Assessment of Organophosphate and Carbamate Pesticide Residues in Cigarette Tobacco with a Novel Cell Biosensor


Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Sensor fabrication from N2a cells
2.3. Sample preparation
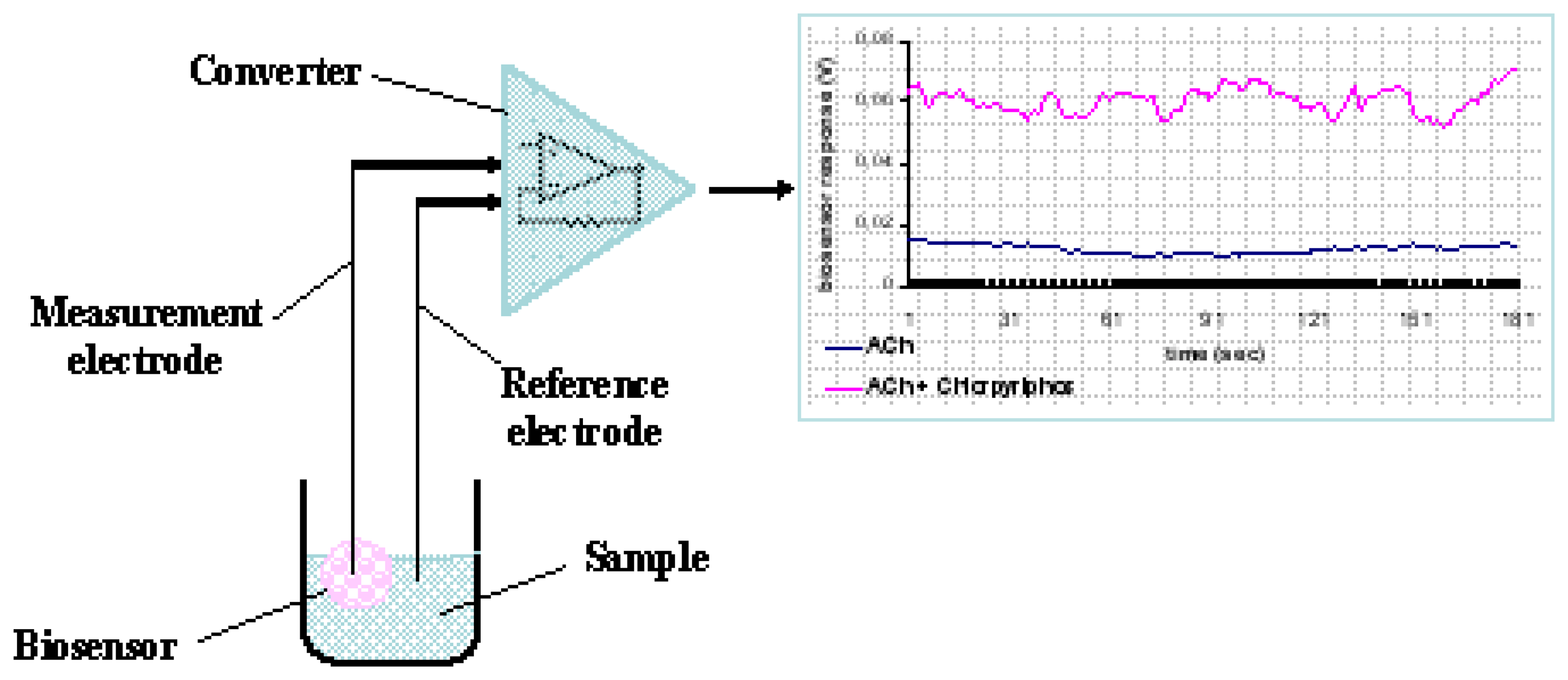
2.4. Assay principle
2.5. Assay procedure
2.6. Conventional sample analysis
2.7. Acetylcholinesterase inhibition assay
