
Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Herbicide Application
2.2. Reflectance/Fluorescence and ΦPSII
2.3. Chlorophyll Fluorescence Imaging
2.4. Image and Data Analysis
2.5. Pixel Intensity and Heat Dissipation
3. Results
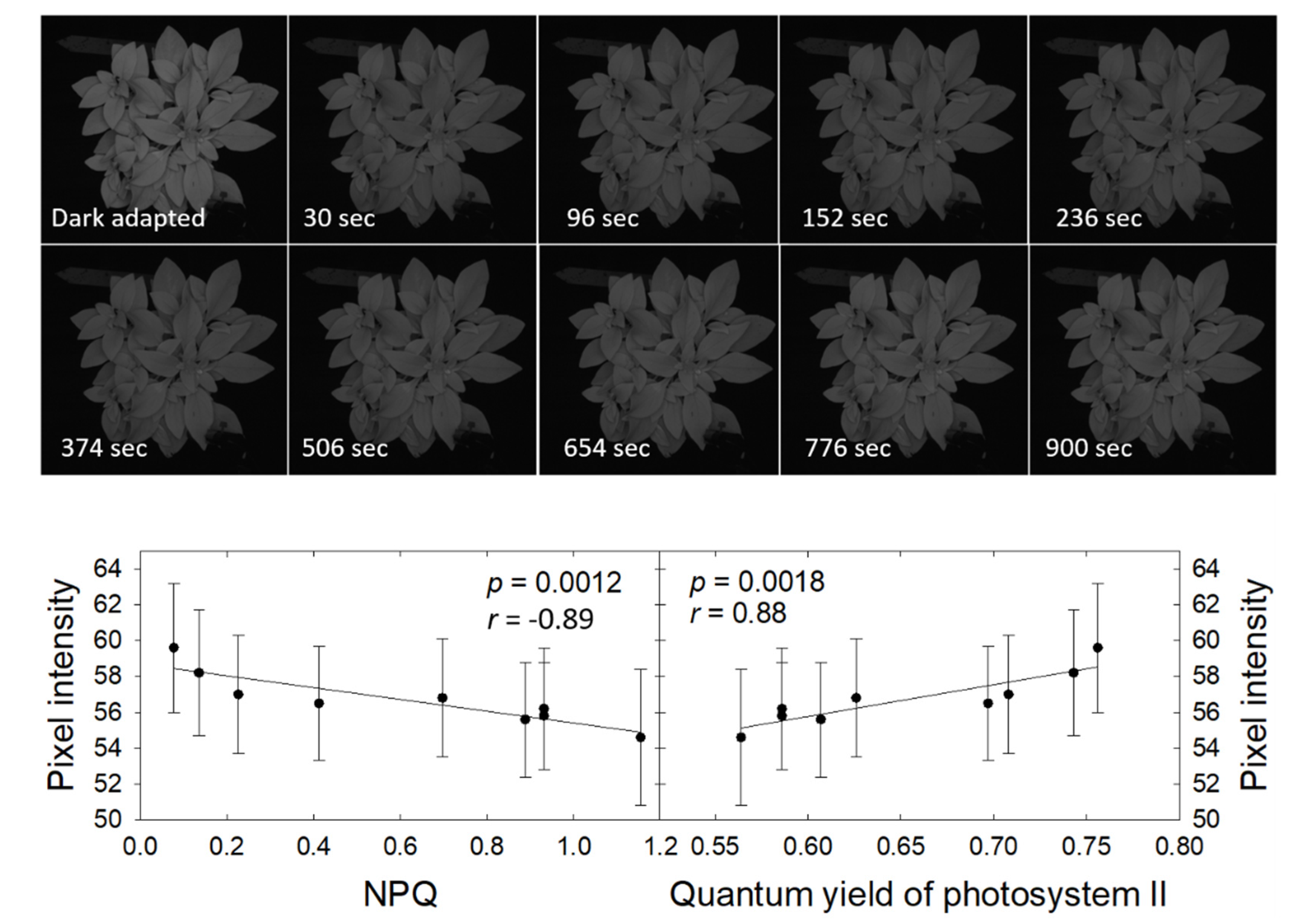
3.1. Chlorophyll Fluorescence Imaging
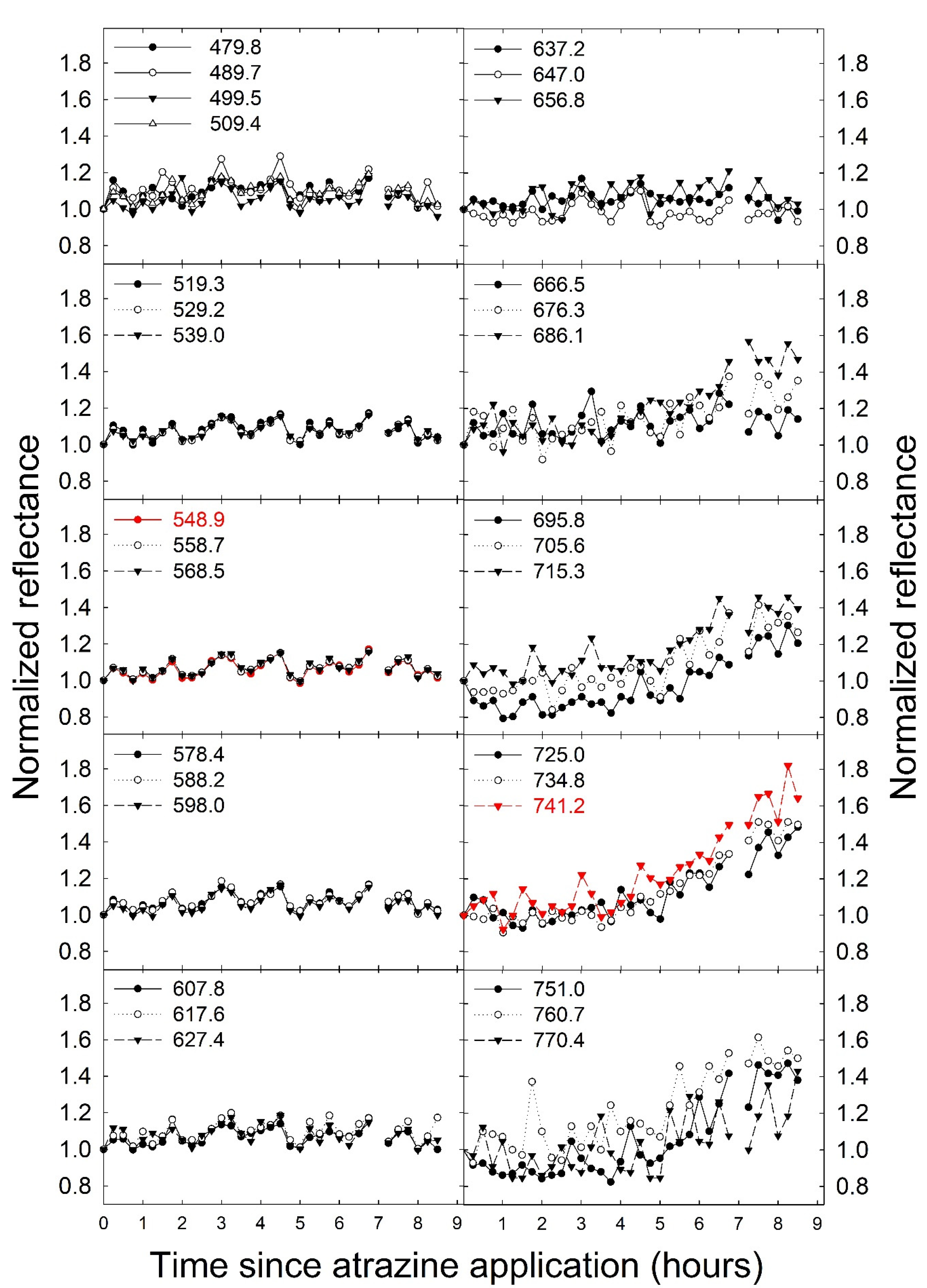
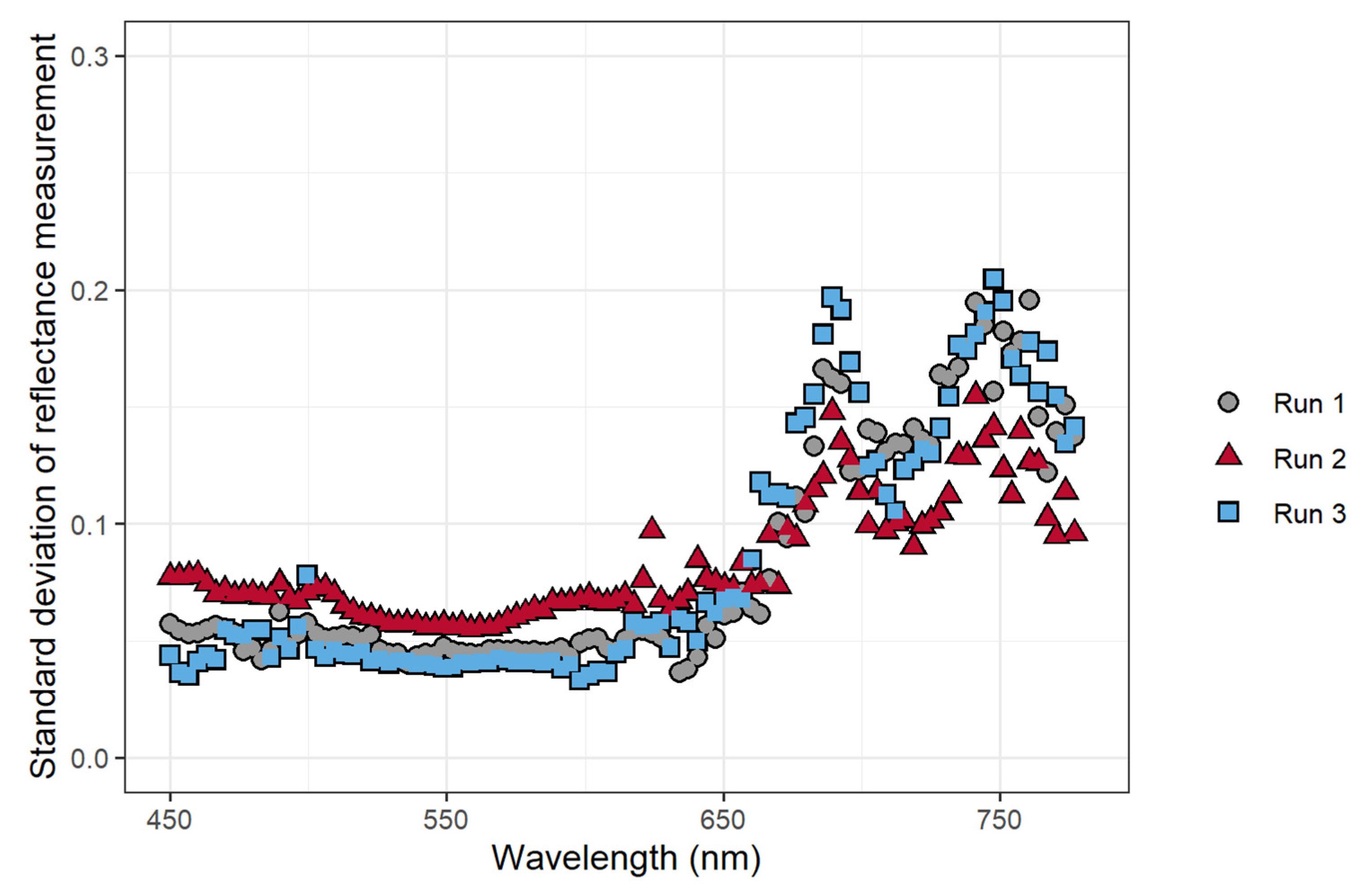
3.2. Examination of Fluorescence from Reflectance Measurements
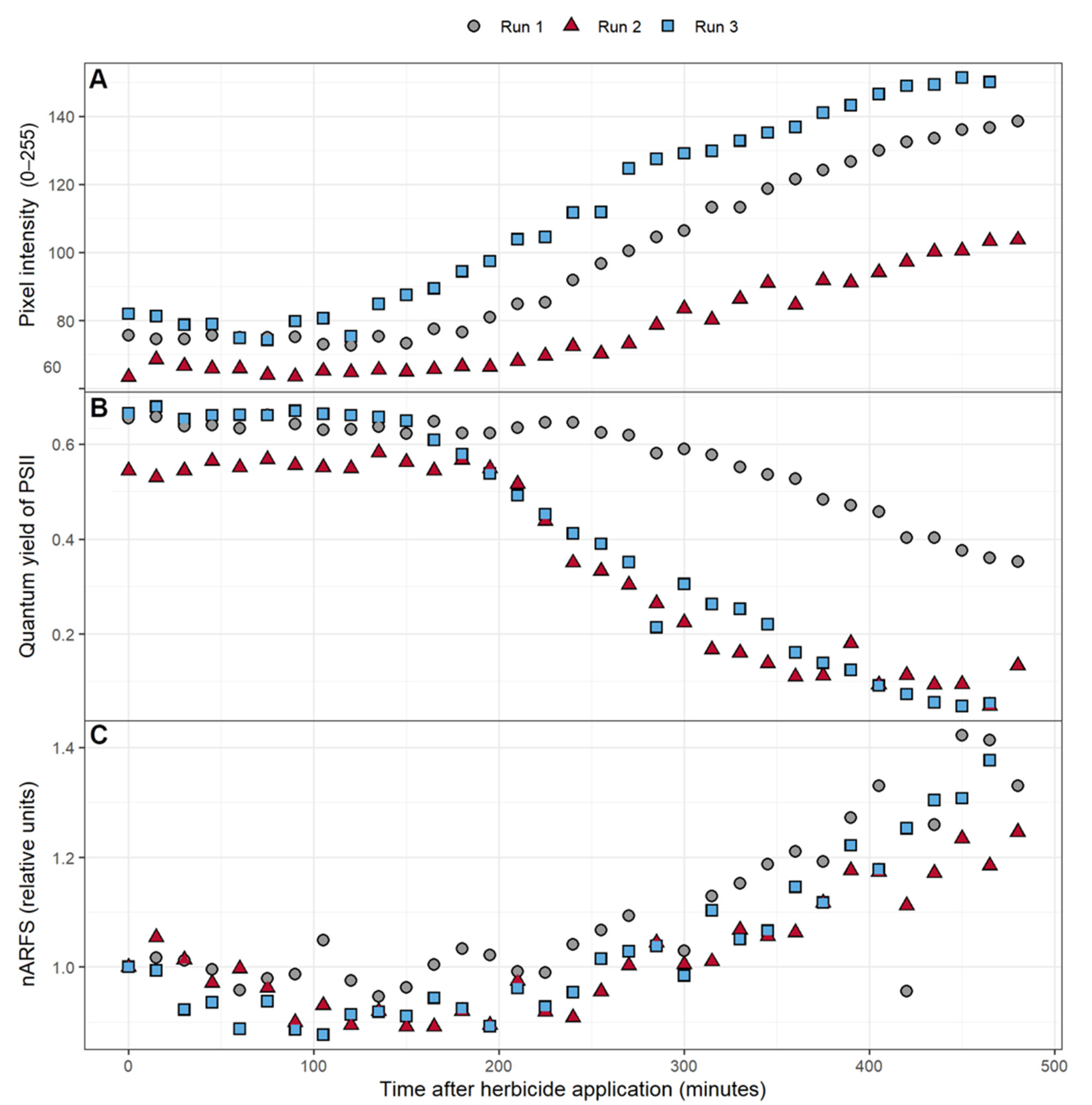
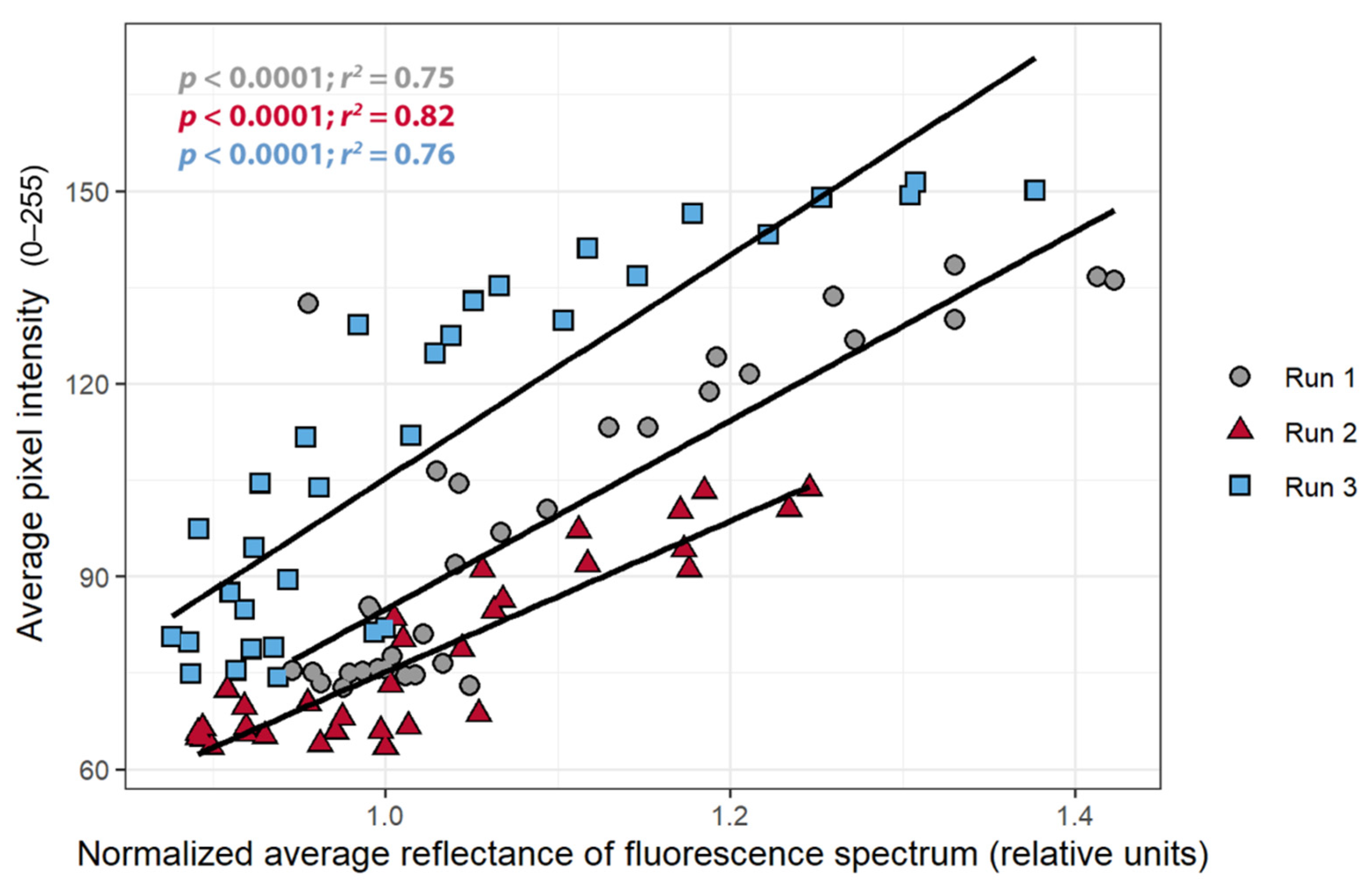
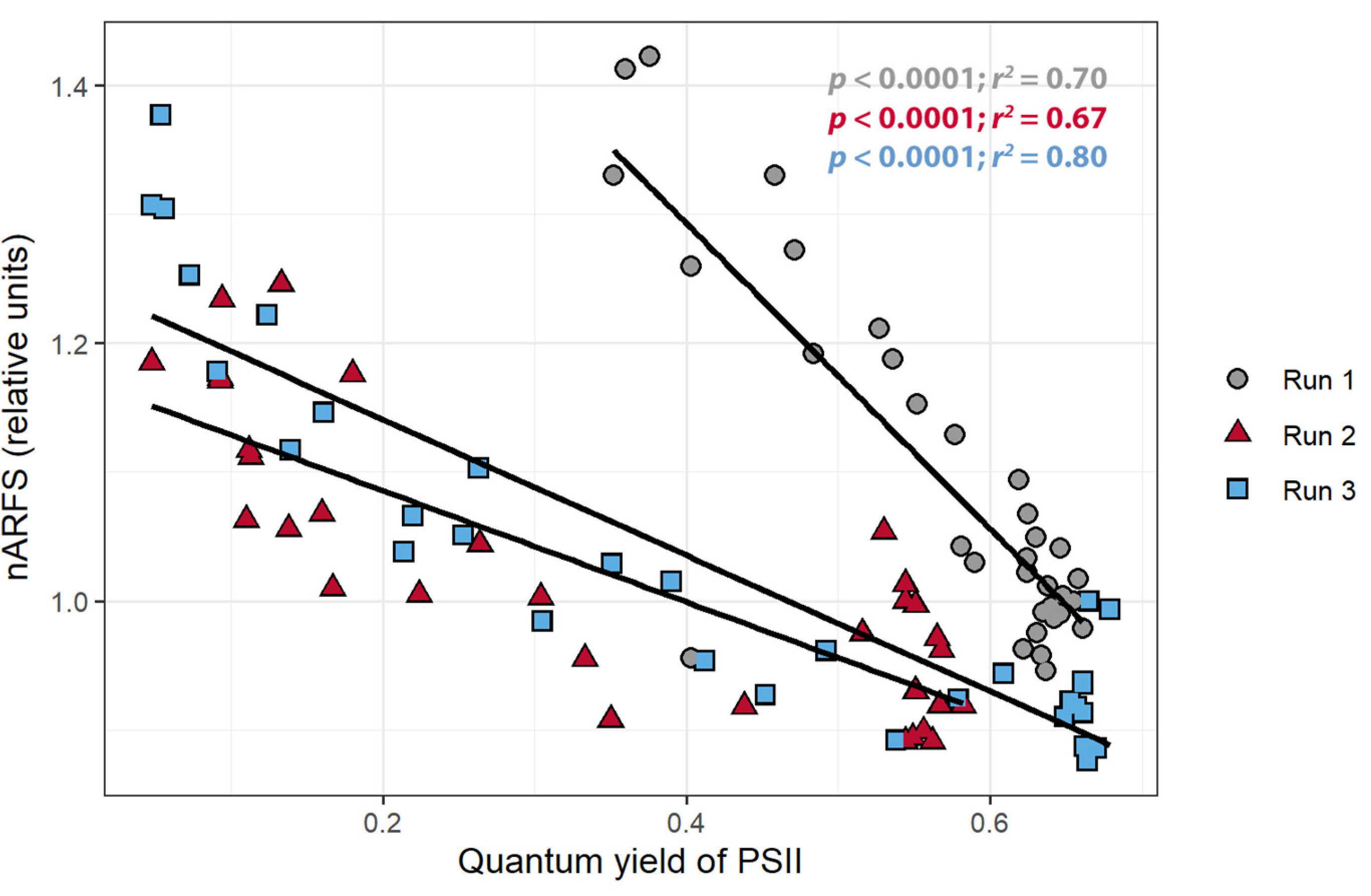
3.3. Pixel Intensity, ΦPSII, and nARFS
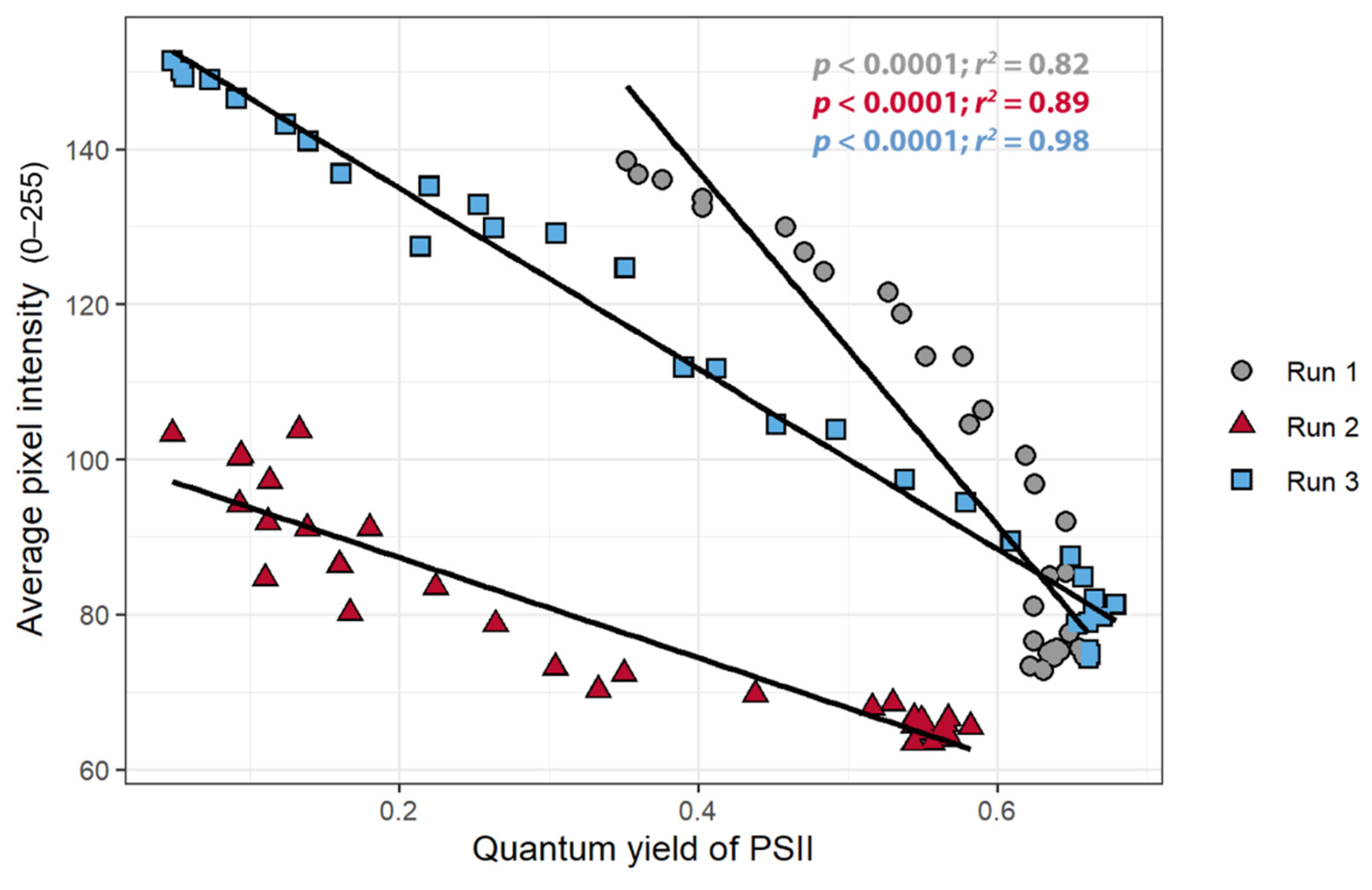
3.4. Relationships between Average Pixel Intensity, ΦPSII, and nARFS
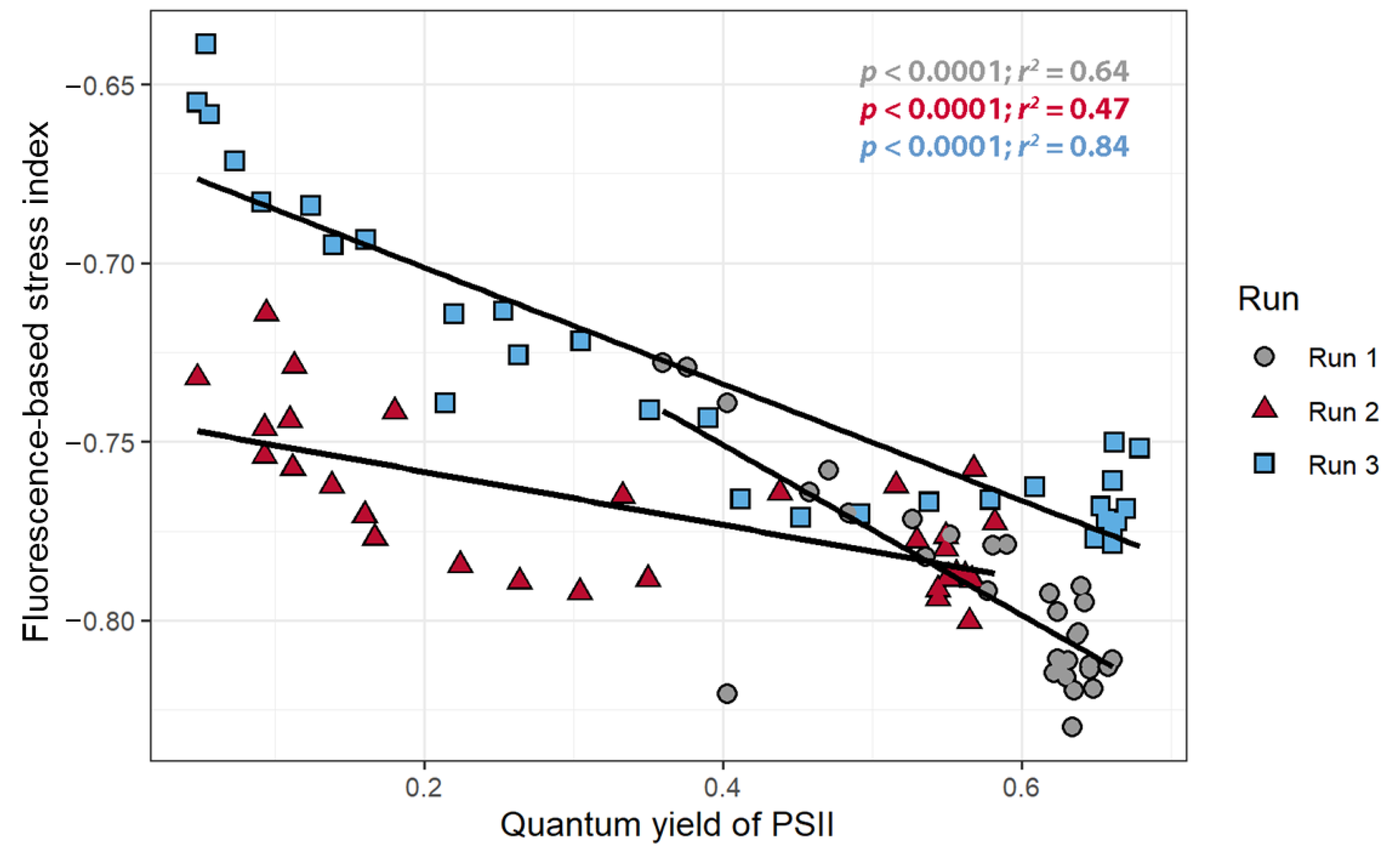
3.5. Development and Support for a Fluorescence-Based Stress Index
3.6. Pixel Intensity, Heat Dissipation, and Quantum Yield Recovery in the Dark
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kautsky, H.; Hirsch, A. Neue versuche zur kohlensäureassimilation. Naturwissenschaften 1931, 19, 964. [Google Scholar] [CrossRef]
- DeEll, J.R.; Toivonen, P.M. Practical Applications of Chlorophyll Fluorescence in Plant Biology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll Fluorescence as a Nonintrusive Indicator for Rapid Assessment of In Vivo Photosynthesis. In Ecophysiology of Photosynthesis; Springer: Berlin/Heidelberg, Germany, 1995; pp. 49–70. [Google Scholar]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence and Photosynthesis: The Basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Leaf chlorophyll fluorescence, reflectance, and physiological response to freshwater and saltwater flooding in the evergreen shrub, Myrica cerifera. Environ. Exp. Bot. 2008, 63, 402–409. [Google Scholar] [CrossRef]
