Laurate Biosensors Image Brain Neurotransmitters In Vivo: Can an Antihypertensive Medication Alter Psychostimulant Behavior?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Conventional Voltammetry and Microvoltammetry
- Involve measurement of current as a function of applied potential, wherein
- electroactive species undergo redox reactions at a characteristic redox potential.
- Formula:O + ne− ⇔ R, wherein, ne = number of electrons, O=oxidation, R= reduction.
- The current that is produced by a specific redox reaction is proportional to the concentration of neurochemicals, according to the Cottrell equation, described below.
- o=concentration of electroactive species oxidized.
- i= current at time, t
- n= number of electron transfers, eq/mol
- F= Faraday's constant, 96486 C/eq
- A= electrode area, cm 2
- C= concentration of o, mol/cm3
- D= Diffusion coefficient of o, cm2/s
2.2. Neuromolecular Imaging (NMI)
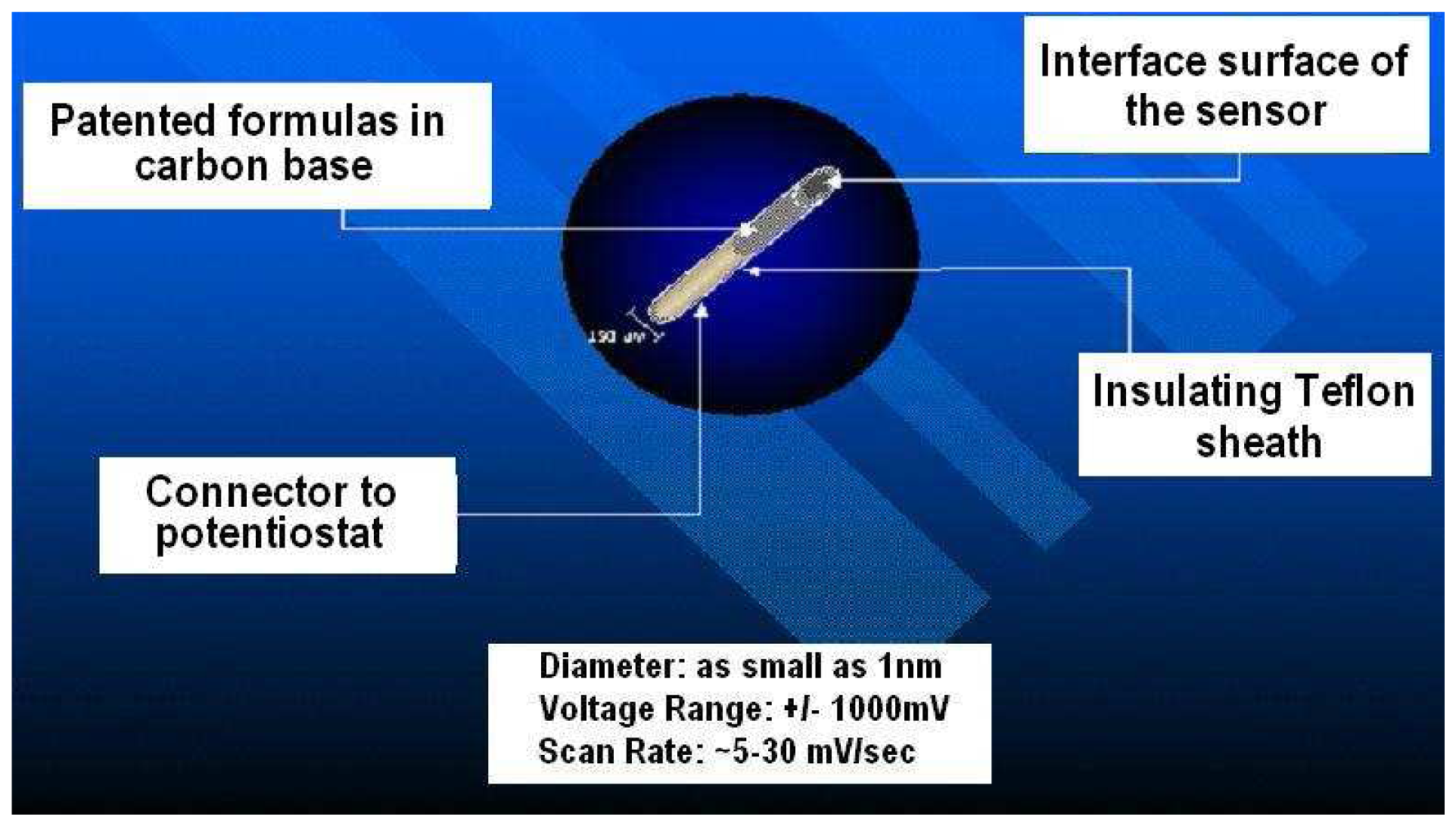
2.3. Manufacture of Laurate Biosensors
2.4. NMI Properties of Laurate Biosensor
2.5. Semiderivative Circuit
2.6. Calibration Procedures
2.7. Interpretation of NMI Signals
2.8. Animals and Surgical Procedures
2.9. Neuromolecular Imaging (NMI) On Line with Open Field Behavior
2.10. Study Design
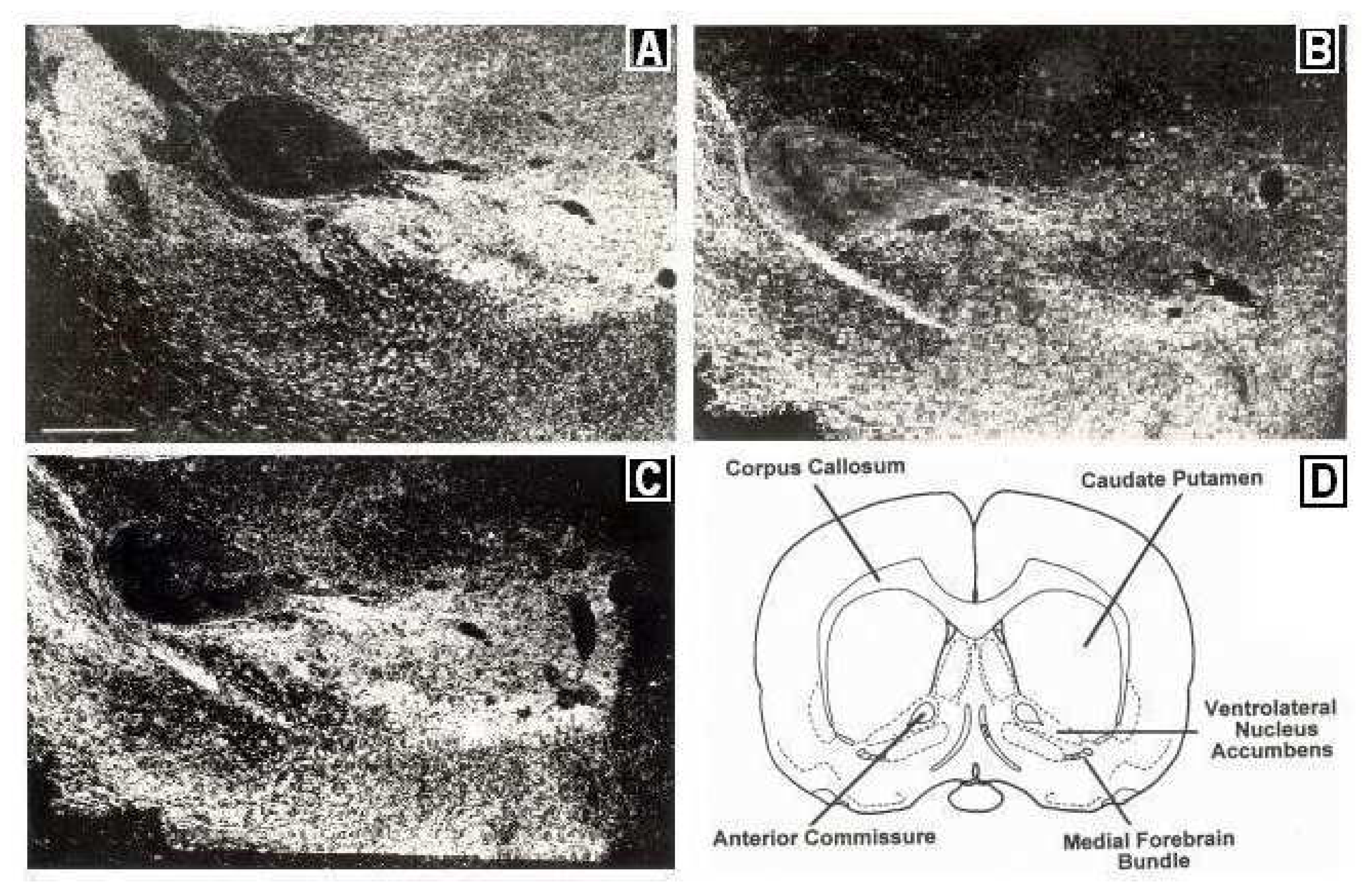
2.11. Confirmation of Biosensor Placement
2.12. Statistics
3. Results and Discussion
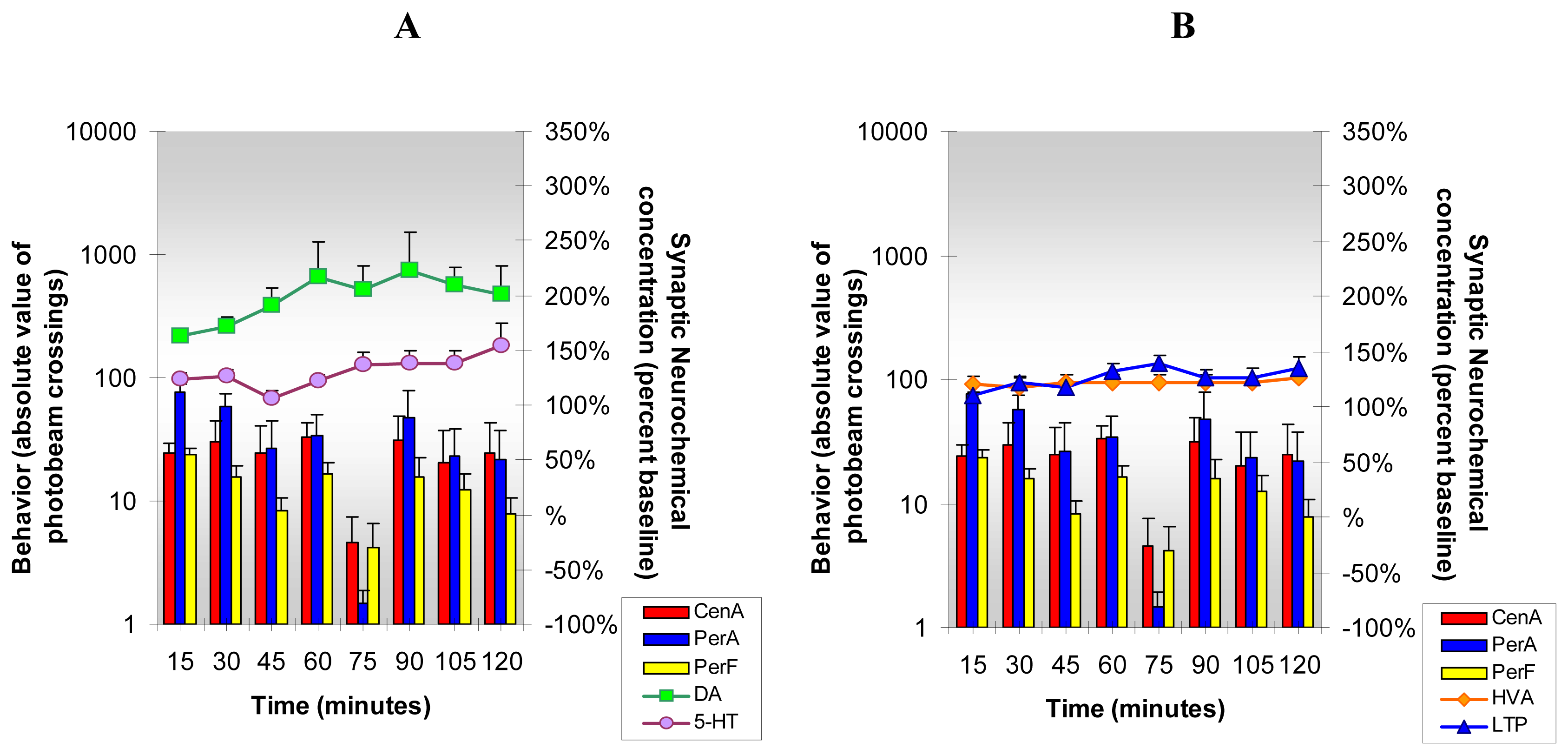
3.1. Changes in neurochemicals and behavior after cocaine
3.2. Changes in neurochemicals and behavior after caffeine
3.3. Changes in neurochemicals and behavior after cocaine and caffeine in combination
3.4. Changes in neurochemicals and behavior after ketanserin
3.5. Changes in neurochemicals and behavior after ketanserin and cocaine
3.6. Changes in neurochemicals and behavior after ketanserin and caffeine
3.7. Changes in neurochemicals and behavior after ketanserin, cocaine and caffeine
4. Conclusions
Acknowledgments
References
- Khan, A.S.; Michael, A.C. Invasive consequences of using microelectrodes and microdialysis probes in the brain. Trends in Anal.Chem. 2003, 22, 503–508. [Google Scholar]
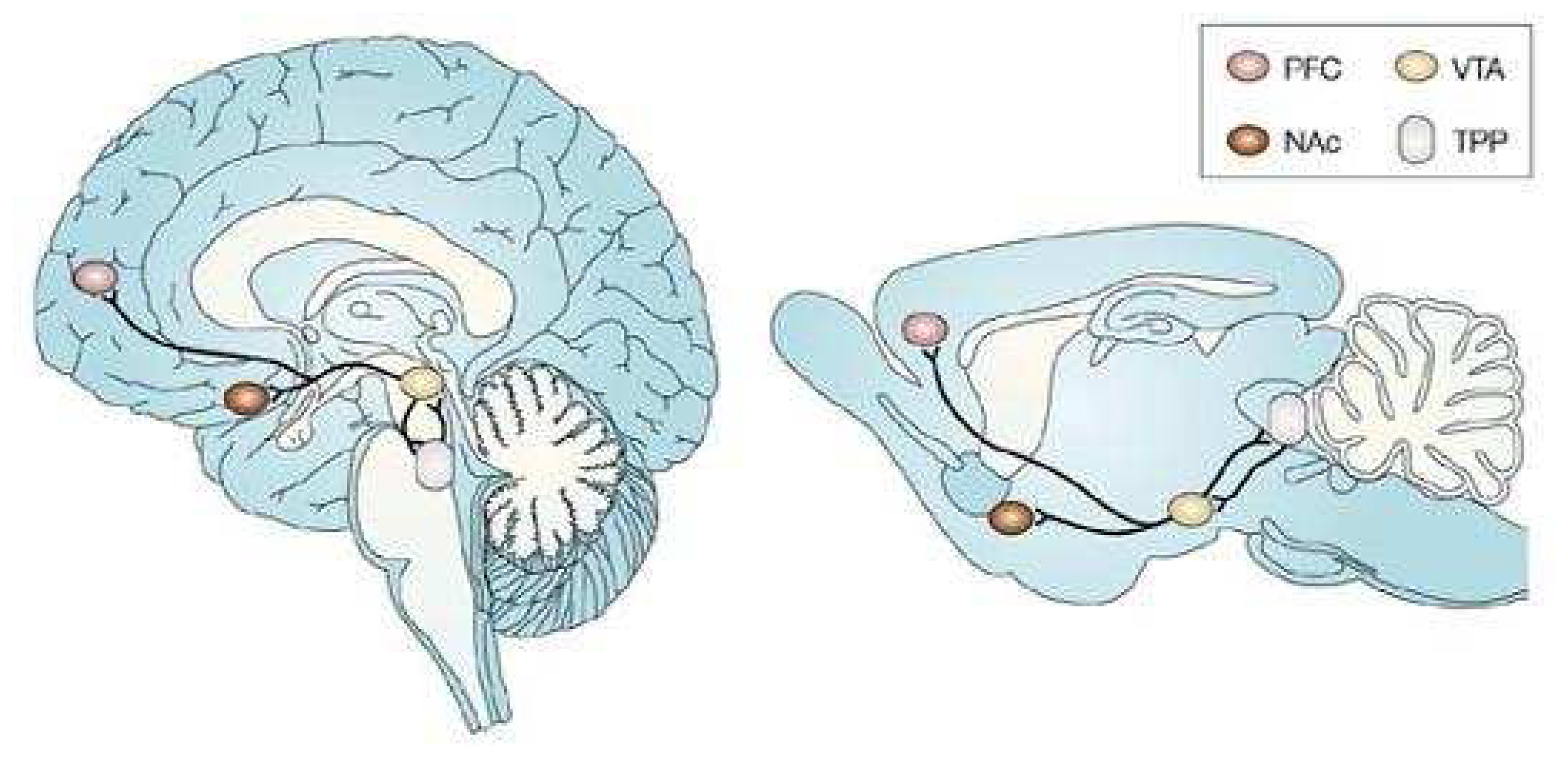
- Phelix, C.F.; Broderick, P.A. Light microscopic immunocytochemical evidence of converging serotonin and dopamine terminals in ventrolateral nucleus accumbens. Brain Res. Bull. 1995, 37, 37–40. [Google Scholar]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar]
- Koob, G.F.; Nestler, E.J. The neurobiology of drug addiction. J. Neuropsychiatry, Clinical Neuroscience 1997, 9, 482–497. [Google Scholar]
- Keiflin, R.; Isingrini, E.; Cador, M. Cocaine-induced reinstatement in rats: evidence for a critical role of cocaine stimulus properties. Psychopharmacology (Berl.) 2008. (Epub ahead of print). [Google Scholar]
