
Deciphering the Disaggregation Mechanism of Amyloid Beta Aggregate by 4-(2-Hydroxyethyl)-1-Piperazinepropanesulfonic Acid Using Electrochemical Impedance Spectroscopy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
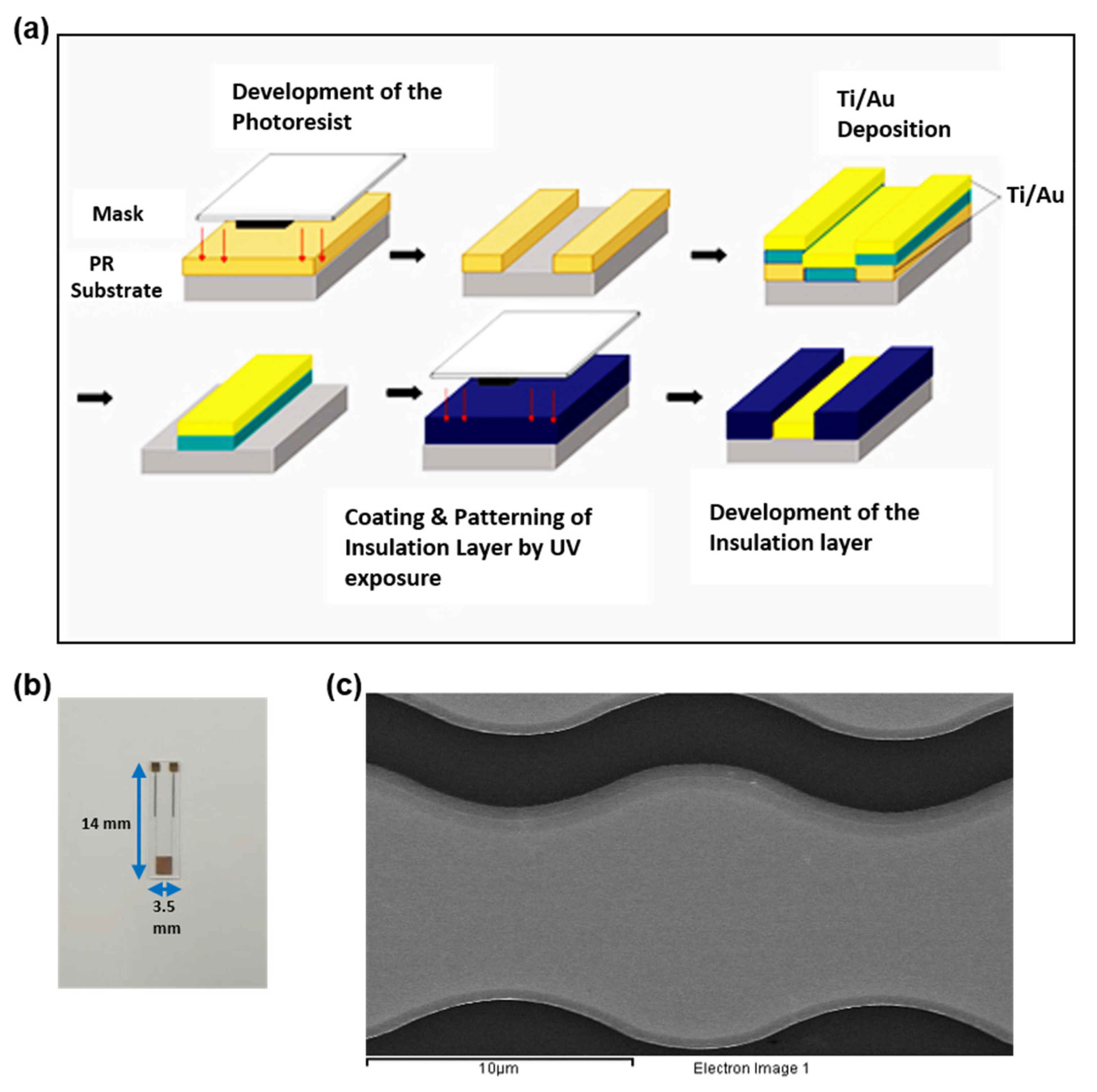
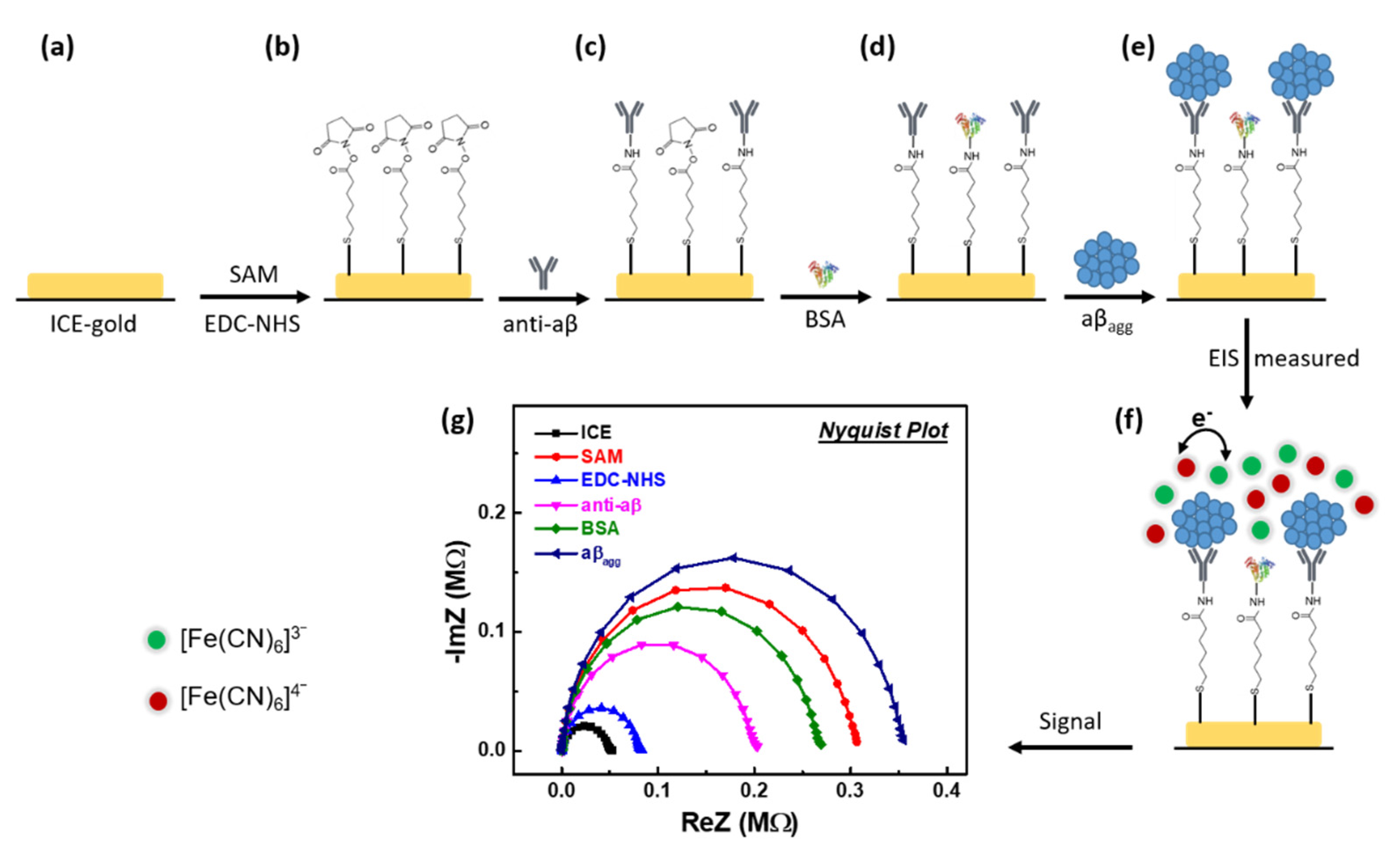
2.2. Development of the Immunosensor for Monitoring the Impact of EPPS on the Disaggregation of aβagg
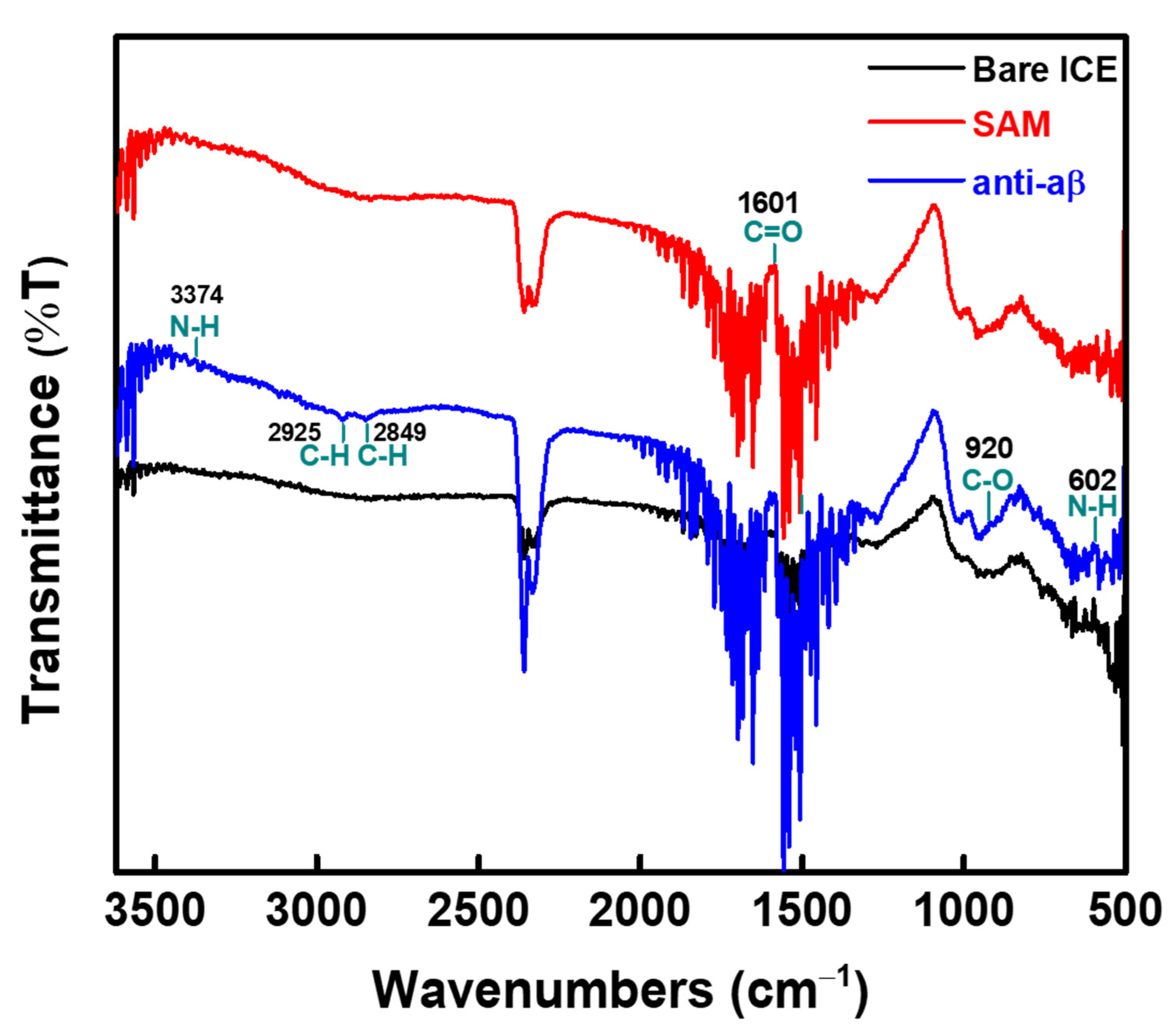
2.3. EIS Measurements and Functional Group Characterization
3. Results and Discussion
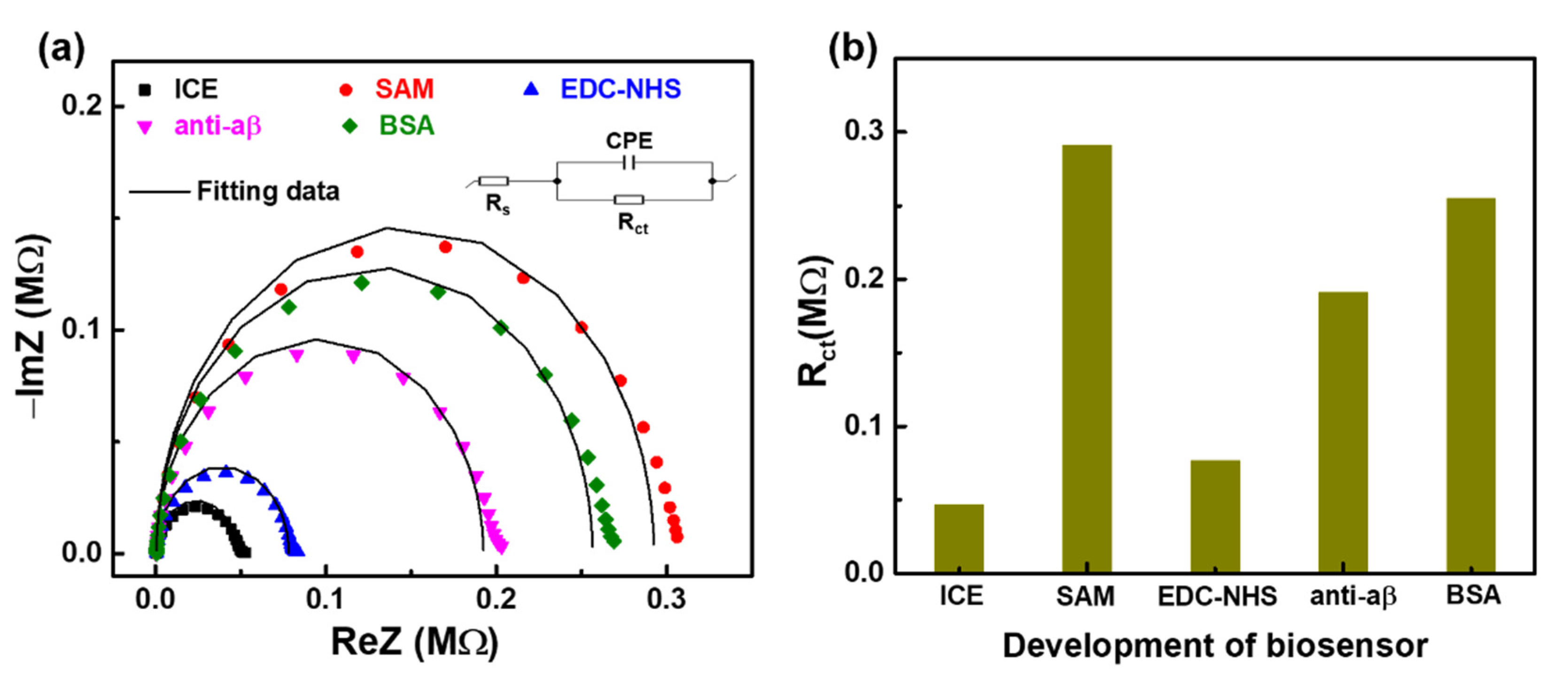
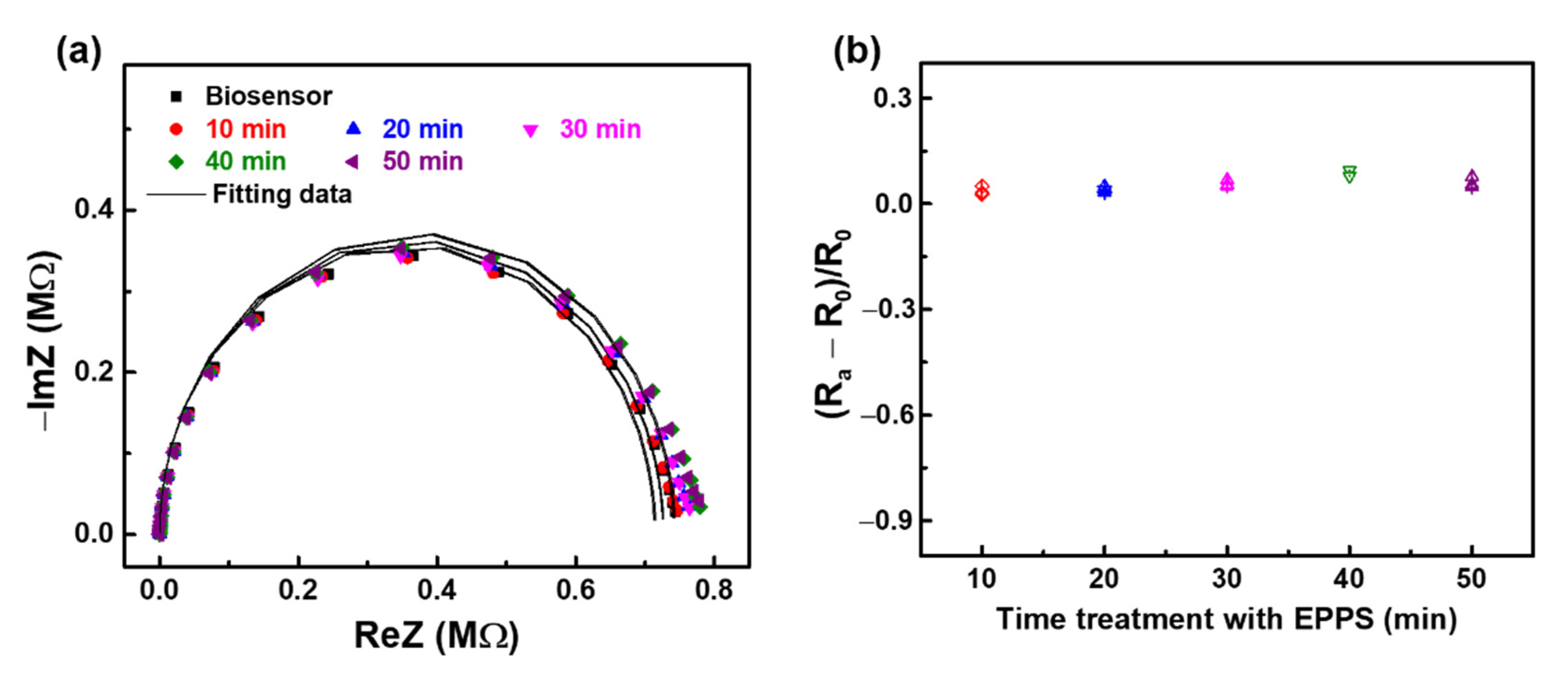
3.1. Investigating the Development of Impedimetric Immunosensor
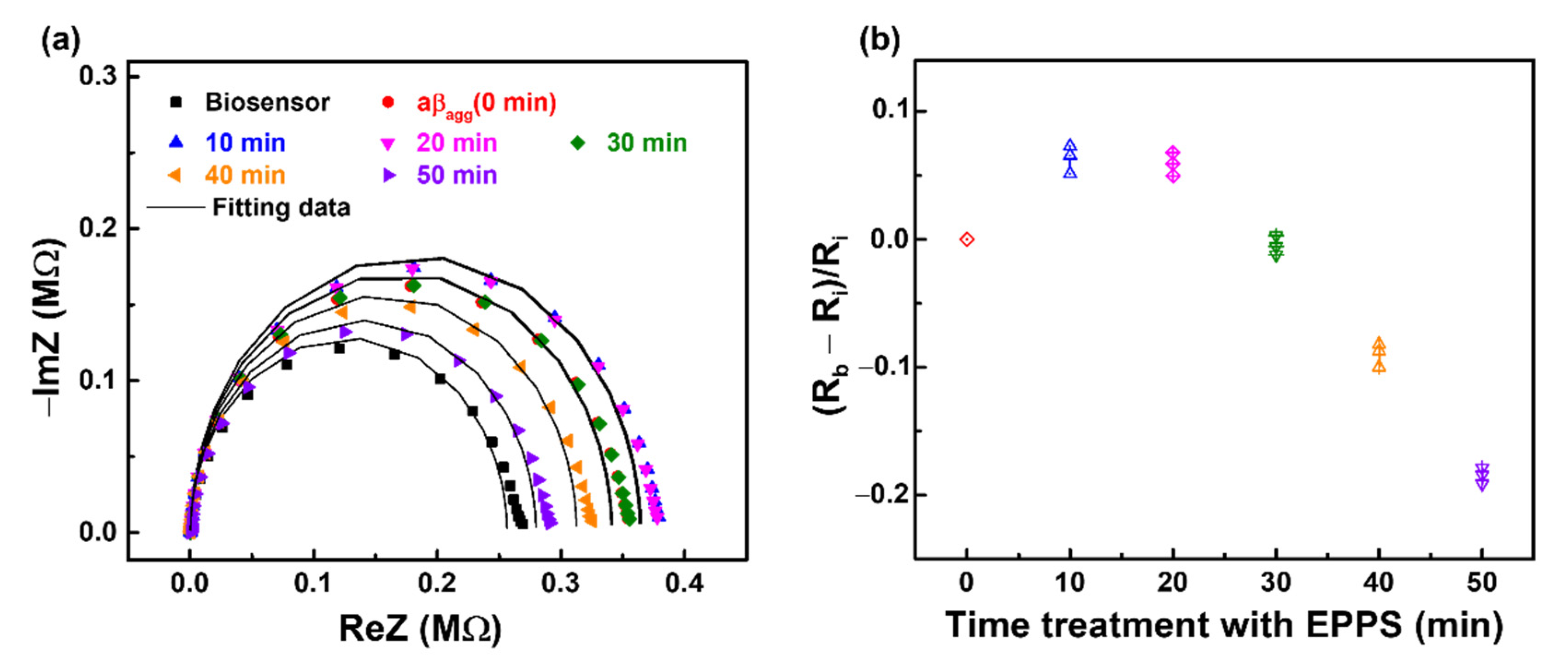
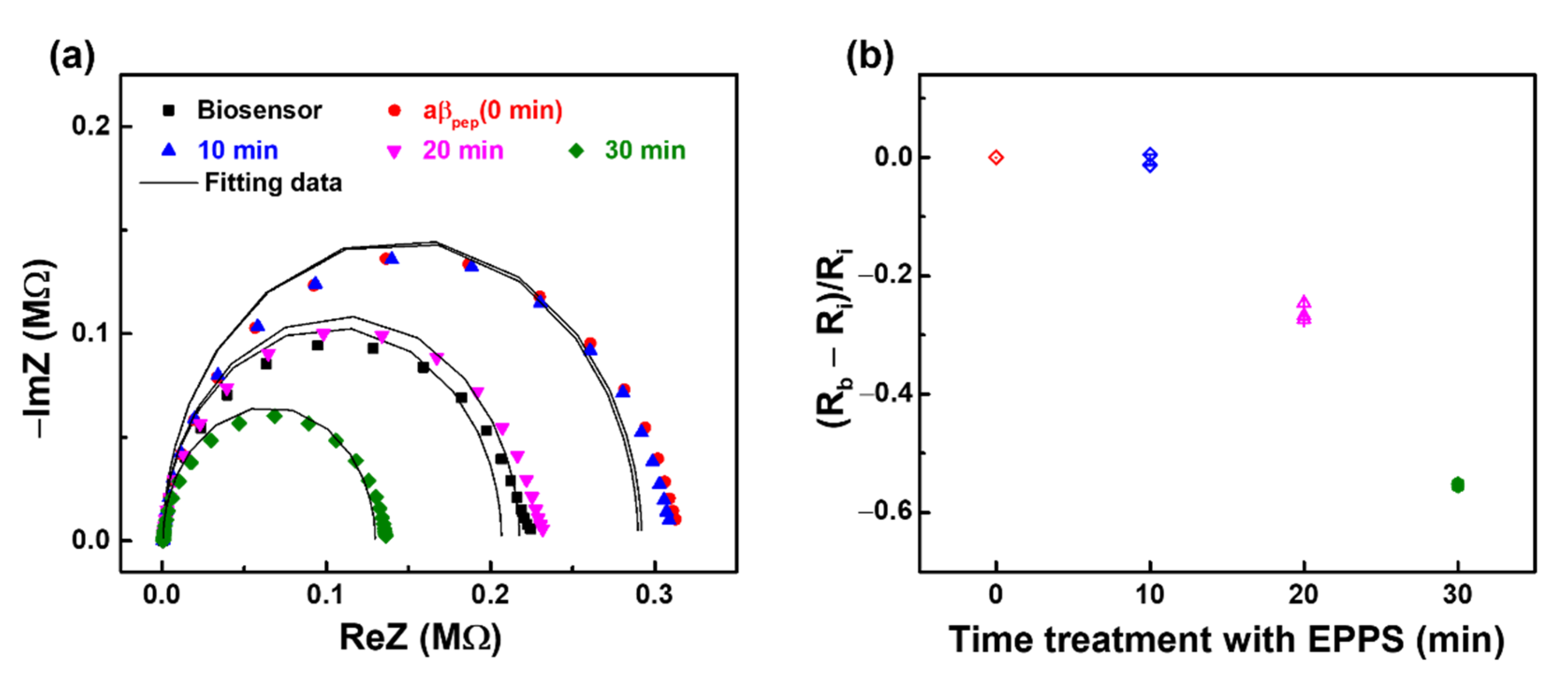
3.2. Monitoring the Effect of EPPS on aβ Disaggregation
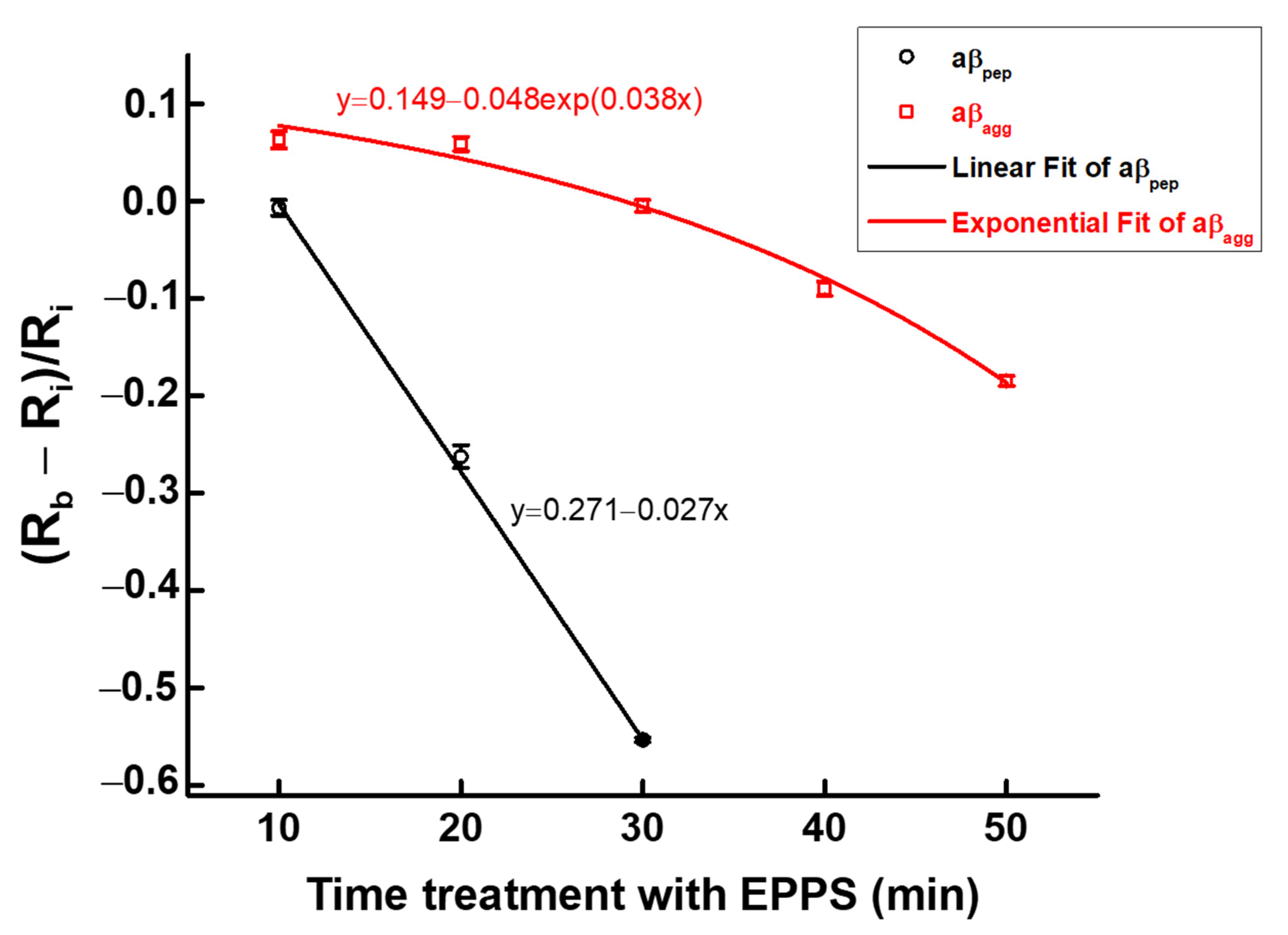
3.3. Kinetic Disaggregation Rate k
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murphy, M.P.; LeVine, H., III. Alzheimer’s Disease and the β-Amyloid Peptide. J. Alzheimer’s Dis. 2010, 19, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blennow, K.; Mattsson, N.; Schöll, M.; Hansson, O.; Zetterberg, H. Amyloid biomarkers in Alzheimer’s disease. Trends Pharmacol. Sci. 2015, 36, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Clifford, R.J., Jr.; Bennett, D.A.; Blennow, K.; Carrillod, M.C.; Dunne, B.; Haeberleinf, S.B.; Holtzmang, D.M.; Jagusth, W.; Jesseni, F.; Karlawishj, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar]