- Chaerle, L.; Hagenbeek, D.; De Bruyne, E.; Valcke, R.; Van Der Straeten, D. Thermal and Chlorophyll-Fluorescence Imaging Distinguish Plant-Pathogen Interactions at an Early Stage. Plant Cell Physiol. 2004, 45, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaerle, L.; Leinonen, I.; Jones, H.G.; Van Der Straeten, D. Monitoring and screening plant populations with combined thermal and chlorophyll fluorescence imaging. J. Exp. Bot. 2006, 58, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Bilger, W.; Schreiber, U.; Lange, O. Chlorophyll fluorescence as an indicator of heat induced limitation of photosyn-thesis in Arbutus unedo L. In Plant Response to Stress; Springer: Berlin/Heidelberg, Germany, 1987; pp. 391–399. [Google Scholar]
- Daley, P.F. Chlorophyll fluorescence analysis and imaging in plant stress and disease. Can. J. Plant Pathol. 1995, 17, 167–173. [Google Scholar] [CrossRef]
- Gibbons, G.C.; Smillie, R.M. Chlorophyll fluorescence photography to detect mutants, chilling injury and heat stress. Carlsberg Res. Commun. 1980, 45, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Rinderle, U. The Role of Chlorophyll Fluorescence in The Detection of Stress Conditions in Plants. C R C Crit. Rev. Anal. Chem. 1988, 19, S29–S85. [Google Scholar] [CrossRef]
- Strand, M.; Öquist, G. Inhibition of photosynthesis by freezing temperatures and high light Levels in cold-acclimated seedlings of Scots pine (Pinus sylvestris).-I. Effects on the light-limited and light-saturated rates of CO2 assimilation. Physiol. Plant. 1985, 64, 425–430. [Google Scholar] [CrossRef]
- Huner, N.P.; Öquist, G.; Hurry, V.M.; Krol, M.; Falk, S.; Griffith, M. Photosynthesis, photoinhibition and low temper-ature acclimation in cold tolerant plants. Photosynth. Res. 1993, 37, 19–39. [Google Scholar] [CrossRef]
- Percival, G.C. The use of chlorophyll fluorescence to identify chemical and environmental stress in leaf tissue of three oak (Quercus) species. J. Arboric. 2005, 31, 215. [Google Scholar]
- Popovic, R.; Dewez, D.; Juneau, P. Applications of Chlorophyll Fluorescence in Ecotoxicology: Heavy Metals, Herbicides, and Air Pollutants. In Practical Applications of Chlorophyll Fluorescence in Plant Biology; Springer: Berlin/Heidelberg, Germany, 2003; pp. 151–184. [Google Scholar]