2.8. Assay of pesticide-cell interaction
2.9. Experimental design
3. Results
3.1. Response of the sensor to standard pesticide solutions
3.2. Residue profile by conventional analysis of tobacco leaf samples
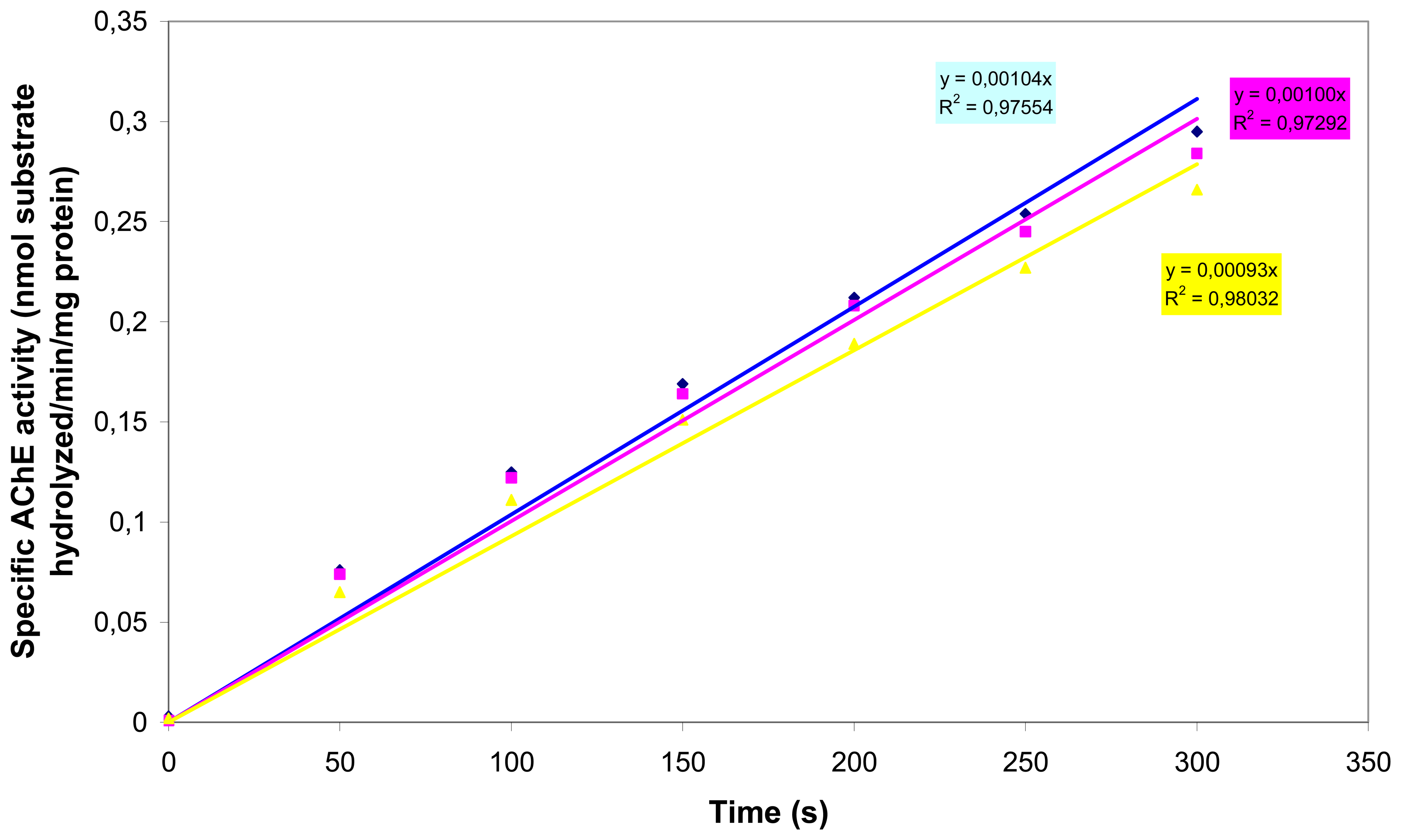
3.3. Acetylcholinesterase inhibition assay
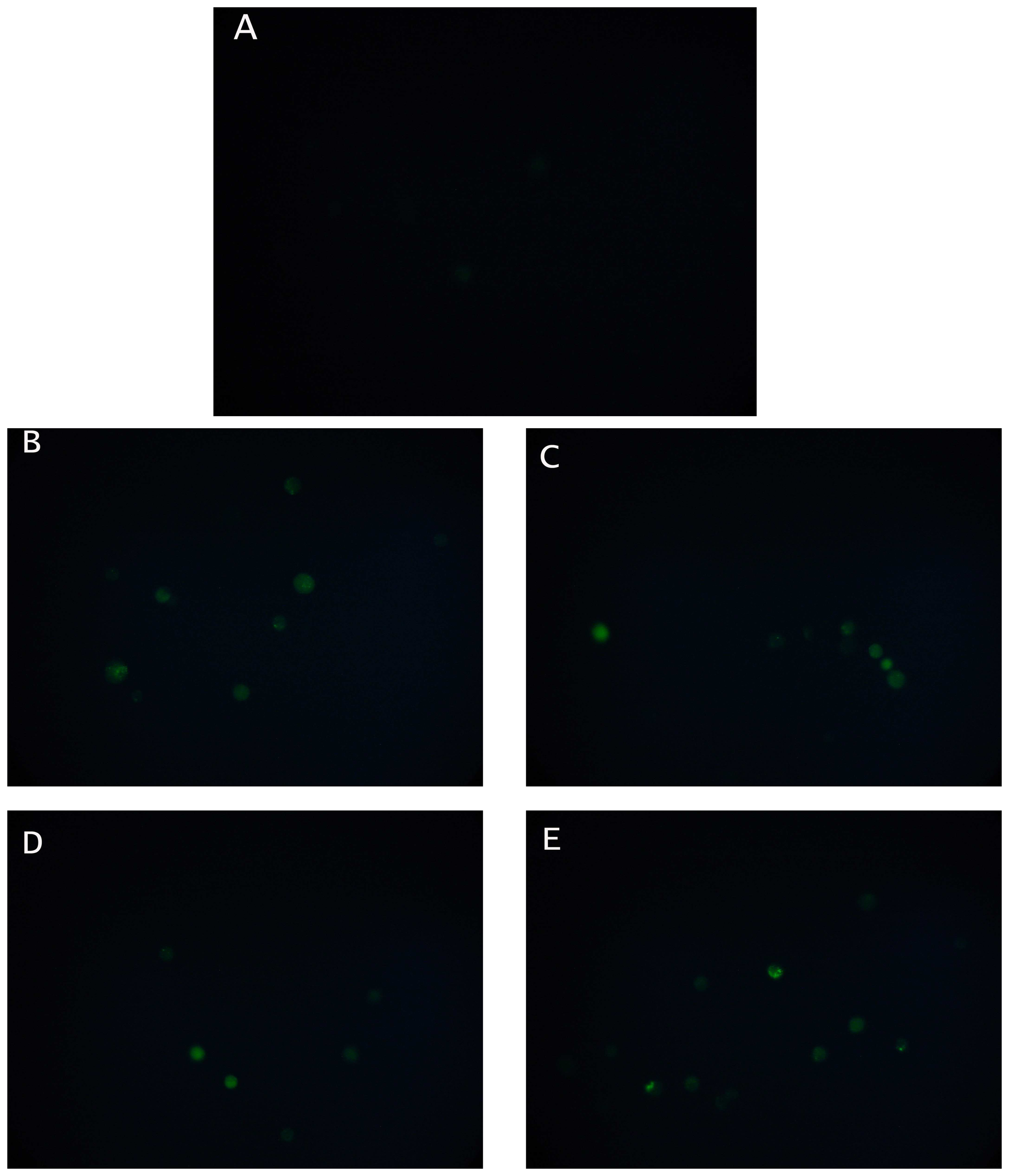
3.4. Effect of pesticides on calcium uptake by N2a cells
4. Discussion
- Ultra-high speed: an assay can be completed in less than 3 min.
- High sensitivity, with the ability to detect pesticides at concentrations as low as 1 ppb (for carbaryl).
- High reproducibility.
- Low cost, not exceeding 30-50 € per assay batch.
- The ability to monitor at real-time conditions.
- Finally, the ability to detect toxicity with complex, “real world” mixtures of toxicants and interfering compounds.