- Filip, M.; Frankowska, M.; Zaniewska, M.; Przegalinski, E.; Muller, C.E.; Agnati, L.; Franco, R.; Roberts, D.C.; Fuxe, K. Involvement of adenosine A2A and dopamine receptors in the locomotor and sensitizing effects of cocaine. Brain Res. 2006, 1077, 67–80. [Google Scholar]
- Broderick, P.A.; Hope, O.; Okonji, C.; Rahni, D.N.; Zhou, Y. Clozapine and cocaine effects on dopamine and serotonin release in nucleus accumbens during psychostimulant behavior and withdrawal. Prog. Neuropsychopharmacol. Biol. Psychiat. 2004, 28, 157–171. [Google Scholar]
- Solinas, M.; Ferre, S.; You, Z.B.; Karcz-Kubicha, M.; Popoli, P.; Goldberg, S.R. Caffeine induces dopamine and glutamate release in the shell of the nucleus accumbens. J. Neuroscience 2002, 22, 6321–6324. [Google Scholar]
- Weinberg, B.A.; Bealer, B.K. The World of Caffeine; Routledge: New York and London, 2001; ISBN-0-415-92722-6. [Google Scholar]
- Li, W.; Dai, S.; An, J.; Li, P.; Chen, X.; Xiong, R.; Liu, P.; Wang, H.; Zhao, Y.; Zhu, M.; Liu, X.; Zhu, P.; Chen, J-F; Zhou, Y. Chronic but not acute treatment with caffeine attenuates traumatic brain injury in the mouse cortical impact model. Neuroscience 2008, 4, 1198–1207. [Google Scholar]
- Campbell, R.; Burchfield, G.; Hopes, M.; Mitchell, J. ABC ONLINE; Australian Broadcasting System, 1997. [Google Scholar]
- Salazar-Martinez, E.; Willet, W.C.; Ascherio, A.; Manson, J.E.; Leitzmann, M.F.; Stampfer, M.J.; Hu, F.B. Coffee consumption and risk for type 2 diabetes mellitus. Ann. Internal Med. 2004, 140, 1–8. [Google Scholar]
- Xu, K.; Xu, Y.; Brown-Jermyn, D.; Chen, J-F; Ascherio, A.; Diuzen, D.; Schwarzschild, M.A. Estrogen prevents neuroprotection by caffeine in the mouse 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson's Disease. Neurobiology of Disease. J. Neuroscience 2006, 26, 515–541. [Google Scholar]
- van der Starre, P.J.; Reneman, R.S. The alpha-adrenergic receptor blocking effect of ketanserin and the interaction between alpha-adrenergic and S2-serotonergic receptor blockade. J. Cardiovasc. Pharmacol. 1988, 11, S54–S61. [Google Scholar]
- Duffy, R.A.; Hunt, M.A.; Wamsley, J.K.; McQuade, R.D. In vivo autoradiography of [3H] SCH 39166 in rat brain: selective displacement by D1/D5 antagonists. J. Chem. Neuroanat. 2002, 19, 41–46. [Google Scholar]
- Lutje -Hulsik, D. Functional and Binding Data for Ketanserin Tartrate; Organon GPCRDB, 2002. [Google Scholar]
- Cazzola, M.; Matera, M.G.; Santangelo, G.; Assogna, G.; D'Amato, G.; Rossi, F.; Girbino, G. Effect of the selective 5-HT2 antagonist ketanserin on adenosine-induced bronchoconstriction in asthmatic patients. Immunopharmacology 1992, 23, 21–28. [Google Scholar]
- Hubbard, A.T.; Anson, F.C. Electroanalytical Chemistry; Bard, Alan J., Ed.; Marcel Dekker: New York, 1970; Volume V. [Google Scholar]