- Barbagallo, R.P.; Oxborough, K.; Pallett, K.E.; Baker, N.R. Rapid, noninvasive screening for perturbations of metab-olism and plant growth using chlorophyll fluorescence imaging. Plant Physiol. 2003, 132, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Soukupová, J.; Smatanová, S.; Nedbal, L.; Jegorov, A. Plant response to destruxins visualized by imaging of chlorophyll fluorescence. Physiol. Plant. 2003, 118, 399–405. [Google Scholar] [CrossRef]
- Hassannejad, S.; Lotfi, R.; Ghafarbi, S.P.; Oukarroum, A.; Abbasi, A.; Kalaji, H.M.; Rastogi, A. Early Identification of Herbicide Modes of Action by the Use of Chlorophyll Fluorescence Measurements. Plants 2020, 9, 529. [Google Scholar] [CrossRef]
- Chaerle, L.; Hagenbeek, D.; De Bruyne, E.; Van Der Straeten, M.; Straeten, D. Chlorophyll fluorescence imaging for disease-resistance screening of sugar beet. Plant Cell Tissue Organ Cult. (PCTOC) 2007, 91, 97–106. [Google Scholar] [CrossRef]
- Christen, D.; Schönmann, S.; Jermini, M.; Strasser, R.J.; Défago, G. Characterization and early detection of grapevine (Vitis vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environ. Exp. Bot. 2007, 60, 504–514. [Google Scholar] [CrossRef]
- Rosyara, U.R.; Subedi, S.; Duveiller, E.; Sharma, R.C. The effect of spot blotch and heat stress on variation of canopy temperature depression, chlorophyll fluorescence and chlorophyll content of hexaploid wheat genotypes. Euphytica 2010, 174, 377–390. [Google Scholar] [CrossRef]
- Cen, H.; Weng, H.; Yao, J.; He, M.; Lv, J.; Hua, S.; Li, H.; He, Y. Chlorophyll fluorescence imaging uncovers photosyn-thetic fingerprint of citrus Huanglongbing. Front. Plant Sci. 2017, 8, 1509. [Google Scholar] [CrossRef] [Green Version]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta (BBA) Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Feyziyev, Y.M. Chlorophyll fluorescence and “maximum quantum efficiency” of photosystem II in plant sciences. Life Sci. Biomed. 2020, 74, 18–28. [Google Scholar]