- O’Brien, R.J.; Wong, P.C. Amyloid Precursor Protein Processing and Alzheimer’s Disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, H.; Ozawa, D.; Sakurai, K.; Kawakami, T.; Kuyama, H.; Nishimura, O.; Shimanouchi, T.; Kuboi, R.; Naiki, H.; Goto, Y. Laser-induced Propagation and Destruction of Amyloid β Fibrils. J. Biol. Chem. 2010, 285, 19660–19667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.S.; Jung, H.H.; Son, M.G.; Lee, H.B.; Kwak, T.J.; Lee, G.D.; Kim, C.H.S.; Lee, W.; Yoon, D.S. Characterization of the regrowth behavior of amyloid-like fragmented fibrils decomposed by ultrasonic treatment. RSC Adv. 2014, 4, 56561–56566. [Google Scholar] [CrossRef]
- Takeda, T.; Klimov, D.K. Temperature-Induced Dissociation of Aβ Monomers from Amyloid Fibril. Biophys. J. 2008, 95, 1758–1772. [Google Scholar] [CrossRef] [Green Version]
- Bayliss, D.L.; Walsh, J.L.; Shama, G.; Iza, F.; Kong, M.G. Reduction and degradation of amyloid aggregates by a pulsed radio-frequency cold atmospheric plasma jet. New J. Phys. 2009, 11, 115024. [Google Scholar] [CrossRef] [Green Version]
- Sacchettini, J.C.; Kelly, J.W. Therapeutic strategies for human amyloid diseases. Nat. Rev. Drug Discov. 2002, 1, 267–275. [Google Scholar] [CrossRef]