Acknowledgments
References
- Bachmann, T.T.; Leca, B.; Villate, F.; Marty, J.L.; Fournier, D.; Schmid, R.D. Improved multianalyte detection of organophosphates and carbamates with disposable multielectrode biosensors using recombinant mutants of Drosophila acetylcholinesterase and artificial neural networks. Biosens. Bioelectron. 2000, 15, 193–201. [Google Scholar]
- Nunes, G.S.; Montesino, T.; Marques, P.B.O.; Fournier, D.; Marty, J.L. Acetylcholine enzyme sensor for determining metamidophos insecticide. Evaluation of some genetically modified acetylcholinesterases from Drosophila melanogaster. Anal. Chim. Acta 2001, 434, 1–8. [Google Scholar]
- Drossopoulos, J.B.; Bouranis, D.L.; Kintzios, S.; Aivalakis, G.; Karides, J.; Chorianopoulou, S.N. Distribution profiles of selected macro-nutrients in oriental field grown tobacco plants as affected by nitrogen fertilization. J.Plant Nutrition 1998, 21, 1391–1406. [Google Scholar]
- Haib, J.; Hofer, I.; Renaud, J.-M. Analysis of multiple pesticide residues in tobacco using pressurized liquid extraction, automated solid-phase extraction clean-up and gas chromatography-tandem mass spectrometry. J. Chromatogr. A 2003, 1020, 173–187. [Google Scholar]
- Sherma, J. Review of advances in the thin layer chromatography of pesticides: 2004-2006. J Environ Sci Health B 2007, 42, 429–40. [Google Scholar]
- Kintzios, S. Cell-based sensors in clinical chemistry. Mini Rev. Clin. Chem. 2007, 7, 1019–1026. [Google Scholar]
- Cairns, J., Jr.; Mount, D.I. Aquatic toxicology. Environ. Sci.Technol. 1990, 24, 154–161. [Google Scholar]
- Daunert, S.; Barrett, G.; Feliciano, J. S.; Shetty, R.S.; Shrestha, S.; Smith-Spencer, W. Genetically engineered whole-cell sensing systems: coupling biological recognition with reporter genes. Chem. Rev. 2000, 100, 2705–2738. [Google Scholar]
- Kintzios, S.; Bem, F.; Mangana, O.; Nomikou, K.; Markoulatos, P.; Alexandropoulos, N.; Fasseas, C.; Arakelyan, V.; Petrou, A-L.; Soukouli, K.; Moschopoulou, G.; Yialouris, C.; Simonian, A. Study on the mechanism of Bioelectric Recognition Assay: evidence for immobilized cell membrane interactions with viral fragments. Biosens. Bioelectron. 2004, 20, 906–916. [Google Scholar]
- Kaiser, K.L.E.; Palabrica, V.S. Photobacterium phosphoreum Toxicity Data Index. Water Pollut. Res. J. Can. 1991, 26, 361–432. [Google Scholar]
- Riska, P.F.; Su, Y.; Bardarov, S.; Freundlich, L.; Sarkis, G.; Hatfull, G.; Carrière, C.; Kumar, V.; Chan, J.; Jacobs, W.R., Jr. Rapid Film-Based Determination of Antibiotic Susceptibilities of Mycobacterium tuberculosis Strains by Using a Luciferase Reporter Phage and the Bronx Box. J. Clin. Microbiol. 1999, 37, 1144–1449. [Google Scholar]
- Choi, S.H.; Gu, M.B. A portable toxicity biosensor using freeze-dried recombinant bioluminescent bacteria. Biosens. Bioelectron. 2002, 17, 433–440. [Google Scholar]
- Gu, M.B.; Min, J.; Kim, E.J. Toxicity monitoring and classification of endocrine disrupting chemicals (EDCs) using recombinant bioluminescent bacteria. Chemosphere 2002, 46, 289–294. [Google Scholar]
- Kim, B.C.; Park, K.S.; Kim, S.D.; Gu, M.B. Evaluation of a high throughput toxicity biosensor and comparison with a Daphnia magna bioassay. Biosens. Biolectron. 2003, 18, 821–826. [Google Scholar]
- Knight, A.W.; Keenan, P.O.; Billington, N.; Cahill, P.A.; Dalrymple, I.M.; Hawkyard, C.J.; Stratton-Campbell, D.; Walmsley, R.M. Clear and present danger? The use of a yeast biosensor to monitor changes in the toxicity of industrial effluents subjected to oxidative colour removal treatments. J. Environ. Monit. 2007, 9, 1394–1401. [Google Scholar]
- Slavik, J. Fluorescent probes in cellular and molecular biology; CRC Press: Boca Raton, 1993. [Google Scholar]
- Peechatinikov, V.A.; Rizvanov, F.F.; Pletnev, V.V. Permeability of sarcoplasma reticulum membrane for monovalent cations. Stud. Biophys. 1983, 28, 669–673. [Google Scholar]
- Eddy, A. Use of carbocyanine dyes to assay membrane potential of mouse ascites tumor cells. Methods in Enzymol 1989, 172, 95–101. [Google Scholar]
- Dixit, R.; Cyr, R. Cell damage and reactive oxygen species production induced by fluorescence microscopy: effect on mitosis and guidelines for non-invasive fluorescence microscopy. Plant J 2003, 36, 280–290. [Google Scholar]
- Kintzios, S.; Pistola, E.; Panagiotopoulos, P.; Bomsel, M.; Alexandropoulos, N.; Bem, F.; Varveri, C.; Ekonomou, G.; Biselis, J.; Levin, R. Bioelectric Recognition Assay (BERA). Biosens. Bioelectron. 2001, 16, 325–336. [Google Scholar]
- Kintzios, S.; Pistola, E.; Konstas, J.; Bem, F.; Matakiadis, T.; Alexandropoulos, N.; Biselis, J.; Levin, R. Application of the Bioelectric recognition assay (BERA) for the detection of human and plant viruses: definition of operational parameters. Biosens. Bioelectron. 2001, 16, 467–480. [Google Scholar]
- Hoang, B.X.; Graeme Shaw, D.; Pham, P.; Levine, S.A. Neuro-bioenergetic concepts in cancer prevention and treatment. Medical Hypotheses 2007, 68, 832–843. [Google Scholar]
- Anonymous. International Standard (ISO) 4389: Tobacco and tobacco products– Determination of organochlorine pesticide residues– gas chromatographic method; ISO: Geneva.
