1. Introduction
Qualitative modeling of species habitat is best done with environmental data inputs that are relevant to the biogeography of the species of concern [
1]. However, the availability of digital data for these ecologically relevant factors is often limiting. Time and financial resources can constrain the effort to collect, analyze, and compile new data sources for digital maps. In addition, an ecologically relevant environmental factor may be difficult to capture directly. An alternative to omitting potentially important data sources in a modeling effort is to use existing data resources to develop best estimates or proxies for the environmental variable of concern.
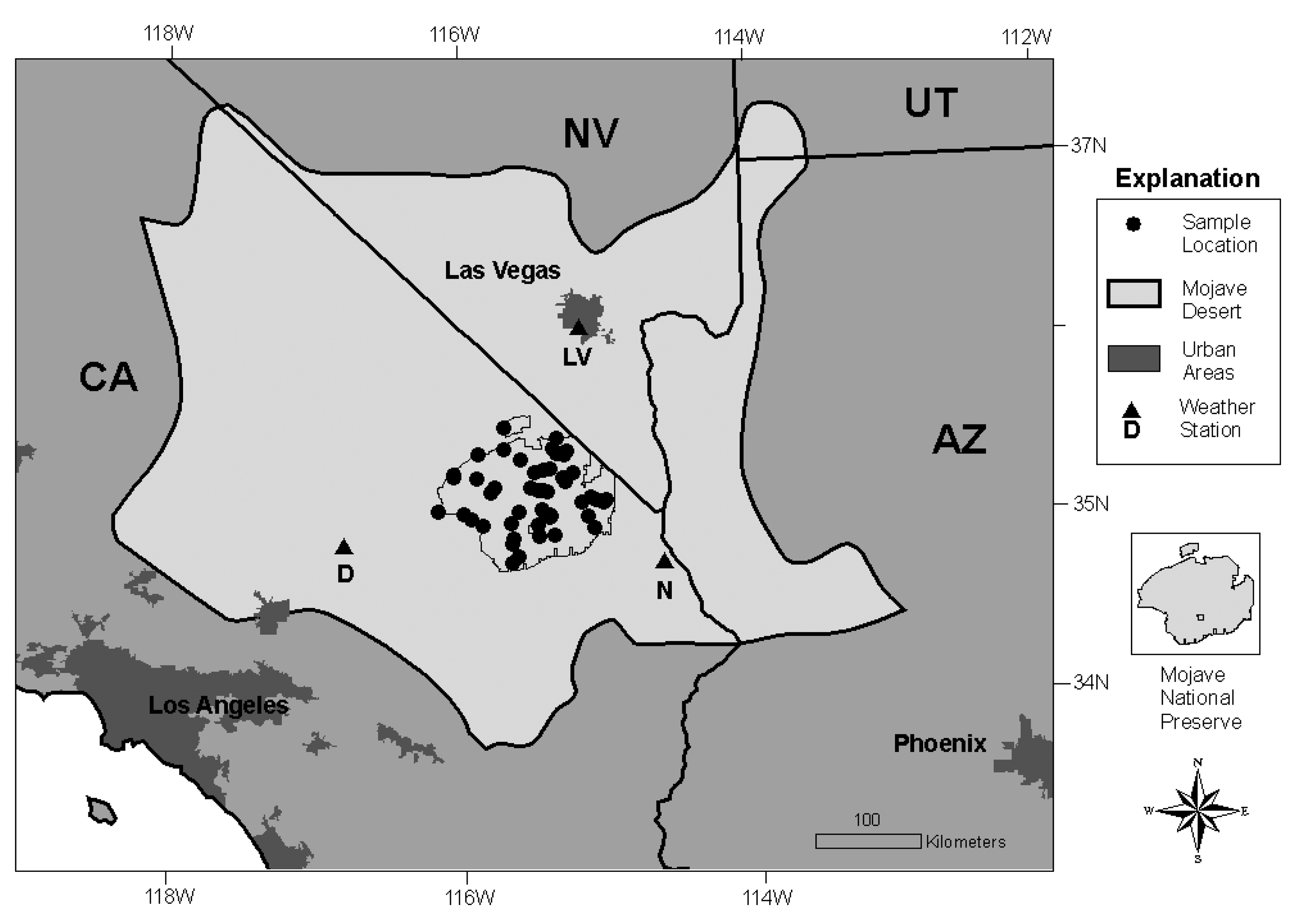
The Mojave Desert in the southwestern United States (U.S.) (
Figure 1) supports habitat for the federally listed Mojave Desert tortoise (
Gopherus agassizii) [
2]. The Department of Interior asked the U.S. Geological Survey to develop a potential habitat map for the desert tortoise using existing data inputs such as tortoise surveys, digital maps of environmental factors, and remotely sensed images. The assembled U.S. Geological Survey team included desert tortoise biologists who identified environmental factors known to be important to desert tortoise biology as suggested model inputs. Included among those factors was the potential of an area to support annual plants over time, a capacity we refer to as annual growth potential.
In the arid environment of the Mojave Desert, annual plant growth, as measured by annual cover, is largely dependent upon the timing and amount of rainfall [
3,
4]. Annual plants may be prolific in one year and not the next, and they may be prolific in one area of the Mojave and simultaneously absent in another suitable area within the Mojave. When annual plants are present, they are most obvious in the interspaces between sparsely spaced shrubs, but they can also occur under the shrubs in so-called “shrub islands” [
5]. The spatial, temporal, and compositional complexity of annual plants in the Mojave Desert has made mapping of annual plant cover at any one point in time technically complex and, for the purposes of characterizing tortoise habitat, ecologically uninformative.
In this paper, we present our approach to developing a proxy of annual plant growth for use as an environmental layer in desert tortoise habitat modeling. We developed provisional models using multiple Moderate-Resolution Imaging Spectroradiometer (MODIS) Enhanced Vegetation Index (EVI) satellite images and metrics designed to capture the plant phenology revealed in these data. Our proxy integrates apparent annual plant response over a number of years and expresses the abiotic potential of the landscape to support annual plant growth under sufficient precipitation.
2. Background
2.1. The Mojave Desert
The Mojave Desert encompasses about 115,000 km
2 of arid landscape in southern Nevada, western Arizona, southwestern Utah, and southeastern California (
Figure 1). Nearly two million people live in this desert, concentrated in large cities such as Las Vegas and Lancaster. However, forty million people in southern California and central Arizona are within a day's drive. Much of the Mojave Desert is under federal management, including four national park units, six major military training bases, and considerable acreage of BLM land.