- Nedbal, L.; Whitmarsh, J. Chlorophyll Fluorescence Imaging of Leaves and Fruits. In Discoveries in Photosynthesis; Springer: Berlin/Heidelberg, Germany, 2004; pp. 389–407. [Google Scholar]
- Buschmann, C.; Lichtenthaler, H.K. Principles and characteristics of multi-colour fluorescence imaging of plants. J. Plant Physiol. 1998, 152, 297–314. [Google Scholar] [CrossRef]
- Sánchez-Moreiras, A.M.; Graña, E.; Reigosa, M.J.; Araniti, F. Imaging of chlorophyll a fluorescence in natural com-pound-induced stress detection. Front. Plant Sci. 2020, 11, 583590. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.F.; Kunz, C.; Peteinatos, G.G.; Santel, H.-J.; Gerhards, R. Utilization of Chlorophyll Fluorescence Imaging Technology to Detect Plant Injury by Herbicides in Sugar Beet and Soybean. Weed Technol. 2017, 31, 523–535. [Google Scholar] [CrossRef]
- Narayanan, M.; van Iersel, M.W.; Haidekker, M. Chlorophyll fluorescence imaging: A novel, simple and non-destructive method for canopy size imaging. HortScience 2019, 54, S179. [Google Scholar]
- Legendre, R.; van Iersel, M.W. Chlorophyll Fluorescence Imaging: A Novel, Low-Cost Method for Early Stress Detection. In Proceedings of the 2020 ASHS Annual Conference, Orlando, FL, USA, 10–13 August 2020. [Google Scholar]
- Agency, U.E.P. Atrazine, simazine, and cyanazine: Notice of initiation of special review. Fed. Reg. 1994, 59, 60412–60443. [Google Scholar]
- Shimabukuro, R.H.; Swanson, H.R. Atrazine metabolism, selectivity, and mode of action. J. Agric. Food Chem. 1969, 17, 199–205. [Google Scholar] [CrossRef]
- Murphy, B.P.; Larran, A.S.; Ackley, B.; Loux, M.M.; Tranel, P.J. Survey of glyphosate-, atrazine- and lactofen-resistance mechanisms in Ohio waterhemp (Amaranthus tuberculatus) populations. Weed Sci. 2019, 67, 296–302. [Google Scholar] [CrossRef]