- Adams, R.N.; Marsden, C.A. New techniques in psychopharmacology. In Handbook of Psychopharmacology; Iversen, L.L, Snyder, S.H, Eds.; Plenum Press: New York, 1982. [Google Scholar]
- Kissinger, P.T.; Preddy, C.R.; Shoup, R.E.; Heineman, W.R. Fundamental concepts of analytical electochemistry. In Laboratory Techniques in Electroanalytical Chemistry; Kissinger, P.T., Heineman, W. R., Eds.; Marcel Dekker Inc.: New York, 1996; pp. 11–50. [Google Scholar]
- Broderick, P.A. Distinguishing in vitro electrochemical signatures for norepinephrine and dopamine. Neurosci Lett 1988, 95, 275–280. [Google Scholar]
- Broderick, P.A. Cathodic Electrochemical Current Arrangement with Telemetric Application. U.S. Patent # 4, 883,057, 1989(a). [Google Scholar]
- Broderick, P.A. Characterizing stearate probes in vitro for the electrochemical detection of dopamine and serotonin. Brain Res. 1989(b), 495, 115–121. [Google Scholar]
- Broderick, P.A. Microelectrodes and their use in cathodic electrochemical current arrangement with telemetric application. U.S. Patent # 5,433,710, 1995. [Google Scholar]
- Broderick, P.A. Microelectrodes and their use in an electrochemical arrangement with telemetric application. U.S. Patent # 5, 938, 903, 1999. [Google Scholar]
- Broderick, P.A.; Pacia, S.V. Identification, diagnosis, and treatment of neuropathologies, neurotoxicities, tumors and brain and spinal cord injuries using microelectrodes with microvoltammetry. U.S. Patent # 7,112,319, 2006. [Google Scholar]
- Garris, P.A.; Christensen, J.R.C.; Rebec, G.V.; Wightman, R.M. Real-time measurement of electrically evoked extracellular dopamine in the striatum of freely moving rats. J. Neurochem. 1997, 68, 152–161. [Google Scholar]
- David, D.J.; Zahniser, N.R.; Hoffer, B.J.; Gerhardt, G.A. In vivo electrochemical studies of dopamine clearance in subregions of rat nucleus accumbens: differential properties of the core and shell. Exp. Neurol. 1998, 153, 277–286. [Google Scholar]
- Kane, D.A.; O'Neill, R.D. Major differences in the behavior of carbon paste and carbon fibre electrodes in a protein-lipid matrix: Implications for voltammetry in vivo. Analyst 1998, 123, 2899–2903. [Google Scholar]
- Foucault, R.; Broderick, P.A.; Rahni, D.N.; Lombardi, J.R.; Birke, R.L. Neurotransmitter signatures: a correlation between Raman spectroscopy and the microvoltammetric BRODERICK PROBE®, NIH/NIGMS Symposium, New Orleans, LA; 2002.
- Broderick, P.A. Studies of oxidative stress mechanism using a morphine/ascorbate animal model and novel N-stearoyl cerebroside and laurate sensors. J. Neural Transm. 2008, 115, 7–17. [Google Scholar]
- Coury, L.A.; Huber, E.W.; Heineman, W.P. Applications of modified electrodes in the voltammetric determination of catecholamine neurotransmitters. Biotechnology 1989, 11, 1–37. [Google Scholar]