- Djordjevic, M.V.; Fan, J.; Hoffmann, D. Assessment of chlorinated pesticide residues in cigarette tobacco based on supercritical fluid extraction and GC-ECD. Carcinogenesis 1995, 16, 2627–2632. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–98. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Whelan, R.J.; Zare, R.N. Single-cell immunosensors for protein detection. Biosens. Bioelectron. 2003, 18, 1065–1072. [Google Scholar]
- Van der Lelie, D.; Regniers, L.; Borremans, B.; Provoost, A.; Verschaeve, L. The VITOTOX® test, an SOS bioluminescence Salmonella typhimurium test to measure genotoxicity kinetics. Mutat. Res. 1997, 389, 279–290. [Google Scholar]
- Anonymous. Commision recommendation concerning a coordinated Community monitoring programme for 2007 to ensure compliance with maxiumum levels of pesticides residues in and on cereals and certain other products of plant origin and national monitoring programmes for 2008. OJ L 2007, 96, 21–27. [Google Scholar]
- Anonymous. European Union Document : Quality control procedures for pesticide residue analysis. 2006, 10232, 1–30. [Google Scholar]
- Krnjevic, K. Early effects of hypoxia on brain cell function. Croat. Med. J. 1999, 40, 375–380. [Google Scholar]
- Moschopoulou, G.; Kintzios, S. Application of “membrane-engineering” to bioelectric recognition cell sensors for the detection of picomole concentrations of superoxide radical: a novel biosensor principle. Anal. Chimica Acta 2006, 573-574, 90–96. [Google Scholar]
- Frossyniotis, D.; Anthopoulos, Y.; Kintzios, S.; Perdikaris, A.; Yialouris, C.P. A multisensor fusion system for the detection of plant viruses by combining artificial neural networks. Lectures Comp. Sci. 2006, 4132, 401–409. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
| chlorpyrifos | 0,04 | 0,07 | 0,06 | 0,05 | 0,06 | 0,05 | 0,04 | 0,16 | 0,21 | 0,15 | 0,14 | 0,36 | 0,27 |
| Carbaryl | 0,23 | 0,42 | 0,21 | 0,48 | 0,25 | 0,2 | 0,97 | 1,4 | 1,1 | 0,64 | 0,94 | 2,26 | 1,6 |
© 2008 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Mavrikou, S.; Flampouri, K.; Moschopoulou, G.; Mangana, O.; Michaelides, A.; Kintzios, S. Assessment of Organophosphate and Carbamate Pesticide Residues in Cigarette Tobacco with a Novel Cell Biosensor. Sensors 2008, 8, 2818-2832. https://doi.org/10.3390/s8042818
Mavrikou S, Flampouri K, Moschopoulou G, Mangana O, Michaelides A, Kintzios S. Assessment of Organophosphate and Carbamate Pesticide Residues in Cigarette Tobacco with a Novel Cell Biosensor. Sensors. 2008; 8(4):2818-2832. https://doi.org/10.3390/s8042818
Chicago/Turabian StyleMavrikou, Sophie, Kelly Flampouri, Georgia Moschopoulou, Olga Mangana, Alexandros Michaelides, and Spiridon Kintzios. 2008. "Assessment of Organophosphate and Carbamate Pesticide Residues in Cigarette Tobacco with a Novel Cell Biosensor" Sensors 8, no. 4: 2818-2832. https://doi.org/10.3390/s8042818
APA StyleMavrikou, S., Flampouri, K., Moschopoulou, G., Mangana, O., Michaelides, A., & Kintzios, S. (2008). Assessment of Organophosphate and Carbamate Pesticide Residues in Cigarette Tobacco with a Novel Cell Biosensor. Sensors, 8(4), 2818-2832. https://doi.org/10.3390/s8042818