- Sui, X.; Wu, Q.; Chang, W.; Fan, X.; Song, F. Proteomic analysis of the response of Funnelifor mismosseae/Medicago sativa to atrazine stress. BMC Plant Biol. 2018, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Iii, W.W.A.; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 2008, 98, 253–264. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Cohu, C.M.; Muller, O.; Adams, W.W. Modulation of photosynthetic energy conversion efficiency in nature: From seconds to seasons. Photosynth. Res. 2012, 113, 75–88. [Google Scholar] [CrossRef]
- Croteau, D.; Guérin, S.; Bruyant, F.; Ferland, J.; Campbell, D.A.; Babin, M.; Lavaud, J. Contrasting nonphotochemical quenching patterns under high light and darkness aligns with light niche occupancy in Arctic diatoms. Limnol. Oceanogr. 2021, 66, 231. [Google Scholar] [CrossRef]
- Heisel, F.; Sowinska, M.; Miehe, J.A.; Lang, M.; Lichtenthaler, H.K. Detection of Nutrient Deficiencies of Maize by Laser Induced Fluorescence Imaging. J. Plant Physiol. 1996, 148, 622–631. [Google Scholar] [CrossRef]
- Langsdorf, G.; Buschmann, C.; Sowinska, M.; Babani, F.; Mokry, M.; Timmermann, F.; Lichtenthaler, H. Multicolour Fluorescence Imaging of Sugar Beet Leaves with Different Nitrogen Status by Flash Lamp UV-Excitation. Photosynthetica 2000, 38, 539–551. [Google Scholar] [CrossRef]
- Tartachnyk, I.I.; Rademacher, I.; Kühbauch, W. Distinguishing nitrogen deficiency and fungal infection of winter wheat by laser-induced fluorescence. Precis. Agric. 2006, 7, 281–293. [Google Scholar] [CrossRef]
- Calatayud, A.; Roca, D.; Martínez, P. Spatial-temporal variations in rose leaves under water stress conditions studied by chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2006, 44, 564–573. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F. Detection of photosynthetic activity and water stressby imaging the red chlorophyll fluorescence. Plant Physiol. Biochem. 2000, 38, 889–895. [Google Scholar] [CrossRef]
