Optoelectronic Plethysmography has Improved our Knowledge of Respiratory Physiology and Pathophysiology

{kind=link}
{kind=link}
Abstract
:1. Introduction
- Accurate computation of volume changes without using a mouthpiece that may alter the normal breathing pattern [3].
- Need of a simple, stable and repeatable calibration.
- Possibility of use in non-collaborating subjects (during sleep, or in unconscious patients).
- Permitting the analysis in different postures.
- Permitting the analysis under dynamic conditions such as walking or cycling.
- Allowing high frequency response in order to accurately describe rapid phenomena (i.e. electric or magnetic stimulation of phrenic nerves).
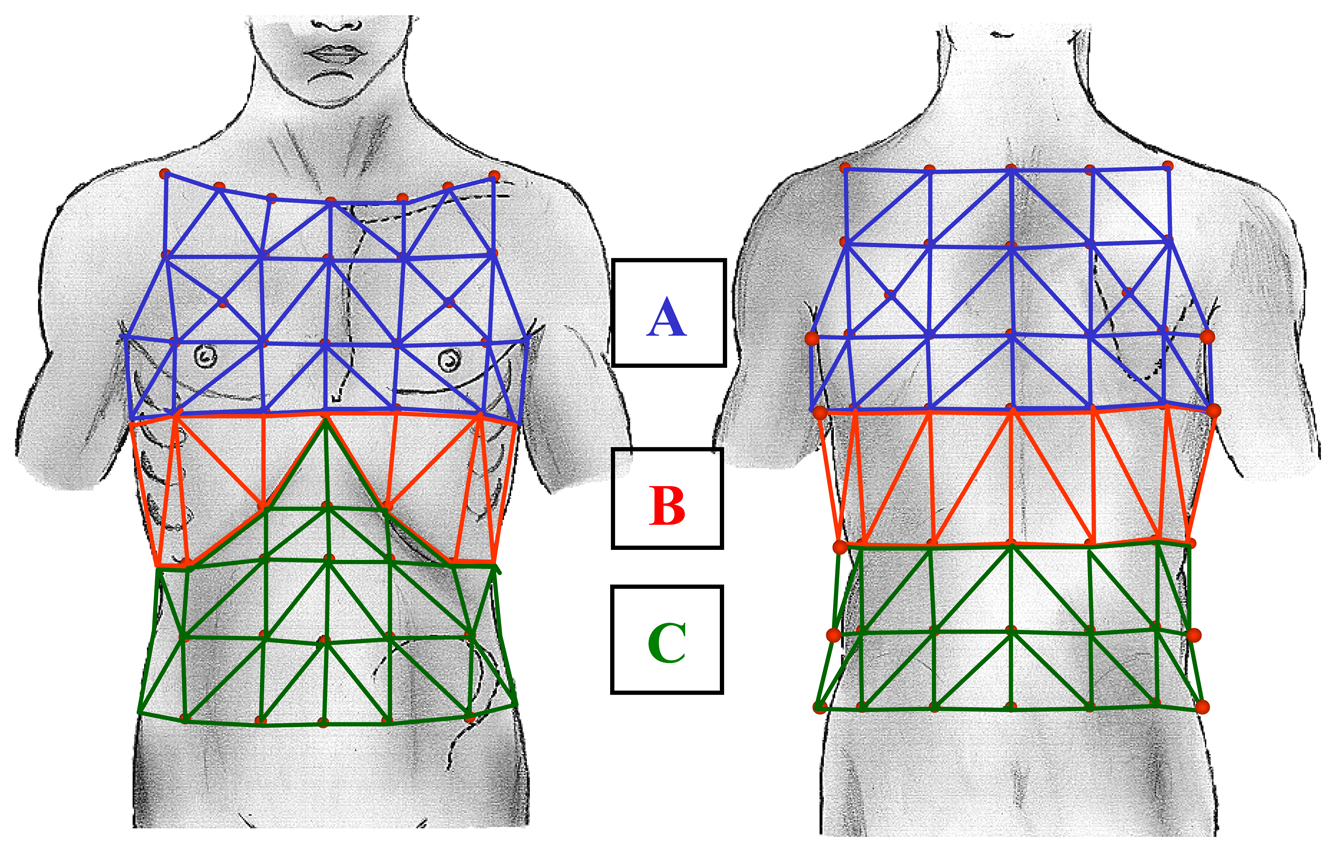
- Allowing the analysis of movements and volume changing of the different compartments of the chest wall: the upper thorax, lower thorax, and abdomen).