- Broderick, P.A.; Pacia, S.V.; Doyle, W.K.; Devinsky, O. Monoamine neurotransmitters in resected hippocampal subparcellations from neocortical and mesial temporal lobe epilepsy patients: in situ microvoltammetric studies. Brain Res. 2000, 878, 49–63. [Google Scholar]
- Broderick, P.A.; Pacia, S.V. Imaging white matter signals in epilepsy patients: A unique sensor technology. In Bioimaging in Neurodegeneration; Broderick, P.A., Rahni, D.N., Kolodny, E.H., Eds.; Humana Press Inc.: Totowa, NJ, 2005; pp. 199–206. [Google Scholar]
- Pellegrino, L.J.; Pellegrino, A.S.; Cushman, A.J. A Stereotaxic Atlas of the Rat Brain; Plenum Press: New York, 1979. [Google Scholar]
- Teitelbaum, P.; Pellis, S.M.; De Vietti, T.L. Disintegration into stereotypy induced by drugs or brain damage: a microdescriptive behavioral analysis. In Neurobiology of Stereotyped Behavior; Cooper, S.J., Dourish, S.T., Eds.; Oxford University Press: New York, 1990; pp. 169–199. [Google Scholar]
- Simon, P.; Dupuis, R.; Costentin, J. Thigmotaxis as an index of anxiety in mice. Influence of dopaminergic transmission. Behavioural Brain Research. 1994, 61, 59–64. [Google Scholar]
- Geyer, M.A.; Russo, P.V.; Masten, V.L. Multivariate assessment of locomotor behavior: pharmacological and behavioral analysis. Pharmacol. Biochem. Behav. 1986, 277–288. [Google Scholar]
- Mallo, T.; Alttoa, A.; Koiv, K.; Tonissaar, M.; Eller, M.; Harro, J. Rats with persistently low or high exploratory activity: behaviour in tests of anxiety and depression, and extracellular levels of dopamine. Behavioral Brain Res. 2007, 177, 269–281. [Google Scholar]
- Broderick, P.A.; Phelix, C.F.I. Serotonin (5-HT) within dopamine reward circuits signals open-field behavior. II. basis for 5-HT-DA interaction in cocaine dysfunctional behavior. Brain Res. Bull. 1997, 21, 227–260. [Google Scholar]
- Chin, J.; Sternin, O.; Fletcher, H.; Jenab, S.; Perrotti, L.I.; Quinones-Jenab, V. Sex differences in cocaine-induced behavioral sensitization. Cell. Mol. Biol. 2001, 47, 1089–1095. [Google Scholar]
- Kosten, T.R.; Kosten, T.A.; McDougle, C.J.; Hameedi, F.A.; McCance, F.F.; Rosen, M.I. Gender differences in response to intranasal cocaine administration in humans. Biological Psychiat. 1996, 39, 147–148. [Google Scholar]
- Nunes, J.; Broderick, P.A. Novel research translates to clinical cases of schizophrenia and cocaine psychosis. Neuropsychiatric Disease and Treatment. 2007, 3, 475–486. [Google Scholar]
- Broderick, P.A.; Kornak, E.P.; Eng, F.; Wechsler, R. Real time detection of acute (IP) cocaine-enhanced dopamine and serotonin release in ventrolateral nucleus accumbens of the behaving Norway rat. Pharmacol Biochem Behav. 1993, 46, 715–722. [Google Scholar]
- Carlsson, A.; Carlsson, M.L. Dopaminergic deficit hypothesis of schizophrenia: the path to discovery. Dialogues Clin. Neurosci. 2006, 8, 1370142. [Google Scholar]