The Mojave has cold winters and very hot summers [
6,
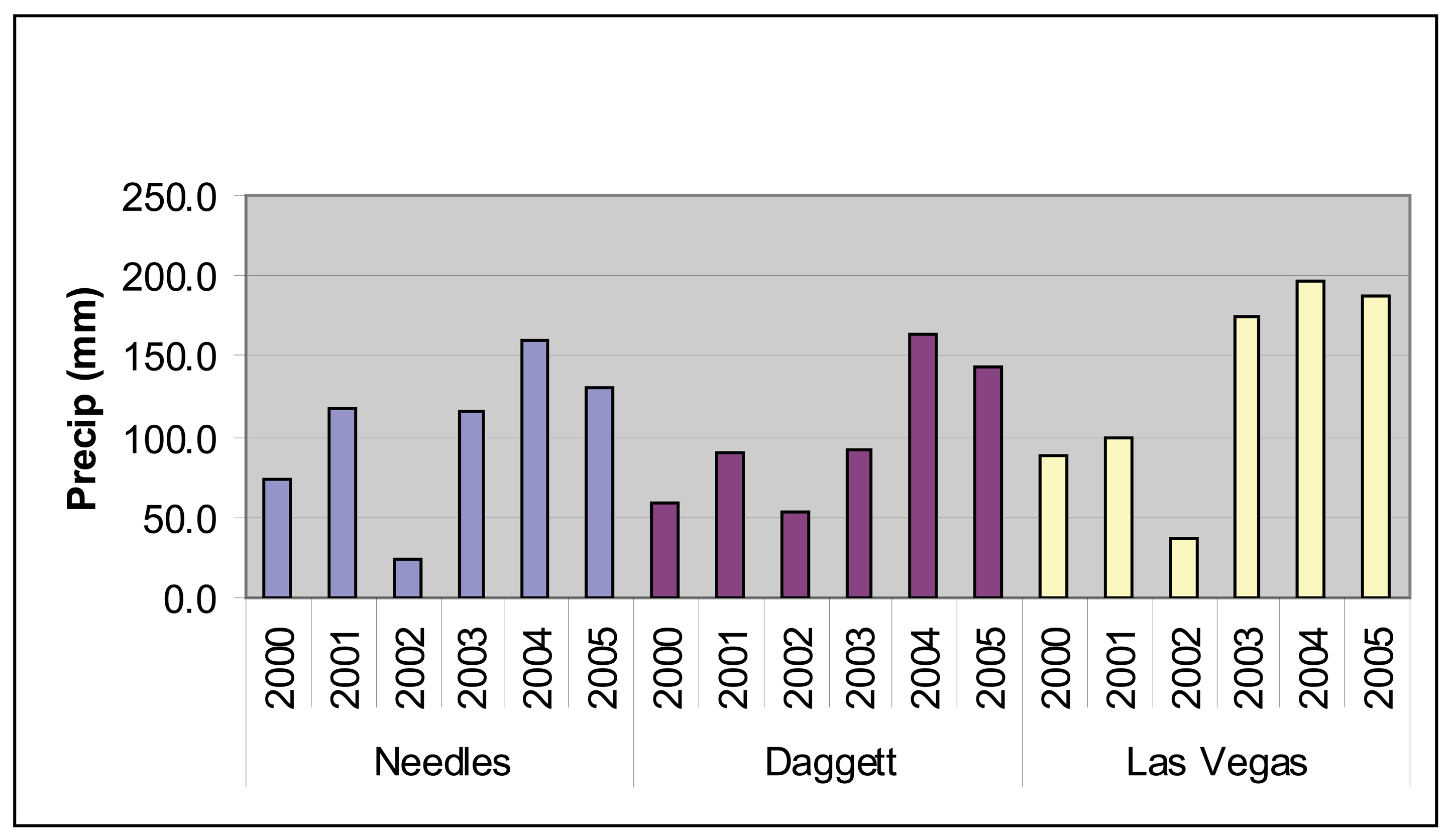
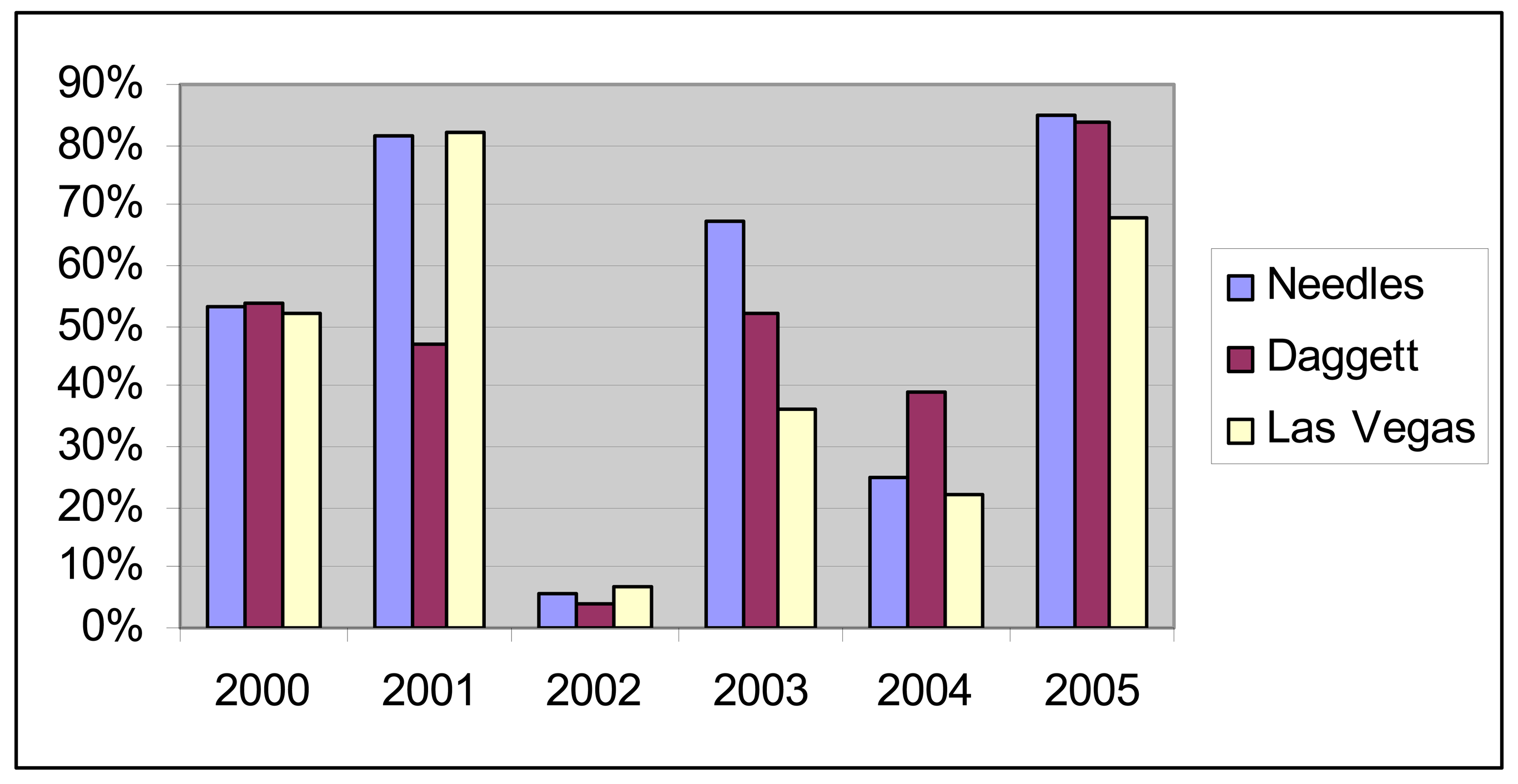
7]. Precipitation varies with elevation and as shown in
Figure 2, precipitation totals are both variable from year to year and geographically variable within a single year. Throughout the Mojave, 2002 was a particularly dry year while 2004 and 2005 had strong precipitation (
Figure 2). However, the spring of 2004 was uncharacteristically dry and most of the precipitation occurred later in the year (
Figure 3).