- Allowing the analysis of movements and volume changing of the two half (left and right) of the chest wall.
- Being non-invasive and safe for the patient.
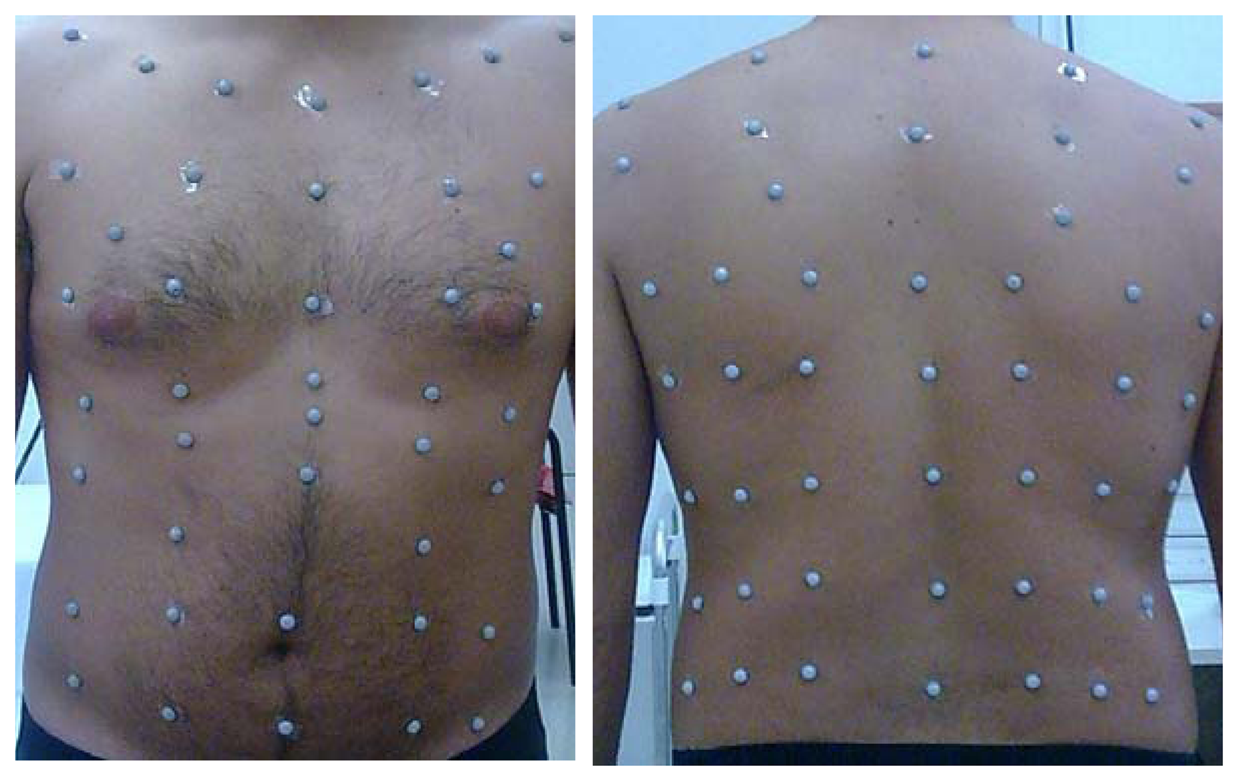
2. Methods
3. Applications
3.1. Physiology
3.1.1. Basic Physiologic Approach to Chest Wall Dynamics during Exercise
3.1.2. Chest Wall and Respiratory Effort during Walking
3.1.3. Chest wall dynamics during externally imposed expiratory flow limitation
3.1.4. Chest Wall Dynamics during Induced Hypercapnia
3.1.5. Chest Wall Kinematics and Mechanics during Arm Exercise. A Comparison with Leg Exercise
3.1.6. Phonetic Tasks (Speech Activities)
3.1.7. Chest Wall Kinematics during Laughter and Playing
3.1.8. Chest Wall Kinematics during Coughing
3.2. Clinical Applications
3.2.1. Asthma
3.2.2. Chronic Obstructive Pulmonary Disease (COPD)
3.2.2.1. Methods
3.2.2.2. Clinics
3.2.2.3. Physical Activities
3.2.3. Pathology of the Rib Cage
3.2.4. Neuromuscolar Disease
3.2.4.1. Chest Wall Kinematics in Patients with Hemiplegia
3.2.4.2. Duchenne Muscular Dystrophy [DMD]
3.2.4.3. Cough ineffectiveness
3.2.5. OEP in Intensive Care Unit
4. Conclusions
References and Notes
- Smith, J.C.; Mead, J. Three degree of freedom description of movement of the human chest wall. J. Appl. Physiol. 1986, 60, 928–934. [Google Scholar]
- Sackner, M.A. Monitoring of ventilation without a physical connection to the airway. In Diagnostic Thechniques in Pulmonary Disease. Part I; Marcel Dekker Inc.: New York, USA, 1980. [Google Scholar]
- Gilbert, R.; Auchincloss, J.H., Jr.; Brodsky, J.; Boden, W. Changes in tidal volume, frequency and ventilation induced by their measurement. J. Appl. Physiol. 1972, 33, 252–254. [Google Scholar]
- Pedotti, A.; Ferrigno, G. Opto-electronics based systems. In Three-Dimensional Analysis of Human Movement, Human Kinetics, 1st Ed.; Allard, P., Stokes, I.A., Bianchi, J.P., Eds.; Human Kinetics Publishers: Champaign, USA, 1995; pp. 57–78. [Google Scholar]
- Cala, S.J.; Kenyon, C.M.; Ferrigno, G.; Carnevali, P.; Aliverti, A.; Pedotti, A.; Macklem, P.T.; Rochester, D.F. Chest wall and lung volume estimation by optical reflectance motion analysis. J. Appl. Physiol. 1996, 81, 2680–2689. [Google Scholar]
- Kenyon, C.M.; Cala, S.J.; Yan, S.; Aliverti, A.; Scano, G.; Duranti, R.; Pedotti, A.; Macklem, P.T. Rib cage mechanics during quiet breathing and exercise in humans. J. Appl. Physiol. 1997, 83, 1242–1255. [Google Scholar]
- Aliverti, A.; Cala, S.J.; Duranti, R.; Ferrigno, G.; Kenyon, C.M.; Pedotti, A.; Scano, G.; Sliwinsky, P.; Macklem, P.T.; Yan, S. Human respiratory muscle actions and control during exercise. J. Appl. Physiol. 1997, 83, 1256–1269. [Google Scholar]
- Ward, M.E.; Ward, J.W.; Macklem, P.T. Analysis of human chest wall motion using a two compartment rib-cage model. J. Appl. Physiol. 1992, 72, 1338–1347. [Google Scholar]