- Bubar, M.J.; Cunningham, K.A. Serotonin 5-HT2A and 5-HT2C receptors as potential targets for modulation of psychostimulant use and dependence. Curr. Top. Med. Chem. 2006, 6, 1971–1985. [Google Scholar]
- Henry, L.K.; Blakely, R.D. Distinctions between dopamine transporter antagonists could be just around the bend. Mol. Pharmacol. 2008, 73, 616–618. [Google Scholar]
- Marcellino, D.; Roberts, D.C.S.; Navarro, G.; Filip, M.; Agnati, L.; Lluis, C.; Franco, R.; Fuxe, K. Increase in A2A receptors in the nucleus accumbens after extended cocaine self-administration and its disappearance after cocaine withdrawal. Brain Res. 2007, 1143, 208–220. [Google Scholar]
- Zelek-Molik, A.; Kreiner, G.; Roman, A.; Bielawski, A.; Nalepa, I. Effect of repeated cocaine administration on the mRNAs expression of main classes of G alpha proteins in the rat brain. Eur. Neuropsychopharmacol. 2006, 16 (Suppl. 1), S8–S9. [Google Scholar]
- Thomas, M.J.; Kalivas, P.W.; Shaham, Y. Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br. J. Pharmacol. 2008, 154, 327–342. [Google Scholar]
- Griffiths, R.R.; Mumford, G.K. Caffeine: A drug of abuse? Neuropsychopharmacology. The Fifth Generation of Progress. 2000. http://www.acnp.org/G4/GN401000165/CH161.html.
- Weerts, E.M.; Griffiths, R.R. The adenosine receptor antagonist CGS15943 reinstates cocaine-seeking behavior and maintains self-administration in baboons. Psychopharmacology (Berl.) 2003, 168, 155–163. [Google Scholar]
- Franco, R.; Casado, V.; Cortes, A.; Mallol, J.; Ciruela, F.; Ferre, S.; Lluis, C.; Canela, E.I. G-protein coupled receptor heteromers: function and ligand pharmacology. Br. J. Pharmacol. 2008, 153, 590–598. [Google Scholar]
- Ward, R.P.; Dorsa, D.M. Molecular and behavioral effects mediated by Gs-coupled adenosine A2a, but not serotonin 5-Ht4 or 5-Ht6 receptors following antipsychotic administration. Neurosci. 1999, 89, 927–938. [Google Scholar]
- Rashid, A.J.; O'Dowd, B.F.; Verma, V.; George, S.R. Neuronal Gq/11-coupled dopamine receptors: an uncharted role for dopamine. Trends in Pharmacol. Sci. 2007, 28, 551–555. [Google Scholar]
- Daly, J.W.; Fredholm, B.B. Caffeine-an atypical drug of dependence. Drug Alcohol Depend. 1998, 55, 199–206. [Google Scholar]
- Fredholm, B.B.; Battig, K.; Holmen, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev. 1999, 55, 88–133. [Google Scholar]
- Solinas, M.; Ferre, S.; Antoniou, K.; Quarta, D.; Justinova, Z.; Hockemeyer, J.; Pappas, L.A.; Segal, P.N.; Wertheim, C.; Muller, C.E.; Goldberg, S.R. Involvement of adenosine A1 receptors in the discriminative-stimulus effects of caffeine in rats. Psychopharmacology (Berl.) 2005, 179, 576–586. [Google Scholar]