The Mojave Desert supports plant communities comprised of annual grasses and forbs, biennial and perennial shrubs, perennial grasses, and succulent plants such as agaves, cacti, and yuccas. The percentage of perennial plant cover is low and estimates based on remotely sensed images have indicated 28% of the Mojave has less than 10% perennial plant cover and 67% of the Mojave has less than 20% plant cover (Wallace, unpublished data). Annual plants consist of ephemeral native and non-native plants that appear seasonally when moisture is sufficient. Green-up of the annual plants is generally strongest in the early to mid spring [
7,
8,
9], after which the annual grasses and forbs generally dry up. When summer rains are adequate, there may be an additional green-up following summer rains, most often seen in the higher elevations of the eastern Mojave in response to summer storms coming from the southeast. Perennial plants likewise respond to precipitation. Many shrubs are drought deciduous and drop part or all of their leaves when rainfall is lower. Shrubs and small trees will green-up in the late spring when rainfall is adequate. Perennial grasses are generally a small component of each plant community and, as distinct plant communities, are found in only localized areas of the Mojave. The perennial grasses green-up later in the spring and summer compared to the annual plants. Succulents also generally occur as scattered components in plant communities except for a few areas in the Mojave. The succulents generally maintain leaves year round, when present, and do not display a distinct green-up period.
2.2. Satellite imagery
Many studies use satellite-derived vegetation indices, such as the Normalized Difference Vegetation Index (NDVI) [
14], coupled with field data to estimate total biomass, cover, or net primary productivity [
10-
13]. Multitemporal imagery, which allows the incorporation of phenological differences among image dates, is used increasingly to eliminate the problem of distinction between soil and vegetation signal in arid and semi-arid environments [
14-
16]. A number of studies derived greenness indices for arid grasslands from multitemporal images using MODIS, Advanced Very High Resolution Radiometer (AVHRR) or Landsat Thematic Mapper (TM) imagery [
17,
18,
19]. Peterson [
20] focused on a single plant species in Nevada and used two dates of Landsat data in a model trained with field-measured data to map the cover of
Bromus tectorum (cheatgrass), a nonnative grass. Wallace
et al. [
17] applied stepwise linear regressions between field measurements of perennial cover and MODIS-EVI data to model regional perennial vegetation cover in the Mojave Desert. These studies typically train their vegetation models with field data they collect, using the field data as the dependent variable and deriving the relationship to the image values used as independent variables in regression analysis.
MODIS- EVI images provide regional coverage, capture the dynamics of vegetation distributions across the landscape, are calibrated, and are freely [
21] available from the USGS Global Visualization Viewer website (
http://glovis.usgs.gov). These attributes help quantify even the small amounts of vegetation found in sparse deserts, because differences between the images are dominated by differences in vegetation phenology, with soils and topographic effects remaining relatively constant across adjacent images, if not across years.
MODIS data are collected daily at 250-meter resolution in the red (620-670 nm) and near-infrared bands (841-876 nm) and at 500-meter resolution for the blue band (459-479 nm) [
21]. The EVI product is a spectral measure of the amount of photosynthetically-active vegetation on the ground, calculated using the red, near-infrared (NIR), and blue bands, as follows:
Where
ρ is the atmospherically corrected or partially atmospherically-corrected (Rayleigh and ozone absorption) surface reflectance,
L is the canopy background adjustment that addresses nonlinear, differential NIR and red radiant transfer through a canopy, and
C1,
C2 are the coefficients of the aerosol resistance term, which uses the blue band to correct for aerosol influences in the red band. The coefficients adopted in the
EVI algorithm are,
L = 1,
C1 = 6,
C2 = 7.5, and
G (gain factor) = 2.5 [
16]. The
EVI equation optimizes the vegetation signal, de-couples the canopy background signal, and reduces atmospheric influences to allow for precise inter-comparisons of spatial and temporal variations in terrestrial photosynthetic activity [
16]. Daily MODIS-EVI data are combined into single 16-day composites using an improved Constrained View Maximum Value Composite (CV-MVC) scheme that reduces sun-target-sensor angular variations. Each year comprises twenty-three 16-day composites.
3. Data
3.1. MODIS-EVI images
MODIS-EVI satellite data provide a regional view of the Mojave ecosystem captured over 16-day intervals. Landscapes contain a mix of perennial and annual vegetation types that green up at rates and times characteristic for their distinct species, and these phenologies are captured by MODIS data [
17,
18,
20]. We used 115 MODIS-EVI images for 2001-2005.
EVI values are delivered as 16-bit numbers scaled to a valid range from -2,000 to 10,000, in which non-land surfaces (such as water or snow) typically assume negative values and land surfaces typically assume positive values [
16]. As landscapes become more densely vegetated, the calculated EVI approaches 10,000. To reduce data volumes, we rescaled the 16-bit data to 8-bit as follows:
This rescaling shifts the valid EVI 16-bit values to the range of 80 to 200, with a calculated EVI 16-bit = 0 shifted to EVI 8-bit = 100. We inspected each image for non-data (non-valid) values (flagged with EVI 16-bit = -3000) and observed only scattered pixels in water bodies. These non-data pixels are represented by an EVI 8-bit value of 70; they do not represent land surfaces and were not involved in model development.
3.2. Evaluation data
We used annual cover estimates made at observation sites in the Mojave National Preserve to evaluate the performance of the relative annual growth models. These data were collected for a previous study. The locations were initially selected using April 1992 Landsat Thematic Mapper (TM) imagery, which represented a wet month in a wet year for the Mojave Desert, to select locales with strong vegetation signature and nearby locales with weak vegetation signature. These sites excluded bedrock mountainous erosional highlands. Geographic coordinates of the sites were field recorded, and each site center was marked for relocation in the field.
In the spring of 2003 and 2005, field observers estimated the total perennial plant cover, total annual cover, non-native annual forb cover, and non-native annual grass cover at the selected sites. The visual estimates were for an area represented by a circle extending 50 meters from the site center (approximately .75 ha). The field crew, consisting of two experienced botanists, approximated the observation boundaries by pacing with a range finder. The observers calibrated their ocular estimates to a measured area on the ground and then made independent estimates of cover. Any differences in cover estimates were reconciled among the observers. In 2003, the sites were visited four times during the spring and the maximum annual cover among those visits was used. In 2005, 37 of the sites were visited once in late April. Not all 50 sites were revisited due to time and resource constraints. The 2003 field effort originated for a separate project and the 2005 revisits were done to capture the abundant annuals present that year for comparison.