- Sanna, A.; Bertoli, F.; Misuri, G.; Gigliotti, F.; Iandelli, I.; Mancini, M.; Duranti, R.; Ambrosino, N.; Scano, G. Chest wall kinematics and respiratory muscle action in walking healthy man. J. Appl. Physiol. 1999, 87, 938–946. [Google Scholar]
- Druz, W.S.; Sharp, J.T. Electrical and mechanical activity of the diaphragm accompanying body position in severe chronic obstructive pulmonary disease. Am. Rev. Respir. Dis. 1982, 125, 275–280. [Google Scholar]
- Banzett, R.B.; Topulos, G.P.; Leith, D.E.; Nations, C.S. Bracing arms increases the capacity for sustained hyperpnea. Am. Rev. Respir. Dis. 1988, 138, 106–109. [Google Scholar]
- De Troyer, A.; Pride, N. The chest wall and respiratory muscles in chronic obstructive pulmonary disease. In The thorax, part C, Disease; Roussos, C., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1995; Part C; pp. 1975–2006. [Google Scholar]
- Duranti, R.; Sanna, A.; Romagnoli, I.; Nerini, M.; Gigliotti, F.; Ambrosino, N.; Scano, G. Walking modality affects respiratory muscle action and contribution to respiratory effort. Pflugers. Arch. 2004, 448, 222–230. [Google Scholar]
- Aliverti, A.; Iandelli, I.; Duranti, R.; Cala, S.J.; Kayser, B.; Kelly, S.; Misuri, G.; Pedotti, A.; Scano, G.; Sliwinski, P.; Yan, S.; Macklem, P.T. Respiratory muscle dynamics and control during exercise with externally imposed expiratory flow limitation. J. Appl. Physiol. 2002, 92, 1953–1963. [Google Scholar]
- Iandelli, I.; Aliverti, A.; Kayser, B.; Dellacà, R.; Cala, S.J.; Duranti, R.; Kelly, S.; Scano, G.; Sliwinski, P.; Yan, S.; Macklem, P.T.; Pedotti, A. Determinants of exercise performance in normal men with externally imposed expiratory flow limitation. J. Appl. Physiol. 2002, 92, 1943–1952. [Google Scholar]
- Aliverti, A.; Dellacà, R.; Lotti, P.; Bertini, S.; Duranti, R.; Scano, G.; Heyman, J.; Lo Mauro, A.; Pedotti, A.; Macklem, P.T. Influence of expiratory flow-limitation during exercise on systemic oxygen delivery in humans. Eur. J. Appl. Physiol. 2005, 95, 229–242. [Google Scholar]
- Aliverti, A.; Ghidoli, G.; Dellacà, R.; Pedotti, A.; Macklem, P.T. Chest wall kinematic determinants of diaphragm length by optoelectronic plethysmography and ultrasonography. J. Appl. Physiol. 2003, 94, 621–630. [Google Scholar]
- Yan, S.; Sliwinski, P.; Macklem, P.T. Association of chest wall motion and tidal volume responses during CO2 rebreathing. J. Appl. Physiol. 1996, 81, 1528–1534. [Google Scholar]
- Gorini, M.; Corrado, A.; Aito, S.; Ginanni, R.; Villella, G.; Lucchesi, G.; De Paola, E. Ventilatory and respiratory muscle responses to hypercapnia in patients with paraplegia. Am. J. Respir. Crit. Care Med. 2000, 162, 203–208. [Google Scholar]
- Henke, K.G.; Sharratt, M.; Pegelow, D.; Dempsey, J.A. Regulation of end expiratory lung volume during exercise. J. Appl. Physiol. 1988, 64, 135–146. [Google Scholar]
- Romagnoli, I.; Gigliotti, F.; Lanini, B.; Bianchi, R.; Soldani, N.; Nerini, M.; Duranti, R.; Scano, G. Chest wall kinematics and respiratory muscle coordinated action during hypercapnia in healthy males. Eur. J. Appl. Physiol. 2004, 91, 525–533. [Google Scholar]