- Ciruela, F.; Casado, V.; Rodrigues, R.J.; Lujan, R.; Burgueno, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; Cortes, A.; Canela, E.I.; Lopez-Gimenez, J.F.; Milligan, G.; Lluis, C.; Cunha, R.A.; Ferre, S.; Franco, R. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar]
- Ciruela, F.; Ferre, S.; Casado, V.; Cortes, A.; Cunha, R.A.; Lluis, C.; Franco, R. Heterodimeric adenosine receptors: a device to regulate neurotransmitter release. Cell.Mol.Life Sci. 2006, 63, 2427–2431. [Google Scholar]
- Capper-Loup, C.; Canales, J.J.; Kadaba, N.; Graybiel, A.M. Concurrent activation of dopamine D1and D2 receptors is required to evoke neural and behavioral phenotypes of cocaine sensitization. J.Neurosci. 2002, 6218–6227. [Google Scholar]
- Broderick, P.A.; Medicherla, R.; Zhou, F.; Forrester, L.W. Imaging accumbens monoamines with behavior: ketanserin, caffeine and cocaine interactions. In Society for Neuroscience; 2005; Wash. D.C., November 12-16. [Google Scholar]
- Horger, B.A.; Wellman, P.J.; Morien, A.; Davies, B.T.; Schenk, S. Caffeine exposure sensitizes rats to the reinforcing effects of cocaine. Neuroreport 1991, 2, 53–56. [Google Scholar]
- Schenk, S.; Worley, C.M.; McNamara, C.; Valadez, A. Acute and repeated exposure to caffeine: effects on reinstatement of extinguished cocaine-taking behavior in rats. Psychopharmacology 1996, 126, 17–23. [Google Scholar]
- Carroll, M.E.; Lac, S.T. Dietary additives and the acquisition of cocaine self-administration in rats. Psychopharmacology 1998, 137, 81–89. [Google Scholar]
- Broderick, P.A; Ho, H.; Huang, J.; Wat, K.; Shahidullah, J.; Murthy, V.; Wenning, L.; Buchheim, L. Caffeine may neuroprotect against cocaine-induced dopamine release in accumbens of female rats. In Society for Neuroscience; 2007; San Diego, CA, November 3-7. [Google Scholar]
- Nesbitt, J.; Raju, C.; Vidal, L.; Alimova, A.; Steiner, J.; Broderick, P.A.; Friedman, E. Sexual dimorphism studies of cocaine and caffeine in conditioned place preference: estrous cycle stage and open-field behaviors. In Society for Neuroscience; 2008; Wash, D.C., November 15-19. [Google Scholar]
- Schiffman, S.N.; Fisone, G.; Moresco, R.; Cunha, R.A.; Ferre, S. Adenosine A2A receptors and basal ganglia physiology. Prog. Neurobiol. 2007, 63, 277–292. [Google Scholar]
- Leysen, J.E.; Gommeren, W.; Schotte, A. Serotonin receptor subtypes: possible roles and implications in antipsychotic drug action. In Serotonergic Mechanisms in Antipsychotic Treatment; Kane, J.M., Moller, H.-J., Awouters, F., Eds.; Marcel-Dekker: New York, 1996; pp. 51–76. [Google Scholar]
- Gleerup, G.; Persson, B.; Hedner, T.; Winther, K. Serotonin-induced platelet aggregation predicts the antihypertensive response to serotonin receptor antagonists. Eur J Clin Pharmacol 1993, 44, 121–125. [Google Scholar]