- Karran, E.; Mercken, M.; Strooper, B.D. The amyloid cascade hypothesis for Alzheimer’s disease: An appraisalfor the development of therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef]
- Gerlinde, G.; Albrecht, O.; Mario, L.; Ronald, F.F.; Erich, E.W.; Jan, B. Black tea theaflavins inhibit formation of toxic amyloid-β and α-synuclein fibrils. Biochemistry 2011, 50, 10624–10636. [Google Scholar]
- Lee, J.W.; Lee, Y.K.; Ban, J.O.; Ha, T.Y.; Yun, Y.P.; Han, S.B.; Oh, K.W.; Hong, J.T. Green tea (-)-epigallocatechin-3-gallate inhibits beta-amyloid-induced cognitive dysfunction through modification of secretase activity via inhibition of ERK and NF-kappaB pathways in mice. J. Nutr. 2009, 139, 1987–1993. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Liu, X.; Gong, H.; Huang, L.; Chen, H.; Zhang, X.; Li, C.; Yang, M.; Ma, B.; Jiao, L.; et al. Coffee Components Inhibit Amyloid Formation of Human Islet Amyloid Polypeptide in Vitro: Possible Link between Coffee Consumption and Diabetes Mellitus. J. Agric. Food Chem. 2011, 59, 13147–13155. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, H.V.; Jo, S.M.; Lee, C.J.; Choi, S.Y.; Kim, D.J.; Kim, Y.S. EPPS rescues hippocampus-dependent cognitive deficits in APP/PS1 mice by disaggregation of amyloid-β oligomers and plaques. Nat. Commun. 