4. Model development
4.1. Approach
In contrast to previous studies, in this project we created a number of provisional models of relative annual growth using satellite data alone, with field data used to evaluate rather than to calibrate the models. We identified groupings of the MODIS-EVI images that captured different growing season characteristics of annual plants in the Mojave and then developed twelve metrics that incorporated these groupings to create provisional annual growth models. Model performance was evaluated with the existing data collected in the Mojave in 2003 and repeat measures of the same sites in 2005.
4.2. Phenological groupings and reference points
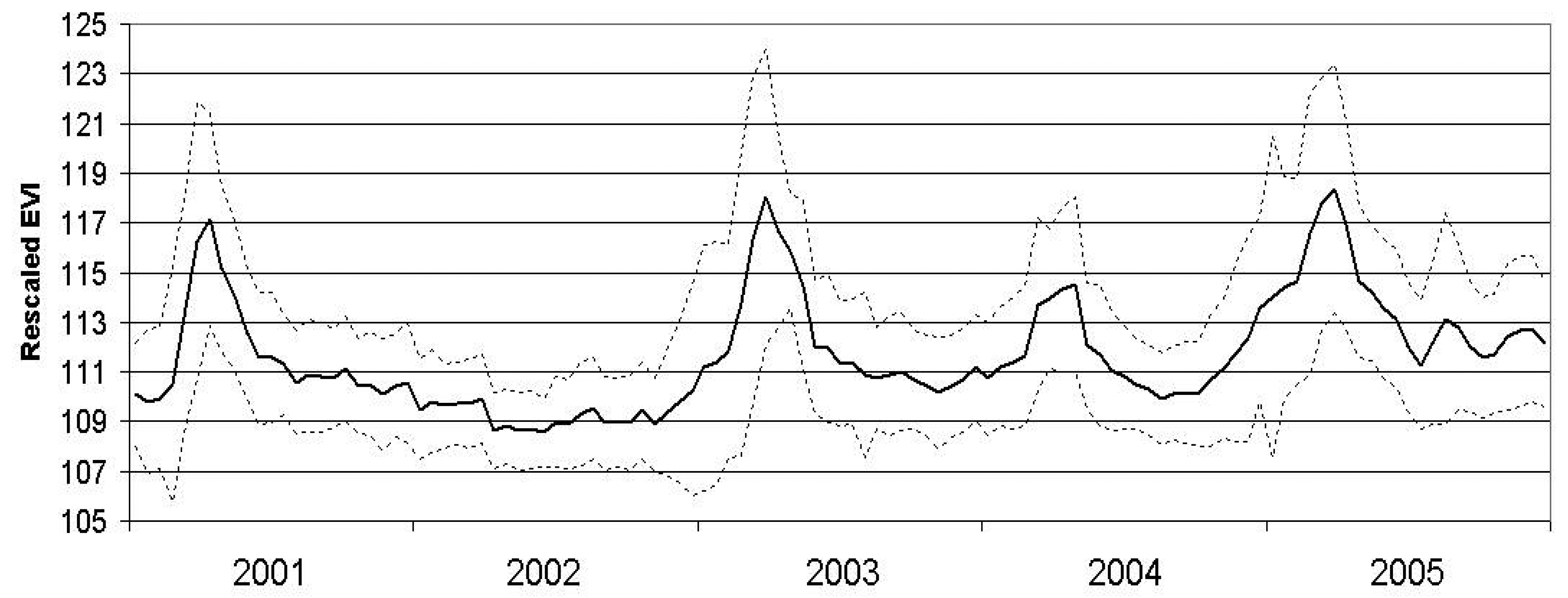
We inspected the MODIS-EVI values at all 50 field sites averaged from 2001 through 2005 to determine the relationship of the values with the expected phenological response of desert vegetation (
Figure 4).
We assembled the MODIS-EVI images into the following groupings based on the phenology profile above and input from desert ecologists, and roughly labeled them according to seasons:
Spring: Composites 4, 5, and 6 with composite start dates of February 18, March 6, and March 22, respectively. The Mojave Desert typically exhibits a flush of annual greenness in the early springtime and many areas host their most abundant annual cover at that time. The MODIS-EVI profile for that period is typically highest.
Early Summer: Composites 10, 11, 12, and 13 with composite start dates of May 25, June 10, June 26, and July 12, respectively. Annual plants in the Mojave Desert typically dry up as rainfall diminishes and the temperature rises. The MODIS-EVI profile for this period is typically lowest.
Winter: Composites 20-23 of the previous year (composite start dates November 1, 17 and December 3, 19) and composites 1-8 of the current year (composite start dates January 1, 17, February 2, 18, March 6, 22, and April 7, 23). Annual plants may begin to respond to precipitation as early as November of the previous year and grow through the early spring.
Late Summer: Composites 15, 16, and 17 with composite start dates of August 13, 29 and September 14. In some Mojave Desert landscapes, a pulse of annual vegetation may be present in the fall as a response to summer rains.
For the period of this study (2000 to 2005), the MODIS-EVI profile (
Figure 4) and field observations (Thomas pers. obs.) showed virtually no annual vegetation in 2002 and high annual vegetation in 2005. The year 2002 was an exceptionally dry year throughout the Mojave Desert and in particular for the critical growing season for annuals (
Figures 2 and
3). In comparison, 2005 was a wet year with high precipitation in the critical growing season following a wet year with lower precipitation in the critical growing season (
Figures 2 and
3). Based on the contrasts in image values and precipitation patterns between these two years, we identified 2002 as a reference year for very low annual growth and 2005 as a reference year for high annual growth.
Several of the provisional models we tested compare the average image values for a particular season for a year to that same season in 2002. Our rationale was that environmental factors such as sun angle, soils, substrate, and topographic shadowing remain constant for a given time of year, so subtracting the average image values observed in 2002 will compensate for the spectral contributions of these factors. Perennial plants also respond to drought, as evidenced by leaf drop and subdued green-up. However, unlike annual plants, which respond to extreme drought by not germinating, i.e. being nonexistent, the structure of a perennial plant remains and contributes to the satellite signal. Therefore, although differences in greenness of the perennial vegetation due to relative climate conditions are expected, we assume the signal of the annual vegetation is dominant and that the difference between the seasonal image values for a given year and the image values of 2002 will primarily reflect the relative amount of annual growth in a given year.
4.3. Annual growth metrics
Inspection of the MODIS-EVI profile (
Figure 4) reveals the vegetation “greenness” dynamics of the landscape. There are several ways to capture measures of this profile, including basic statistics, form fitting and Fourier transforms [
22-
25]. For this study, basic statistics were used to derive metrics based on seasonal groupings of selected MODIS composites and Fourier transforms were used to derive metrics based on the entire MODIS-EVI profile for each individual year. Applying Fourier analysis, we treat the ordered 23 EVI values at each pixel as a vector (or waveform) and extract the additive term (best fit of a flat line to the profile), and the first frequency magnitude and phase (the amplitude and timing of the peak for the best fit of a single sine wave to the profile). These measures capture information on vegetation vigor or density, timing of maximum greenness, and variability of greenness, respectively [
22,
23].
Table 1 shows the 12 metrics we derived and tested as possible measures related to annual growth.