- Mandhane, S.N.; Khisti, R.T.; Chopde, C.T. Effect of serotonergic agents on adenosine A2 receptor mediated catalepsy in rats. Psychopharmacology (Berl.) 1998, 137, 113–119. [Google Scholar]
- Shi, J.; Damjanoska, K.J.; Singh, R.K.; Carrasco, G.A.; Garcia, F.; Grippo, A.J.; Landry, M.; Sullivan, N.R.; Battaglia, G.; Muma, N.A. Agonist induced-phosphorylation of Galpha11 protein reduces coupling to 5-HT2A receptors. J. Pharmacol. Exp. Ther. 2007, 323, 248–256. [Google Scholar]
- Adlersberg, M.; Arango, V.; Hsiung, S.; Mann, J.J.; Underwood, M.D.; Liu, K.; Kassir, S.A.; Ruggiero, D.A.; Tamir, H. In vitro autoradiography of serotonin 5-HT(2A/2C) receptor-activated G protein: guanosine-5′-(gamma-[(35)S]thio)triphosphate binding in rat brain. J. Neurosci. Res. 2000, 61, 674–685. [Google Scholar]
- McMahon, L.R.; Cunningham, K.A. Antagonism of 5-hydroxytryptamine(2a) receptors attenuates the behavioral effects of cocaine in rats. J. Pharmacol. Exp.Ther. 2001, 297, 357–363. [Google Scholar]
- Broderick, P.A.; Olabisi, O.A.; Rahni, D.N.; Zhou, Y. Cocaine acts on accumbens monoamines and locomotor behavior via a 5-HT2A/2C receptor mechanism as shown by ketanserin: 24-h follow-up studies. Prog. Neuropsychopharmacol.Biol. Psychiat. 2004, 28, 547–557. [Google Scholar]
- Schindler, C. Cocaine and cardiovascular toxicity. Addict. Biol. 1996, 1, 31–47. [Google Scholar]
- Knuepfer, M.M. Muscarinic cholinergic and beta-adrenergic contribution to hindquarters vasodilation and cardiac responses to cocaine. J. Pharmacol. Exp. Ther. 2003, 306, 515–522. [Google Scholar]
- Miao, C.Y.; Yuan, W.J.; Su, D.F. Comparative study of sinoaortic denervation rats and spontaneously hypertensive rats. Am. J. Hypertension 2003, 16, 585–591. [Google Scholar]
- Kirouac, G.J.; Ganouly, P.K. Up-regulation of dopamine receptors in the brain of the spontaneously hypertensive rat: an autoradiographic study. Neuroscience 1993, 52, 135–141. [Google Scholar]
- Kirouac, G.J.; Ganouly, P.K. Cholecystokinin-induced release of dopamine in the nucleus accumbens of the spontaneously hypertensive rat. Brain Res. 1995, 689, 245–253. [Google Scholar]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Molina, P.E.; Logan, J.; Gatley, S.J.; Gifford, A.; Ding, Y.S.; Wong, C.; Pappas, N.R.; Zhu, W.; Swanson, J.M. Cardiovascular effects of methylphenidate in humans associated with increases of dopamine in brain and of epinephrine in plasma. Psychopharmacology (Berl.) 2003, 166, 264–270. [Google Scholar]
- Toda, S.; Alguacil, L.F.; Kalivas, P.W. Repeated cocaine administration changes the function and subcellular distribution of adenosine A1 receptor in the rat nucleus accumbens. J. Neurochem. 2003, 86, 1478–1484. [Google Scholar]
- Bowers, M.S.; McFarland, K.; Russell, W.L.; Peterson, Y.K.; Lapish, C.C.; Gregory, M.L.; Lanier, S.M.; Kalivas, P.W. Activator of G protein signalling 3 A gatekeeper of cocaine sensitization and drug seeking. Neuron 2004, 42, 269–281. [Google Scholar]