- Massacci, A.; Nabiev, S.; Pietrosanti, L.; Nematov, S.; Chernikova, T.; Thor, K.; Leipner, J. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2008, 46, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Willits, D.; Peet, M. Measurement of Chlorophyll Fluorescence as a Heat Stress Indicator in Tomato: Laboratory and Greenhouse Comparisons. J. Am. Soc. Hortic. Sci. 2001, 126, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Harbinson, J. Insights on the development, kinetics, and variation of photoinhibition using chlorophyll fluorescence imaging of a chilled, variegated leaf. J. Exp. Bot. 2006, 58, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tevini, M. Plant Responses to Ultraviolet Radiation Stress. In Plant Cell Monographs; Springer International Publishing: Berlin/Heidelberg, Germany, 2007; Volume 19, pp. 605–621. [Google Scholar]
- Takayama, K.; Sakai, Y.; Nishina, H.; Omasa, K. Chlorophyll fluorescence imaging at 77 K for assessing the hetero-geneously distributed light stress over a leaf surface. Environ. Control Biol. 2007, 45, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Fuerst, E.P.; Norman, M.A. Interactions of Herbicides with Photosynthetic Electron Transport. Weed Sci. 1991, 39, 458–464. [Google Scholar] [CrossRef]
- Ruban, A.V. Evolution under the sun: Optimizing light harvesting in photosynthesis. J. Exp. Bot. 2015, 66, 7–23. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Trebst, A. The Mode of Action of Triazine Herbicides in Plants. Triazine Herbic. 2008, 50, 101–110. [Google Scholar] [CrossRef]
- Van Rensen, J.J.S. Action of Some Herbicides in Photosynthesis of Scenedesmus As Studied by Their Effects on Oxygen Evolution and Cyclic Photophosphorylation; Veenman: Wageningen, The Netherlands, 1971. [Google Scholar]
- Velthuys, B. Electron-dependent competition between plastoquinone and inhibitors for binding to photosystem II. FEBS Lett. 1981, 126, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, P.; Weber, J.F.; Gerhards, R. Early Identification of Herbicide Stress in Soybean (Glycine max (L.) Merr.) Using Chlorophyll Fluorescence Imaging Technology. Sensors 2018, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mopper, S.; Hasenstein, K.H. Effects of Salinity on Endogenous Aba, Iaa, Ja, and Sa in Iris hexagona. J. Chem. Ecol. 2001, 27, 327–342. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.; Daferera, D.; Polissiou, M.; Passam, H. The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Sci. Hortic. 2008, 115, 393–397. [Google Scholar] [CrossRef]