4.4. Annual growth model development and evaluation
We created 24 provisional models of relative annual growth applying the 12 test metrics to MODIS-EVI images for 2002, 2003, and 2005 and evaluated the models using the field estimates of annual plant cover for 2003 and 2005. For each field site, we extracted the model values for 2005 (37 sites) and 2003 (50 sites) and calculated a linear regression between the field-estimated annual cover (percent) and the model predictions of annual growth. We assumed the provisional model that best predicted the measured annual-cover amounts at the field sites was superior. Clearly, there are potential problems with this assumption because: (1) the annual-cover estimates were collected in the local area of the Mojave National Preserve and are, therefore, not necessarily representative of the entire Mojave Desert and, (2) the field data were collected for a different study and were, therefore, not necessarily representative of 250m MODIS pixels (see [
10] and [
26] for MODIS sampling strategies). Given these potential problems, we consider a significant fit between the annual-growth model and the field data to be fortuitous and evidence that the metric applied was a reasonable approximation of relative annual growth.
5. Results
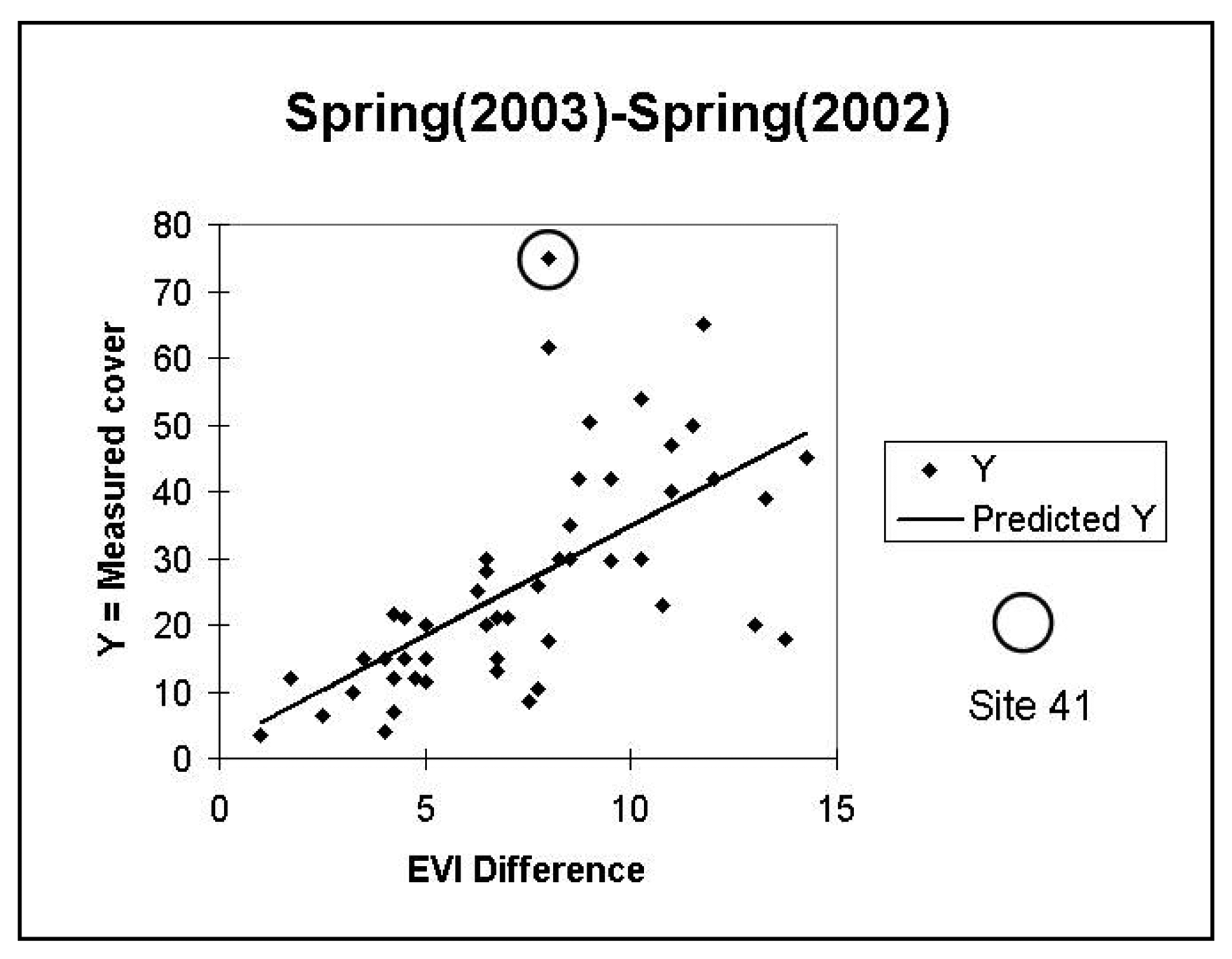
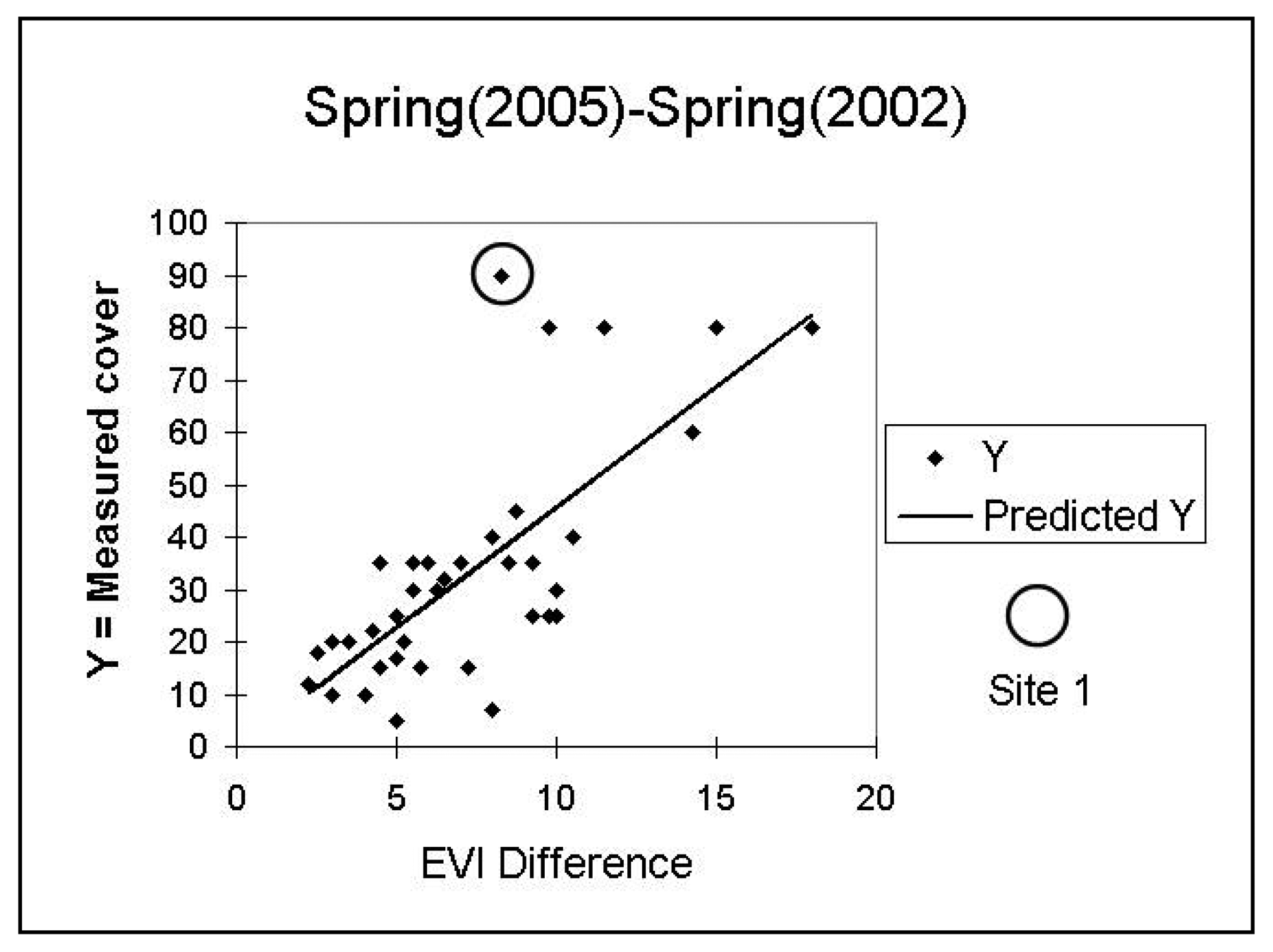
Initial investigations of the relationship of the field-estimated annual cover with model values revealed apparent outliers, site 41 for 2003 and site 1 for 2005 (see
Figures 5 and