2015, 6, 8997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, Y.K.; Kim, J.S.; Kim, G.; Kim, Y.S.; Kim, H.Y.; Lee, S.J.; Cho, W.W.; Kim, S.S.; Lee, S.M.; Lee, B.C.; et al. A highly sensitive plasma-based amyloid-β detection system through medium-changing and noise cancellation system for early diagnosis of the Alzheimer’s disease. Sci. Rep. 2017, 7, 8882. [Google Scholar] [CrossRef] [Green Version]
- Rushworth, J.V.; Ahmed, A.; Griffiths, H.H.; Pollock, N.M.; Hooper, N.M.; Millner, P.A. A label-free electrical impedimetric biosensor for the specific detection of Alzheimer’s amyloid-beta oligomers. Biosens. Bioelectron. 2014, 56, 83–90. [Google Scholar] [CrossRef]
- Xing, Y.; Feng, X.Z.; Zhang, L.; Hou, J.; Han, G.C.; Chen, Z. A sensitive and selective electrochemical biosensor for the determination of beta-amyloid oligomer by inhibiting the peptide-triggered in situ assembly of silver nanoparticles. Int. J. Nanomed. 2017, 12, 3171–3179. [Google Scholar] [CrossRef] [Green Version]
- Truong, T.N.L.; Takamura, Y.; Tamiya, E.; Vestergaard, M.C. Modified screen printed electrode for development of a highly sensitive label-free impedimetric immunosensor to detect amyloid beta peptides. Anal. Chim. Acta 2015, 892, 69–76. [Google Scholar]
- Chinnadayyala, S.R.; Cho, S.B. Electrochemical Immunosensor for the Early Detection of Rheumatoid Arthritis Biomarker: Anti-Cyclic Citrullinated Peptide Antibody in Human Serum Based on Avidin-Biotin System. Sensors 2021, 21, 124. [Google Scholar] [CrossRef]