- Specht, J.E.; Chase, K.; Macrander, M.; Graef, G.L.; Chung, J.; Markwell, J.P.; Germann, M.; Orf, J.H.; Lark, K.G. Soybean Response to Water: A QTL Analysis of Drought Tolerance. Crop. Sci. 2001, 41, 493–509. [Google Scholar] [CrossRef]
- Bray, E.; Bailey-Serres, J.; Weretilnyk, E.; Buchannan, B.; Jones, R. Responses to abiotic stresses. In Biochemistry and Molecular Biology of Plants; American Society of Plant Biologists: Rockville, MD, USA, 2000; pp. 149–158. [Google Scholar]
- Mathobo, R.; Marais, D.; Steyn, J.M. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.; Rinderle, U. Chlorophyll fluorescence spectra of leaves as induced by blue light and red laser light. In Proceedings of the Spectral Signatures of Objects in Remote Sensing 1988, Aussois, France, 18–22 January 1988. [Google Scholar]
- Brewer, P.E.; Arntzen, C.J.; Slife, F.W. Effects of Atrazine, Cyanazine, and Procyazine on the Photochemical Reactions of Isolated Chloroplasts. Weed Sci. 1979, 27, 300–308. [Google Scholar] [CrossRef]
- Fan, X.; Chang, W.; Feng, F.; Song, F. Responses of photosynthesis-related parameters and chloroplast ultrastructure to atrazine in alfalfa (Medicago sativa L.) inoculated with arbuscular mycorrhizal fungi. Ecotoxicol. Environ. Saf. 2018, 166, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Iriel, A.; Novo, J.M.; Cordon, G.B.; Lagorio, M.G. Atrazine and Methyl Viologen Effects on Chlorophyll-a Fluores-cence Revisited—Implications in Photosystems Emission and Ecotoxicity Assessment. Photochem. Photobiol. 2014, 90, 107–112. [Google Scholar] [CrossRef]
- Ahrens, W.H.; Arntzen, C.J.; Stoller, E.W. Chlorophyll Fluorescence Assay for the Determination of Triazine Resistance. Weed Sci. 1981, 29, 316–322. [Google Scholar] [CrossRef]