6). The field notes for these sites reveal that they are anomalous. On 4/24/2003, site 41 had the highest total perennial plus annual coverage of all sites, at 93 %. On 4/21/05, site 1 had nearly 90% annual cover and, in contrast to other high-annual cover-sites that were dominated by forbs, it was composed of nearly all non-native
Schismus. The vegetation composition of other high cover sites was inspected and did not reveal any distinct compositional differences. We excluded sites 41 and 1 from further analysis.
Table 2 summarizes the results of the linear regressions between the suite of provisional relative annual growth models and the field-estimated annual cover data from 2003 and 2005. The highest regression result was R
2 = 0.61 significant at p < 0.01 between measured 2005 annual cover and the model based on the metric Average Spring (2005)-Average Spring (2002). This metric also gave the highest correlation (R
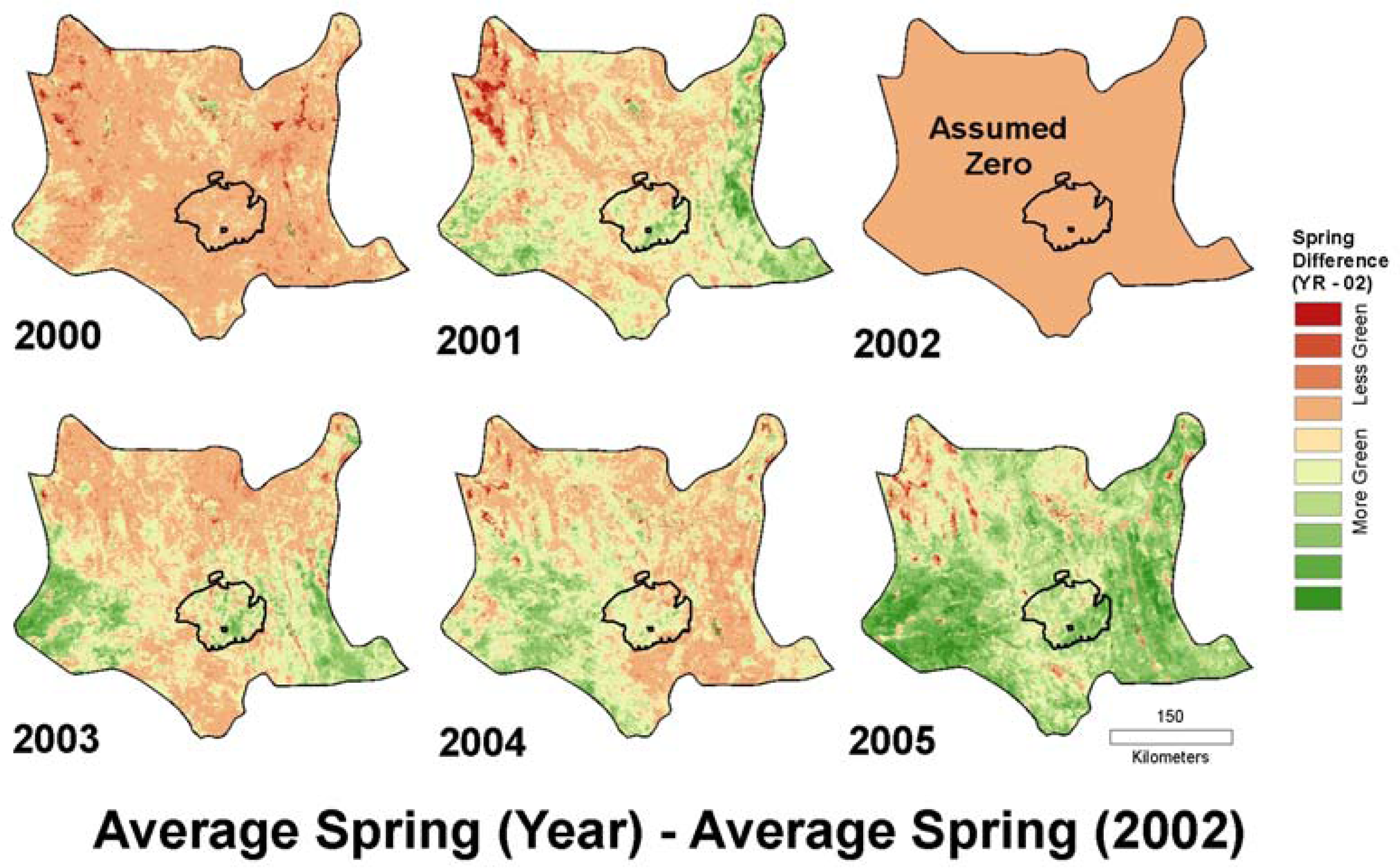
2 = 0.47, significant at p < 0.01) for field-estimated 2003 annual cover. The annual growth models represented by this spring difference metric for 2000 to 2005 are shown in
Figure 7.
Inspection of additional results (
Table 2) revealed several significant correlations, but with low R
2 in both years for annual growth models based on metrics that that included some part of the spring grouping and models based on the Fourier Magnitude difference. “Late Summer” based models were not correlated, “Early Summer” based models were poorly correlated, and the models based on “Winter” groupings were inconsistent.