- Martic, S.; Rains, M.K.; Kraatz, H.B. Probing copper/tau protein interactions electrochemically. Anal. Biochem. 2013, 442, 130–137. [Google Scholar] [CrossRef]
- Smith, B.C. Infrared Spectral Interpretation: A Systematic Approach; CRC Press Inc.: Boca Raton, FL, USA, 1998. [Google Scholar]
- Phan, L.M.T.; Kim, E.B.; Cheon, S.A.; Shim, T.S.; Kim, H.J.; Park, T.J. Reliable naked-eye detection of Mycobacterium tuberculosis antigen 85B using gold and copper nanoshell-enhanced immunoblotting techniques. Sens. Actuators B Chem. 2020, 317, 1282202. [Google Scholar] [CrossRef]
- Gaddam, S.A.; Kotakadi, V.S.; Gopal, D.V.R.S.; Rao, Y.S.; Reddy, A.V. Efficent and robust biofabrication of silver nanoparticles by Cassia alata leaf extract and their antimicrobial activity. J. Nanostruct. Chem. 2014, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Phan, L.M.T.; Rafique, R.; Baek, S.H.; Nguyen, T.P.; Park, K.Y.; Kim, E.B.; Kim, J.G.; Park, J.P.; Kailasa, S.K.; Kim, H.J.; et al. Gold-copper nanoshell dot-blot immunoassay for naked-eye sensitive detection of tuberculosis specific CFP-10 antigen. Biosens. Bioelectron. 2018, 121, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.H.V.; Gupta, Y.K. Effect of different extracts of Centella asiatica on cognition and markers of oxidative stress in rats. J. Ethnopharmacol. 2002, 79, 253–260. [Google Scholar] [CrossRef]