- Conrad, R.; Büchel, C.; Wilhelm, C.; Arsalane, W.; Berkaloff, C.; Duval, J.-C. Changes in yield ofin-vivo fluorescence of chlorophyll a as a tool for selective herbicide monitoring. Environ. Boil. Fishes 1993, 5, 505–516. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, J.; Zhang, J.; Ren, L.; Mao, Y.; Li, B.; Zhang, M.; Liu, D.; Du, M. The impact of the herbicide atrazine on growth and photosynthesis of seagrass, Zostera marina (L.), seedlings. Mar. Pollut. Bull. 2011, 62, 1628–1631. [Google Scholar] [CrossRef] [PubMed]
- Gamon, J.; Peñuelas, J.; Field, C. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote. Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Garrity, S.R.; Vierling, L.A.; Bickford, K. A simple filtered photodiode instrument for continuous measurement of narrowband NDVI and PRI over vegetated canopies. Agric. For. Meteorol. 2010, 150, 489–496. [Google Scholar] [CrossRef]
- Legendre, R.; van Iersel, M.W. Supplemental Far-Red Light Stimulates Lettuce Growth: Disentangling Morphological and Physiological Effects. Plants 2021, 10, 166. [Google Scholar] [CrossRef]
- Cobb, A.H.; Reade, J.P. The Inhibition of Amino Acid Biosynthesis. In Herbicides and Plant Physiology; Wiley: Hoboken, NJ, USA, 2010; pp. 176–199. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legendre, R.; Basinger, N.T.; van Iersel, M.W. Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection. Sensors 2021, 21, 2055. https://doi.org/10.3390/s21062055
Legendre R, Basinger NT, van Iersel MW. Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection. Sensors. 2021; 21(6):2055. https://doi.org/10.3390/s21062055
Chicago/Turabian StyleLegendre, Reeve, Nicholas T. Basinger, and Marc W. van Iersel. 2021. "Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection" Sensors 21, no. 6: 2055. https://doi.org/10.3390/s21062055
APA StyleLegendre, R., Basinger, N. T., & van Iersel, M. W. (2021). Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection. Sensors, 21(6), 2055. https://doi.org/10.3390/s21062055