- Celli, B.R.; Criner, G.; Rassulo, J. Ventilatory muscle recruitment during unsupported arm exercise in normal subjects. J. Appl. Physiol. 1988, 54, 1936–1941. [Google Scholar]
- Romagnoli, I.; Gorini, M.; Gigliotti, F.; Bianchi, R.; Lanini, B.; Grazzini, M.; Stendardi, L.; Scano, G. Chest wall kinematics, respiratory muscle action and dyspnoea during arm vs. leg exercise in humans. Acta. Physiol. (Oxf) 2006, 188, 63–73. [Google Scholar]
- Bradley, T.D.; Chartrand, D.A.; Fitting, J.W.; Killian, K.J.; Grassino, A. The relation of inspiratory effort sensation, fatiguing patterns of diaphragm. Am. Rev. Respir. Dis. 1986, 134, 1119–1124. [Google Scholar]
- Ward, M.E.; Eidelman, D.; Stubbing, D.; Bellemare, F.; Macklem, P.T. Respiratory sensation and pattern of respiratory muscle activation during diaphragm fatigue. J. Appl. Physiol. 1988, 65, 2181–2189. [Google Scholar]
- Kayser, B.; Sliwinski, P.; Yan, S.; Tobiasz, M.; Macklem, P.T. Respiratory effort sensation during exercise with induced expiratory-flow limitation in healthy humans. J. Appl. Physiol. 1997, 83, 936–947. [Google Scholar]
- Muza, S.R.; McDonald, S.; Zechman, F.W. Comparison subjects' perception of inspiratory and expiratory resistance. J. Appl. Physiol. 1984, 56, 211–216. [Google Scholar]
- Romagnoli, I.; Gigliotti, F.; Galarducci, A.; Lanini, B.; Bianchi, R.; Cammelli, D.; Scano, G. Chest wall kinematics and respiratory muscle action in ankylosing spondylitis patients. Eur. Respir. J. 2004, 24, 453–460. [Google Scholar]
- Bunn, J.C.; Mead, J. Control of ventilation during speech. J. Appl. Physiol. 1971, 3, 870–872. [Google Scholar]
- Bouhuys, A.; Proctor, D.F.; Mead, J. Kinetics aspects of singing. J. Appl. Physiol. 1966, 21, 483–496. [Google Scholar]
- Hoit, J.D.; Hixon, T.J.; Altman, M.E.; Morgan, W.J. Speech breathing in women. J. Speech Hear Res. 1989, 32, 353–365. [Google Scholar]
- Binazzi, B.; Lanini, B.; Bianchi, R.; Romagnoli, I.; Nerini, M.; Gigliotti, F.; Duranti, R.; Milic-Emili, J.; Scano, G. Breathing pattern and kinematics in normal subjects during speech, singing and loud whispering. Acta. Physiol. (Oxf) 2006, 186, 233–246. [Google Scholar]
- Citardi, M.J.; Yanagisawa, E.; Estill, J. Videoendoscopic analysis of laryngeal function during laughter. Ann. Otol. Rhinol. Laryngol. 1996, 105, 545–549. [Google Scholar]
- Cox, S.V.; Eisenhauer, A.C.; Hreib, K. Seinfeld syncope. Cathet. Cardiovasc. Diagn. 1997, 42, 242, PMID: 9328725. [Google Scholar]
- Fredberg, J.J.; Jones, K.A.; Nathan, M.; Raboudi, S.; Prakash, Y.S.; Shore, S.A.; Butler, J.P.; Sieck, G.C. Friction in airway smooth muscle: mechanism, latch and implications in asthma. J. Appl. Physiol. 1996, 81, 2703–2712. [Google Scholar]
- Filippelli, M.; Pellegrino, R.; Iandelli, I.; Misuri, G.; Rodarte, J.R.; Duranti, R.; Brusasco, V.; Scano, G. Respiratory dynamics during laughter. J. Appl. Physiol. 2001, 90, 1441–1446. [Google Scholar]