- Smith, B.J. SDS Polyacrylamide Gel Electrophoresis of Proteins. Proteins Methods Mol. Biol. 1984, 1, 41–56. [Google Scholar] [PubMed]
- An, S.S.A.; Lee, B.S.; Yu, J.S.; Lim, K.T.; Kim, G.J.; Lee, R.; Kim, S.W.; Kang, S.M.; Park, Y.H.; Wang, M.J.; et al. Dynamic changes of oligomeric amyloid β levels in plasma induced by spiked synthetic Aβ42. Alzheimer’s Res. Ther. 2017, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Le, H.T.N.; Park, J.S.; Chinnadayyala, S.R.; Cho, S.B. Sensitive electrochemical detection of amyloid beta peptide in human serum using an interdigitated chain-shaped electrode. Biosens. Bioelectron. 2019, 144, 111694. [Google Scholar]
- Lermyte, F.; Everett, J.; Yuko, P.Y.L.; Wootton, C.A.; Brooks, J.; Barrow, M.P.; Telling, N.D.; Sadler, P.J.; O’Connor, P.B.; Collingwood, J.F. Metal Ion Binding to the Amyloid β Monomer Studied by Native Top-Down FTICR Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2019, 30, 2123–2134. [Google Scholar] [CrossRef]
- Nair, G.N.; Perry, G.; Smith, M.A.; Reddy, V.P. NMR Studies of Zinc, Copper, and Iron Binding to Histidine, the Principal Metal Ion Complexing Site of Amyloid-β Peptide. J. Alzheimer’s Dis. 2010, 20, 57–66. [Google Scholar] [CrossRef]
