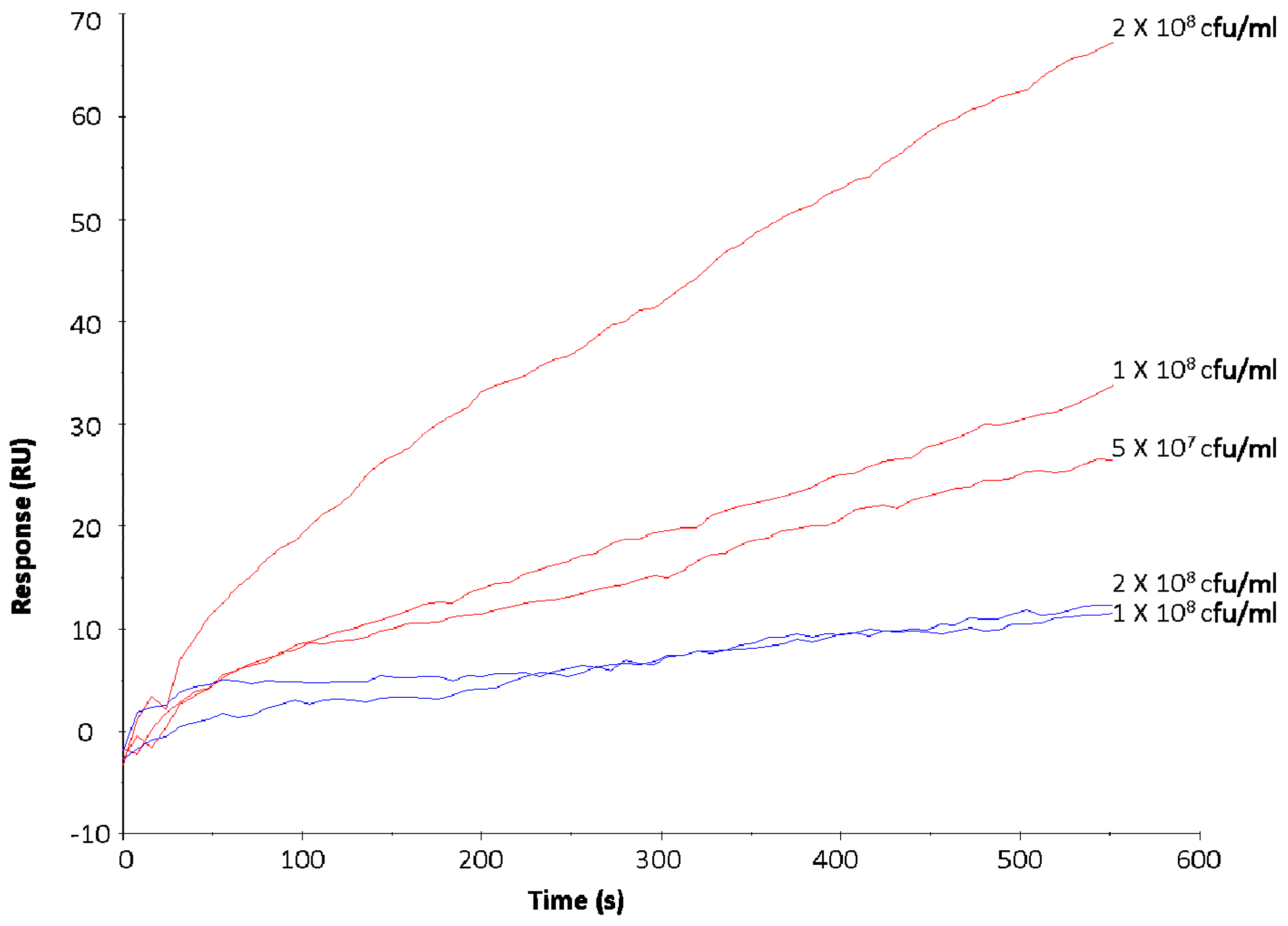{kind=link}
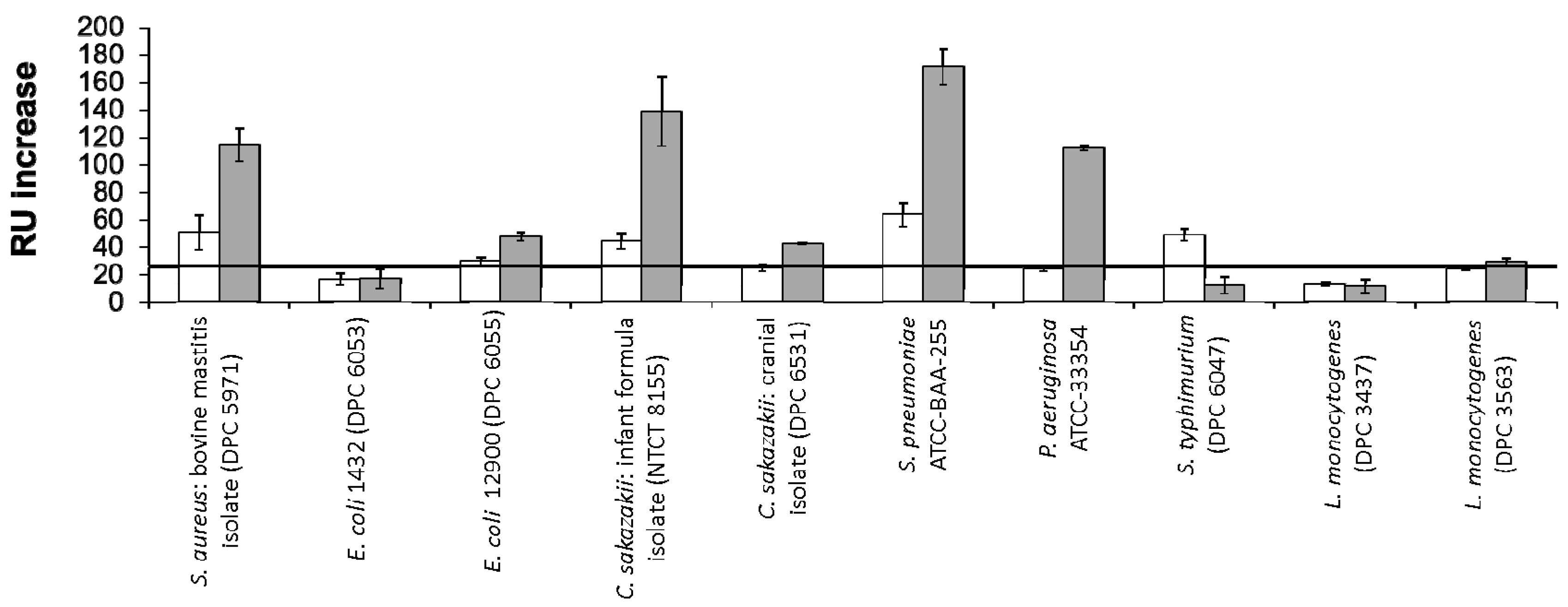{kind=link}
| Strain | Media |
|---|---|
| Staphylococcus aureus DPC 5971 | Muller-Hinton |
| Escherichia coli O157:H7 P1432 | Muller-Hinton |
| Escherichia coli O157:H7 NCTC 12900 | Muller-Hinton |
| Cronobacter sakazakii NCTC 8155 | Brain Heart Infusion |
| Cronobacter sakazakii DPC 6531 | Brain Heart Infusion |
| Streptococcus pneumoniae ATCC BAA-255 | Todd Hewitt + 0.5% yeast extract |
| Pseudomonas aeruginosa ATCC 33354 | Tryptic soy broth |
| Salmonella enterica subsp. enterica serovar Typhimurium ATCC BAA-185 | Brain Heart Infusion |
| Listeria monocytogenes DPC 3437 | Brain Heart Infusion |
| Listeria monocytogenes NCTC 11994 | Brain Heart Infusion |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Riordan, N.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors 2017, 17, 1515. https://doi.org/10.3390/s17071515
O’Riordan N, Kilcoyne M, Joshi L, Hickey RM. Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors. 2017; 17(7):1515. https://doi.org/10.3390/s17071515
Chicago/Turabian StyleO’Riordan, Noelle, Michelle Kilcoyne, Lokesh Joshi, and Rita M. Hickey. 2017. "Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria" Sensors 17, no. 7: 1515. https://doi.org/10.3390/s17071515
APA StyleO’Riordan, N., Kilcoyne, M., Joshi, L., & Hickey, R. M. (2017). Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors, 17(7), 1515. https://doi.org/10.3390/s17071515