- Cossette, I.; Monaco, P.; Aliverti, A.; Macklem, P.T. Chest wall dynamics and muscle recruitment during professional flute playing. Respir. Physiol. Neurobiol. 2008, 160, 187–195. [Google Scholar]
- Man, W.D.; Kyroussis, D.; Fleming, T.A.; Chetta, A.; Harraf, F.; Mustfa, N.; Rafferty, G.F.; Polkey, M.I.; Moxham, J. Cough gastric pressure and maximum expiratory mouth pressure in humans. Am. J. Respir. Crit. Care Med. 2003, 168, 714–717. [Google Scholar]
- Lanini, B.; Bianchi, R.; Binazzi, B.; Romagnoli, I.; Pala, F.; Gigliotti, F.; Scano, G. Chest wall kinematics during cough in healthy subjects. Acta. Physiol. (Oxford) 2007, 190, 351–358. [Google Scholar]
- Morris, A.J.R.; Siafakas, N.; Green, M. Thoracoabdominal motion and pressures during coughing (abstract). Thorax 1979, 34, 421. [Google Scholar]
- Gorini, M.; Iandelli, I.; Misuri, G.; Bertoli, F.; Filippelli, M.; Mancini, M.; Duranti, R.; Gigliotti, F.; Scano, G. Chest wall hyperinflation during acute bronchoconstriction in asthma. Am. J. Respir. Crit. Care Med. 1999, 160, 808–816. [Google Scholar]
- Lougheed, M.D.; Lam, M.; Forkert, L.; Webb, K.A.; O'Donnell, D.E. Breathlessness during acute bronchoconstriction in asthma. Am. Rev. Respir. Dis. 1993, 148, 1452–1459. [Google Scholar]
- Filippelli, M.; Duranti, R.; Gigliotti, F.; Bianchi, R.; Grazzini, M.; Stendardi, L.; Scano, G. Overall contribution of chest wall hyperinflation to breathlessness in asthma. Chest 2003, 124, 2164–2170. [Google Scholar]
- O'Donnell, D.E.; Lam, M.; Webb, K.A. Spirometric correlates of improvement in exercise performance after anticholinergic therapy in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, 542–549. [Google Scholar]
- Pellegrino, R.; Brusasco, V. Lung hyperinflation and flow limitation in chronic airway obstruction. Eur. Respir. J. 1997, 10, 543–549. [Google Scholar]
- Pellegrino, R.; Rodarte, J.R.; Brusasco, V. Assessing the reversibility of airway obstruction. Chest 1998, 114, 1607–1612. [Google Scholar]
- Tantucci, C.; Duguet, A.; Similowski, T.; Zelter, M.; Derenne, J.P.; Milic-Emili, J. Effect of salbutamol on dynamic hyperinflation in chronic obstructive pulmonary disease patients. Eur. Respir. J. 1998, 12, 799–804. [Google Scholar]
- Duranti, R.; Filippelli, M.; Bianchi, R.; Romagnoli, I.; Pellegrino, R.; Brusisco, V.; Scano, G. Inspiratory capacity and decrease in lung hyperinflation with albuterol in COPD. Chest 2002, 122, 2009–2014. [Google Scholar]
- American Thoracic Society. Pulmonary rehabilitation, 1999. Am. J. Respir. Crit. Care Med. 1999, 159, 1666–1682. [Google Scholar]
- Breslin, E.H. The pattern of respiratory muscle recruitment during pursed-lips breathing COPD. Chest 1992, 101, 75–78. [Google Scholar]
- Mueller, R.E.; Petty, T.L.; Filley, G.F. Ventilation and arterial blood gas changes induced by pursed lips breathing. J. Appl. Physiol. 1970, 28, 784–789. [Google Scholar]
- Thoman, R.L.; Stoker, G.L.; Ross, J.C. The efficacy of PLB in patients with chronic obstructive pulmonary disease. Am. Rev. Respir. Dis. 1966, 93, 100–106. [Google Scholar]