- Cellmer, T.; Douma, R.; Huebner, A.; Prausnitz, J.; Blanch, H. Kinetic studies of protein L aggregation and disaggregation. Biophys. Chem. 2007, 125, 350–359. [Google Scholar] [CrossRef]
- Priezzhev, A.V.; Ryaboshapka, O.M.; Firsov, N.N.; Sirko, I.V. Aggregation and Disaggregation of Erythrocytes in Whole Blood: Study by Backscattering Technique. J. Biomed. Opt. 1999, 4, 76–84. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrode | Rs (Ω) | C (F) | Rct (Ω) |
|---|---|---|---|
| ICE | 529.3 | 55.37 × 10−9 | 47,294 |
| SAM | 522.8 | 77.78 × 10−9 | 291,956 |
| EDC-NHS | 524.9 | 69.69 × 10−9 | 77,691 |
| anti-aβ | 524 | 75.88 × 10−9 | 191,820 |
| BSA | 504.5 | 76.49 × 10−9 | 255,926 |
| Electrode | Rs (Ω) | C (F) | Rct (Ω) |
|---|---|---|---|
| Biosensor | 490.9 | 55.55 × 10−9 | 715,084 |
| 10 min | 491.9 | 56.72 × 10−9 | 724,551 |
| 20 min | 487.5 | 57.66 × 10−9 | 726,538 |
| 30 min | 488.7 | 57.93 × 10−9 | 724,904 |
| 40 min | 488.5 | 58.36 × 10−9 | 743,262 |
| 50 min | 486.3 | 58.83 × 10−9 | 730,896 |
| Electrode | Rs (Ω) | C (F) | Rct (Ω) |
|---|---|---|---|
| Biosensor | 504.8 | 76.10 × 10−9 | 228,331 |
| aβagg (0 min) | 519.2 | 76.29 × 10−9 | 331,631 |
| 10 min | 517.4 | 75.98 × 10−9 | 366,841 |
| 20 min | 517.1 | 75.89 × 10−9 | 363,145 |
| 30 min | 519.1 | 75.08 × 10−9 | 334,918 |
| 40 min | 519.5 | 74.81 × 10−9 | 307,364 |
| 50 min | 517.6 | 74.71 × 10−9 | 274,678 |