Our original purpose for this study was to create a proxy for annual growth potential of the landscape for use in desert tortoise habitat modeling. We ultimately chose to use a rescaled version of the spring difference model calculated for the year 2005. Not only did this model have the highest R2, it is ecologically intuitive in that it evaluates the typical green-up period for annuals and captures the high contrast of annual response between an exceptionally wet spring (2005) and exceptionally dry spring (2002).
High elevation landscapes in the northwest Mojave Desert show low annual response for the 2005 spring difference model (
Figure 7), which is likely due to the presence of snow persisting at higher elevations later into the spring of 2005 compared to 2002. As an environmental layer for the tortoise, we maintained this contrast, since desert tortoises are not found at these higher elevations.
We rescaled the model values into 8-bit data with a range between 0 and 200 using the following formula:
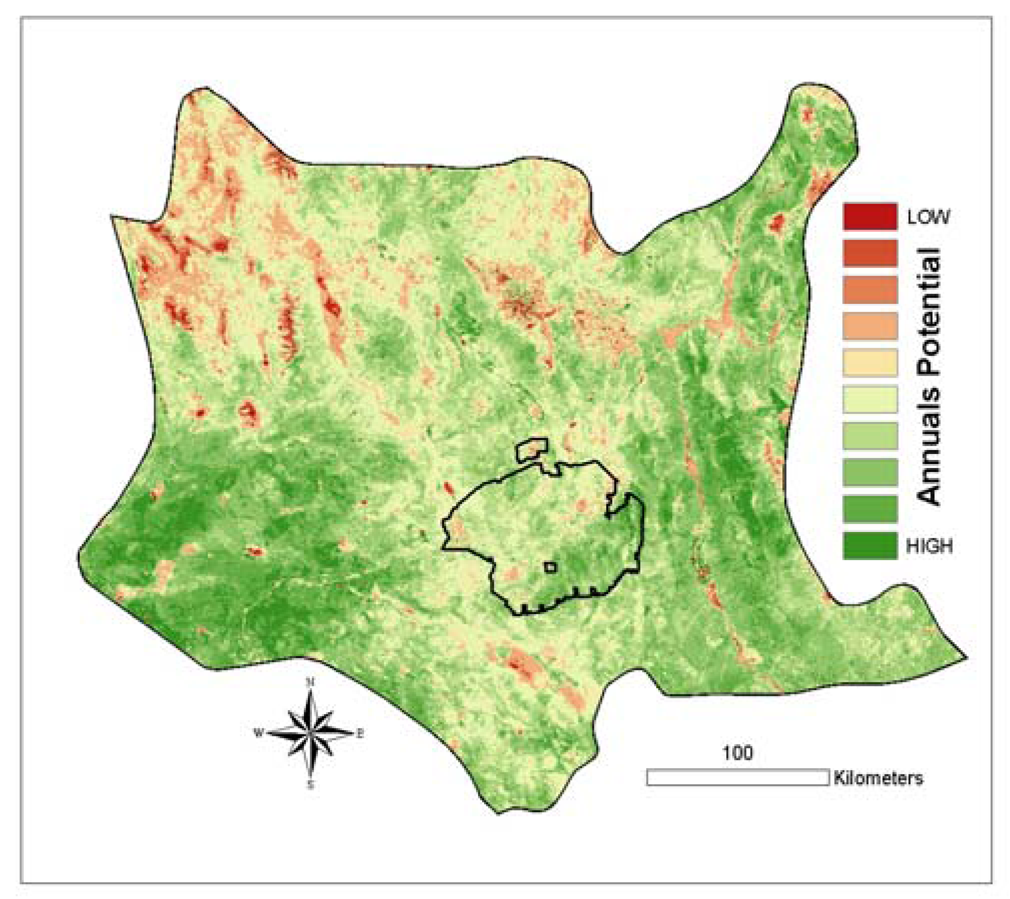
This formula is analogous to a Normalized Difference Vegetation Index (NDVI), which extracts a vegetation signal by contrasting the high near-infrared (NIR) reflectance with the low red (RED) reflectance value, calculated as: [(NIR-RED) / (NIR+RED)]. In our case (
equation 3), the high value is Spring 2005 greenness and the low value is Spring 2002 greenness. The resulting image (
Figure 8) was applied as the proxy in the tortoise modeling effort of the landscape's potential to support annual plants.
6. Discussion
We evaluated seasonal groupings of MODIS-EVI images and identified a metric to use as a functional proxy for annual growth potential that correlated with annual plant cover as estimated in a restricted portion of the Mojave Desert. Our study found the strongest correlation between measurements of annual plant cover and MODIS derivatives using the satellite metric defined as the difference between the average spring greenness for 2005, a wet year, and the average spring greenness for 2002, a year of record drought. These two years represent recent extremes in climate conditions, with essentially no annual plant production during the 2002 drought year and abundant annual plant growth during the record rainfall year of 2005. As such, the spring greenness observed in 2002 captures invariant environmental factors, including sun angle, soils, substrate, topographic shadowing and, to some extent, perennial vegetation cover. Our study created provisional models using basic statistics and Fourier Transforms, which capture information on selected portions of the annual greenness curve and on the entire curve, respectively. Other tools exist for developing phenological metrics and could be examined for refinement of this approach. For example, phenological metrics extracted using difference metrics, smoothing filters, and fitting functions calculated using TIMESAT [
27] are used to evaluate the effect of pre-fire fuels treatments and to monitor recovery in the Rodeo-Chediski fire perimeter of central Arizona [
24].
It is important to remember that this study used the field data only to evaluate the provisional satellite-based models and not for calibration in model development. Also remember these data are from the local area of the Mojave National Preserve and are not necessarily representative of the entire Mojave Desert. Because of these limitations, this model is not a calibrated map of annual vegetation cover, but is instead an environmental layer intended specifically for habitat modeling - a layer that captures a relative index of potential annual plant response. Although the model could be improved with additional field data collected using methods designed for MODIS pixels that represent all landscape types in the Mojave Desert, it is worth noting that this environmental data layer was found to be highly predictive of desert tortoise presence in the tortoise habitat modeling effort [Gass et al., in prep].
While the project had a pragmatic goal and was able to leverage limited resources, the study identified the value of using MODIS imagery from the spring of 2002 as a reference to index relative annual growth in other years. The metric Average Spring (Year) - Average Spring (2002) applied to MODIS-EVI images potentially provides a means to monitor annual growth in any year of consideration. This capacity could be of use in yearly fire-management planning, landscape-level invasive species surveys, and prediction of dust emissions in the Mojave. Since we could only provide a limited test of the efficacy of the spring difference metric, additional field work is needed to fully verify the approach and potentially to calibrate the approach to particular vegetation communities within the Mojave Desert.
Additional analysis of the yearly models created with the spring difference metric could include composites of the models displayed in
Figure 7 (e.g., retaining the highest or lowest greenness value at each pixel), measures of variability in annual growth across years (e.g. the range, coefficient of variation, and standard deviation) as well as trends in greenness for two or more years. Since the predictability of forage across years is an important consideration for any species, the landscape variability for annual growth captured by such metrics may also be useful in determining the distribution of desert herbivores.