- Casciari, R.J.; Fairshter, R.D.; Harrison, A.; Morrison, J.T.; Blackburn, C.; Wilson, A.F. Effects of breathing retraining in patients with COPD. Chest 1981, 79, 393–398. [Google Scholar]
- Ingram, R.H.; Schilder, D.P. Effects of pursed lip expiration on the pulmonary pressure-flow relationship in obstructive lung disease. Am. Rev. Respir. Dis. 1967, 96, 381–388. [Google Scholar]
- Sharp, J.T.; Goldberg, N.B.; Druz, W.S.; Danon, J. Respiratory muscle function in patients with chronic obstructive pulmonary disease: its relationship to disability and to respiratory therapy. Am. Rev. Respir. Dis. 1974, 110, 154–167. [Google Scholar]
- American Thoracic Society. Dyspnea: mechanisms, assessment, and management; a consensus statement. Am. J. Respir. Crit. Care Med. 1999, 159, 321–340. [Google Scholar]
- Martinez, F.J.; de Oca, M.M.; Whyte, R.I.; Stetz, J.; Gay, S.E.; Celli, B.R. Lung volume reduction improves dyspnea, dynamic hyperinflation, and respiratory muscle function. Am. J. Respir. Crit. Care Med. 1997, 155, 1984–1990. [Google Scholar]
- Bianchi, R.; Gigliotti, F.; Romagnoli, I.; Lanini, B.; Castellani, C.; Grazzini, M.; Scano, G. Chest wall kinematics and breathlessness during pursed-lip breathing in patients with COPD. Chest 2004, 125, 459–465. [Google Scholar]
- Bianchi, R.; Gigliotti, F.; Romagnoli, I.; Lanini, B.; Castellani, C.; Binazzi, B.; Stendardi, L.; Grazzini, M.; Scano, G. Patterns of chest wall kinematics during volitional pursed-lip breathing in COPD at rest. Respir. Med. 2007, 101, 1412–1418. [Google Scholar]
- Binazzi, B.; Bianchi, R.; Romagnoli, I.; Lanini, B.; Stendardi, L.; Gigliotti, F.; Scano, G. Chest wall kinematics and Hoover's sign. Respir. Physiol. Neurobiol. 2008, 160, 325–333. [Google Scholar]
- Sharp, J.T. The chest wall and respiratory muscles in airflow limitation. In Thorax:Vital pump. Lung Biology in Healthy and Disease Series; Macklem, P.T., Roussos, C., Lenfant, C., Eds.; Marcel Dekker Inc.: NewYork, NY, USA, 1985; Part C; pp. 1155–1202. [Google Scholar]
- Hoover, C.F. The diagnostic significance of inspiratory movements of the costal margins. Am. J. Med. Sci. 1920, 159, 633–646. [Google Scholar]
- Jubran, A.; Tobin, M.J. The effect of hyperinflation on rib cage-abdominal motion. Am. Rev. Respir. 1992, 146, 1378–1382. [Google Scholar]
- Briscoe, W.A.; Dubois, A.B. The relationship between airway resistance, airway conductance and lung volume in subjects of different age and body size. J. Clin. Invest. 1958, 37, 1279–1285. [Google Scholar]
- Tobin, M.J.; Perez, W.; Guenther, S.M.; Lodato, R.F.; Dantzker, D.R. Does rib-cage abdominal paradox signify respiratory muscle fatigue? J. Appl. Physiol. 1987, 63, 851–860. [Google Scholar]
- Chihara, K.; Kenyon, C.M.; Macklem, P.T. Human rib cage distortability. J. Appl. Physiol. 1996, 81, 437–447. [Google Scholar]