- McGinty, J.F.; Shi, X.D.; Schwendt, M.; Saylor, A.; Toda, S. Regulation of psychostimulant-induced signaling and gene expression in the striatum. J. Neurochem. 2008, 104, 1440–1449. [Google Scholar]
- Harvey, J.A.; Romano, A.G.; Gabriel, M.; Simansky, K.J.; Du, W.; Aloyo, V.J.; Friedman, E. Effects of prenatal exposure to cocaine on the developing brain: anatomical, chemical, physiological and behavioral consequences. Neurotox. Res. 2001, 3, 117–143. [Google Scholar]
- Svenningsson, P.; Nishi, A.; Fisone, G.; Girault, J.-A.; Nairn, A.C.; Greengard, P. DARPP-32: An integrator of neurotransmission. Ann. Rev. Pharmacol. Toxicol. 2004, 269–296. [Google Scholar]
- Broderick, P.A.; Li, Y-S.; Ho, H.; Wat, K.; Haile, M.M.; Bekker, A.Y.; Kolodny, E.H. Neuromolecular imaging of monoamine and peptide neurotransmitters in an animal model of Parkinson's disease: effects of the dopamine agonist, bromocriptine. In Society for Neuroscience; 2008; Wash. D.C., November 15-19. [Google Scholar]
- Haile, M.M.; Broderick, P.A.; Li, Y-S.; Quartermain, D.; Blanck, T.J.J; Bekker, A.Y. Nimodipine reverses the hypoxia-induced elevation of dopamine and serotonin in striatum of adult rats. In American Society of Anesthesiologists; 2008; Orlando, FL., October 18-22.
- Laviolette, S.R.; van der Kooy, D. The neurobiology of nicotine addiction: bridging the gap from molecules to behavior. Nature Reviews Neuroscience. 2004, pp. 55–65. http://www.nature.com/nrn/journal/v5/n1/fig_tab/nrn1298_F2.html.
- Swanson, L.W. The projections of the ventral tegmental area and adjacent regions: a combined retrograde tracer and immunofluourescence study in the rat. Brain Res. Bull. 1982, 9, 321–353. [Google Scholar]













© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Broderick, P.A.; Ho, H.; Wat, K.; Murthy, V. Laurate Biosensors Image Brain Neurotransmitters In Vivo: Can an Antihypertensive Medication Alter Psychostimulant Behavior? Sensors 2008, 8, 4033-4061. https://doi.org/10.3390/s8074033
Broderick PA, Ho H, Wat K, Murthy V. Laurate Biosensors Image Brain Neurotransmitters In Vivo: Can an Antihypertensive Medication Alter Psychostimulant Behavior? Sensors. 2008; 8(7):4033-4061. https://doi.org/10.3390/s8074033
Chicago/Turabian StyleBroderick, Patricia A., Helen Ho, Karyn Wat, and Vivek Murthy. 2008. "Laurate Biosensors Image Brain Neurotransmitters In Vivo: Can an Antihypertensive Medication Alter Psychostimulant Behavior?" Sensors 8, no. 7: 4033-4061. https://doi.org/10.3390/s8074033
APA StyleBroderick, P. A., Ho, H., Wat, K., & Murthy, V. (2008). Laurate Biosensors Image Brain Neurotransmitters In Vivo: Can an Antihypertensive Medication Alter Psychostimulant Behavior? Sensors, 8(7), 4033-4061. https://doi.org/10.3390/s8074033