7. Summary
Regional maps that provide information on annual plant cover are critical for informed natural-resource management of habitat because these plants are a seasonally fluctuating food base for many species. Despite the importance of such maps, their creation is hampered by the ephemeral nature of the vegetation they hope to capture and by the remoteness and extent of the areas for which they are needed, as well as by lack of funding to collect and analyze field data. Our study demonstrated that a successful proxy of annual growth potential can be created using MODIS-EVI satellite data alone by examining landscape phenologies revealed in these data combined with expert knowledge of annual plant seasonality. Field data were used to evaluate satellite-based models but were not required for model development or calibration. This study revealed the usefulness of contrasting two years of extremes-in our case comparing 2002 and 2005, a year of record drought with little evidence of annual growth and a year of exceptional spring precipitation with abundant annual growth, respectively. Our final model, found to be predictive of tortoise presence in the habitat modeling effort, is intuitively and ecologically satisfying in that it contrasts the greenness of the two extreme years during the season of expected maximum annual vegetation activity. This type of spatially distributed information is critical to many environmental tasks and is particularly well suited to the development of habitat suitability models for important or endangered species, in our case the desert tortoise.
Acknowledgments
The authors received funding for this project from the U.S. Geological Survey's Recoverability and Vulnerability of Desert Ecosystems, Global Climate Change, and D.O.I. Science on the Landscape programs. Robert H. Webb, Laura Norman, and Susan Benjamin provided thoughtful perspectives. We also thank field botanists Robert Hunt, Mimi Murov, and Keith Pohs for data collection in 2003 and 2005. We also thank two anonymous reviewers for valuable critical feedback.
References and Notes
- Austin, M.P. Spatial prediction of species distribution: an interface between ecological theory and statistical modeling. Eco. Model. 2002, 157, 101–118. [Google Scholar]
- Department of the Interior, Fish and Wildlife Service. 50 CFR part 17, RIN 1018-AB35. Endangered and Threatened Wildlife and Plants; Determination of threatened status for the Mojave population of the desert tortoise (final rule). Fed. Register 55 1990, 63, 12178–12191. [Google Scholar]
- Beatley, J.C. Phenological Events and their Environmental Triggers in Mojave Desert ecosystems. Ecology 1974, 55, 856–863. [Google Scholar]
- Mulroy, T.W.; Rundel, P.W. Annual plants: adaptations to desert environments. BioScience 1977, 27, 109–144. [Google Scholar]
- Noy-Meir, I. Desert ecosystem structure and function. In Hot deserts and arid shrublands; Evanari, M., Noy-Meir, I., Goodall, D.W., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1985; pp. 93–103. [Google Scholar]
- MacMahon, J.A.; Wagner, F.H. The mojave, sonoran and chihuahuan deserts of north america. In Hot deserts and arid shrublands; Evanari, M., Noy-Meir, I., Goodall, D.W., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1985; pp. 93–103. [Google Scholar]
- Rowlands, P.H.; Johnson, H.; Ritter, E.; Endo, A. The Mojave Desert. In Reference handbook on the deserts of North America; Bender, G.L., Ed.; Greenwood Press: Westport, Connecticut, 1982; pp. 103–162. [Google Scholar]
- Beatley, J.D. Biomass of desert winter annual plant populations in southern Nevada. Oikos 1969, 20, 261–273. [Google Scholar]
- Beatley, J.D. Phenological events and their environmental triggers in Mojave Desert ecosystems. Ecology 1974, 55, 856–863. [Google Scholar]
- Huete, A. R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar]
- Cohen, W.B.; Maiersperger, T.K.; Gower, S.T.; Turner, D.P. An improved strategy for regression of biophysical variables and Landsat ETM+ Data. Remote Sens. Environ. 2003, 84, 561–571. [Google Scholar]
- Chen, Z.M.; Babiker, I.S.; Chen, Z.X.; Komaki, K.; Mohamed, M.A.; Kato, K. Estimation of interannual variation in productivity of global vegetation using NDVI data. Int. J. Remote Sens. 2004, 25, 3139–3159. [Google Scholar]
- Rahman, M.M.; Csaplovics, E.; Koch, B. An efficient regression strategy for extracting forest biomass information from satellite sensor data. Int. J. Remote Sens. 2005, 26, 1511–1519. [Google Scholar]
- Huete, A.R; Jackson, R.D.; Post, D.F. Spectral response of a plant canopy with different soil backgrounds. Remote Sens. Environ. 1985, 17, 37–53. [Google Scholar]
- Tucker, C.J.; Sellers, P.J. Satellite remote sensing of primary production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar]
- Todd, S.W.; Hoffer, R.M.; Milchunas, D.G. Biomass estimation on grazed and ungrazed rangelands using spectral indices. Int. J. Remote Sens. 1998, 19, 427–438. [Google Scholar]
- Wallace, C.S.A.; Webb, R.H.; Thomas, K.A. Estimation of perennial vegetation cover distribution in the Mojave Desert using MODIS-EVI data. GIS Remote Sens. 2008, 2, 167–187. [Google Scholar]
- Tucker, C.J.; Vanparet, C.L. Satellite remote sensing of total herbaceous biomass in the Sengalese Sahel: 1980-1984. Remote Sens. Environ. 1985, 233–249. [Google Scholar]
- Kawamura, K.; Akiyama, T.; Watanabe, O.; Hasegawa, H.; Zhang, F.P.; Yokota, H.; Wang, S. Estimation of aboveground biomass in Xilingol Steppe, Inner Mongolia using NOAA/NDVI. Grassl. Sci. 2003, 49, 1–9. [Google Scholar]
- Peterson, E.B. Estimating cover of an invasive grass (Bromus tectorum) using tobit regression and phenology derived from two dates of Landsat ETM+ Data. Int. J. Remote Sens. 2005, 26, 2491–2507. [Google Scholar]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar]
- Jakubauskas, M.E.; Legates, D.R.; Kastens, J.H. Crop identification using harmonic analysis of time-series AVHRR NDVI data. Comp. Electronic. Agric. 2002, 37, 127–139. [Google Scholar]
- Moody, A.; Johnson, D.M. Land-surface phenologies from AVHRR using the discrete Fourier transform. Remote Sens. Environ. 2001, 75, 305–323. [Google Scholar]
- van Leeuwen, Willem J.D. Monitoring the Effects of Forest Restoration Treatments on Post-Fire Vegetation Recovery with MODIS Multitemporal Data. Sensors 2008, 8, 2017–2042. [Google Scholar]
- Reed, B.C.; White, M.A.; Brown, J.F. Remote sensing phenology. In Phenology: An Integrative Environmental Science; Shwartz, M.D., Ed.; Kluwer Publishing: Dordrecht, Netherlands, 2003. [Google Scholar]
- Hansen, M.C.; Townshend, R.G.; DeFries, R.; Carroll, M. Estimation of tree cover using MODIS data at global, continental, and regional/local scales. Int. J. Remote Sens. 2005, 26, 4359–4380. [Google Scholar]
- Jönsson, P.; Eklundh, L. TIMESAT-a program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar]
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}