- Gilmartin, J.J.; Gibson, G.J. Mechanisms of paradoxical rib cage motion in patients with chronic pulmonary disease. Am. Rev. Respir. Dis. 1986, 134, 683–687. [Google Scholar]
- Gorman, R.B.; McKenzie, D.K.; Pride, N.B.; Tolman, J.F.; Gandevia, S.C. Diaphragm length during tidal breathing in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2002, 166, 1461–1469. [Google Scholar]
- Scano, G.; Stendardi, L.; Grazzini, M. Understanding dyspnoea by its language. Eur. Respir. J. 2005, 25, 380–385. [Google Scholar]
- Leblanc, P.; Summers, E.; Inman, M.D.; Jones, N.L.; Campbell, E.J.; Killian, K.J. Inspiratory muscles during exercise: a problem of supply and demand. J. Appl. Physiol. 1988, 64, 2482–2489. [Google Scholar]
- Gorini, M.; Misuri, G.; Corrado, A.; Duranti, R.; Iandelli, I.; De Paola, E.; Scano, G. Breathing pattern and carbon dioxide retention in severe chronic obstructive pulmonary disease. Thorax 1996, 51, 677–683. [Google Scholar]
- Calverley, P.M. Dynamic hyperinflation: is it worth measuring? Proc. Am. Thorac. Soc. 2006, 3, 239–244. [Google Scholar]
- Hamilton, A.L.; Killian, K.J.; Summers, E.; Jones, N.L. Muscle strength, symptom intensity, and exercise capacity in patients with cardiorespiratory disorders. Am. J. Respir. Crit. Care Med. 1995, 152, 2021–2031. [Google Scholar]
- Hamilton, A.L.; Killian, K.J.; Summers, E.; Jones, N.L. Symptom intensity and subjective limitation to exercise in patients with cardiorespiratory disorders. Chest 1996, 110, 1255–1263. [Google Scholar]
- Maltais, F.; LeBlanc, P.; Simard, C.; Jobin, J.; Bérubé, C.; Bruneau, J.; Carrier, L.; Belleau, R. Skeletal muscle adaptation to endurance training in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1996, 154, 442–447. [Google Scholar]
- Aliverti, A.; Macklem, P.T. How and why exercise is impaired in COPD. Respiration 2001, 68, 229–239. [Google Scholar]
- Aliverti, A.; Stevenson, N.; Dellacà, R.L.; Lo Mauro, A.; Pedotti, A.; Calverley, P.M. Regional chest wall volumes during exercise in chronic obstructive pulmonary disease. Thorax 2004, 59, 210–216. [Google Scholar]
- De Troyer, A.; Sampson, M.; Sigrist, S.; Macklem, P.T. The diaphragm: two muscles. Science 1981, 213, 237–238. [Google Scholar]
- De Troyer, A.; Zegerdes De Beyl, D.; Thirion, M. Function of respiratory muscles in acute hemiplegia. Am. Rev. Respir. Dis. 1981, 123, 631–632. [Google Scholar]
- Cohen, E; Mier, A.; Heywood, P.; Murphy, K.; Boultbee, J.; Guz, A. Diaphragmatic movement in hemiplegic patients measured by ultrasonography. Thorax 1994, 49, 890–895. [Google Scholar]
- Fluck, D.C. Chest movements in hemiplegia. Clin. Sci. 1966, 31, 383–388. [Google Scholar]
- Klassen, A.C.; Heaney, L.M.; Lee, M.C.; Kronenberg, R.S. Altered cerebral inhibition of respiratory and cardiac responses to hypercapnia in acute stroke. Neurology 1980, 30, 951–955. [Google Scholar]
- Lanini, B.; Bianchi, R.; Romagnoli, I.; Coli, C.; Binazzi, B.; Gigliotti, F.; Pizzi, A.; Grippo, A.; Scano, G. Chest wall kinematics in patients with hemiplegia. Am. J. Respir. Crit. Care Med. 2003, 168, 109–113. [Google Scholar]