| Electrode | Rs (Ω) | C (F) | Rct (Ω) |
|---|---|---|---|
| Biosensor | 633.5 | 91.3 × 10−9 | 205,978 |
| aβpep (0 min) | 635 | 92.96 × 10−9 | 289,018 |
| 10 min | 632.6 | 92.17 × 10−9 | 289,110 |
| 20 min | 629.8 | 90.69 × 10−9 | 209,893 |
| 30 min | 639.9 | 88.06 × 10−9 | 129,281 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, H.T.N.; Cho, S. Deciphering the Disaggregation Mechanism of Amyloid Beta Aggregate by 4-(2-Hydroxyethyl)-1-Piperazinepropanesulfonic Acid Using Electrochemical Impedance Spectroscopy. Sensors 2021, 21, 788. https://doi.org/10.3390/s21030788
Le HTN, Cho S. Deciphering the Disaggregation Mechanism of Amyloid Beta Aggregate by 4-(2-Hydroxyethyl)-1-Piperazinepropanesulfonic Acid Using Electrochemical Impedance Spectroscopy. Sensors. 2021; 21(3):788. https://doi.org/10.3390/s21030788
Chicago/Turabian StyleLe, Hien T. Ngoc, and Sungbo Cho. 2021. "Deciphering the Disaggregation Mechanism of Amyloid Beta Aggregate by 4-(2-Hydroxyethyl)-1-Piperazinepropanesulfonic Acid Using Electrochemical Impedance Spectroscopy" Sensors 21, no. 3: 788. https://doi.org/10.3390/s21030788
APA StyleLe, H. T. N., & Cho, S. (2021). Deciphering the Disaggregation Mechanism of Amyloid Beta Aggregate by 4-(2-Hydroxyethyl)-1-Piperazinepropanesulfonic Acid Using Electrochemical Impedance Spectroscopy. Sensors, 21(3), 788. https://doi.org/10.3390/s21030788