- Aliverti, A.; Molteni, F.; Carnevali, P.; Pedotti, A.; Bach, J.R.; Lissoni, A. Thoracoabdominal kinematics in Duchenne muscular dystrophy. Eur. J. Med. Rehabil. 1997, 7, 8–13. [Google Scholar]
- Bach, J.R. Mechanical insufflation-exsufflation: comparison of peak expiratory flows with manually assisted and unassisted coughing techniques. Chest 1993, 104, 1553–1562. [Google Scholar]
- Bach, J.R.; Ishikawa, I.; Kim, H. Prevention of pulmonary morbidity for patients with Duchenne muscular dystrophy. Chest 1997, 112, 1024–1028. [Google Scholar]
- De Troyer, A.; Estenne, M. Respiratory system in neuromuscular disorders. In The Thorax; Roussos, C., Ed.; Marcel Dekker: New York, NY, USA, 1995; Part C; pp. 2177–2212. [Google Scholar]
- Lanini, B.; Masolini, M.; Bianchi, R.; Binazzi, B.; Romagnoli, I.; Gigliotti, F.; Scano, G. Chest wall kinematics during voluntary cough in neuromuscular patients. Respir. Physiol. Neurobiol. 2008, 161, 62–68. [Google Scholar]
- Gattinoni, L.; Pelosi, P.; Suter, M.; Pedoto, A.; Vercesi, P.; Lissoni, A. Acute respiratory distress syndrome caused by pulmonary and extrapulmonary disease: different syndromes? Am. J. Respir. Crit. Care Med 1998, 158, 3–11. [Google Scholar]
- Aliverti, A.; Dellacá, R.; Pelosi, P.; Chiumello, D.; Pedotti, A.; Gattinoni, L. Optoelectronic plethysmography in intensive care patients. Am. J. Respir. Crit. Care Med. 2000, 161, 1546–1552. [Google Scholar]
- Aliverti, A.; Carlesso, E.; Dellacà, R.; Pelosi, P.; Chiumello, D.; Pedotti, A.; Gattinoni, L. Chest wall mechanics during pressure support ventilation. Crit. Care 2006, 10, R54. [Google Scholar]


© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the CreativeCommons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Romagnoli, I.; Lanini, B.; Binazzi, B.; Bianchi, R.; Coli, C.; Stendardi, L.; Gigliotti, F.; Scano, G. Optoelectronic Plethysmography has Improved our Knowledge of Respiratory Physiology and Pathophysiology. Sensors 2008, 8, 7951-7972. https://doi.org/10.3390/s8127951
Romagnoli I, Lanini B, Binazzi B, Bianchi R, Coli C, Stendardi L, Gigliotti F, Scano G. Optoelectronic Plethysmography has Improved our Knowledge of Respiratory Physiology and Pathophysiology. Sensors. 2008; 8(12):7951-7972. https://doi.org/10.3390/s8127951
Chicago/Turabian StyleRomagnoli, Isabella, Barbara Lanini, Barbara Binazzi, Roberto Bianchi, Claudia Coli, Loredana Stendardi, Francesco Gigliotti, and Giorgio Scano. 2008. "Optoelectronic Plethysmography has Improved our Knowledge of Respiratory Physiology and Pathophysiology" Sensors 8, no. 12: 7951-7972. https://doi.org/10.3390/s8127951
APA StyleRomagnoli, I., Lanini, B., Binazzi, B., Bianchi, R., Coli, C., Stendardi, L., Gigliotti, F., & Scano, G. (2008). Optoelectronic Plethysmography has Improved our Knowledge of Respiratory Physiology and Pathophysiology. Sensors, 8(12), 7951-7972. https://doi.org/10.3390/s8127951