Electrographic Changes Accompanying Recurrent Seizures under Ketogenic Diet Treatment




Abstract
:1. Introduction
2. Results
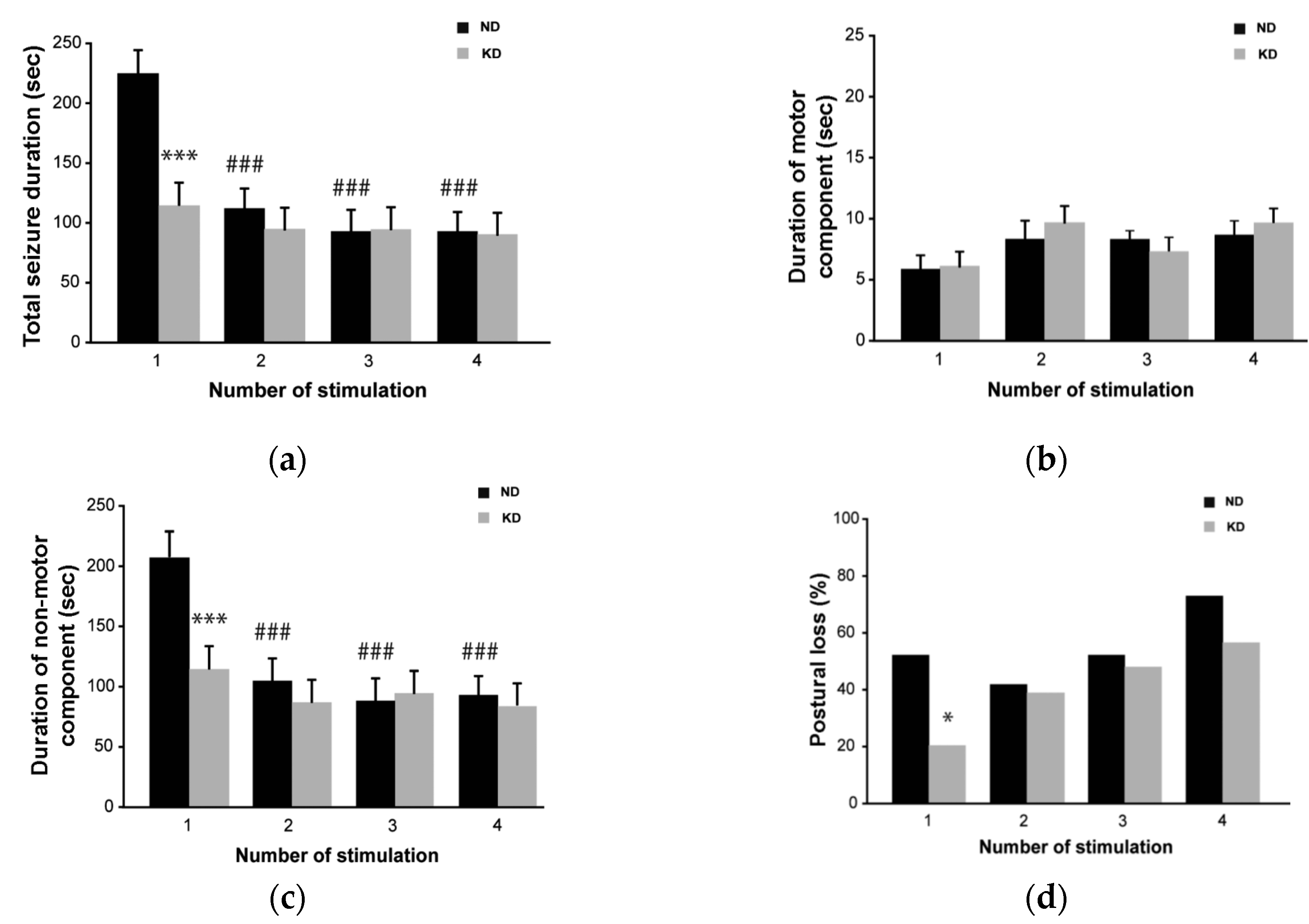
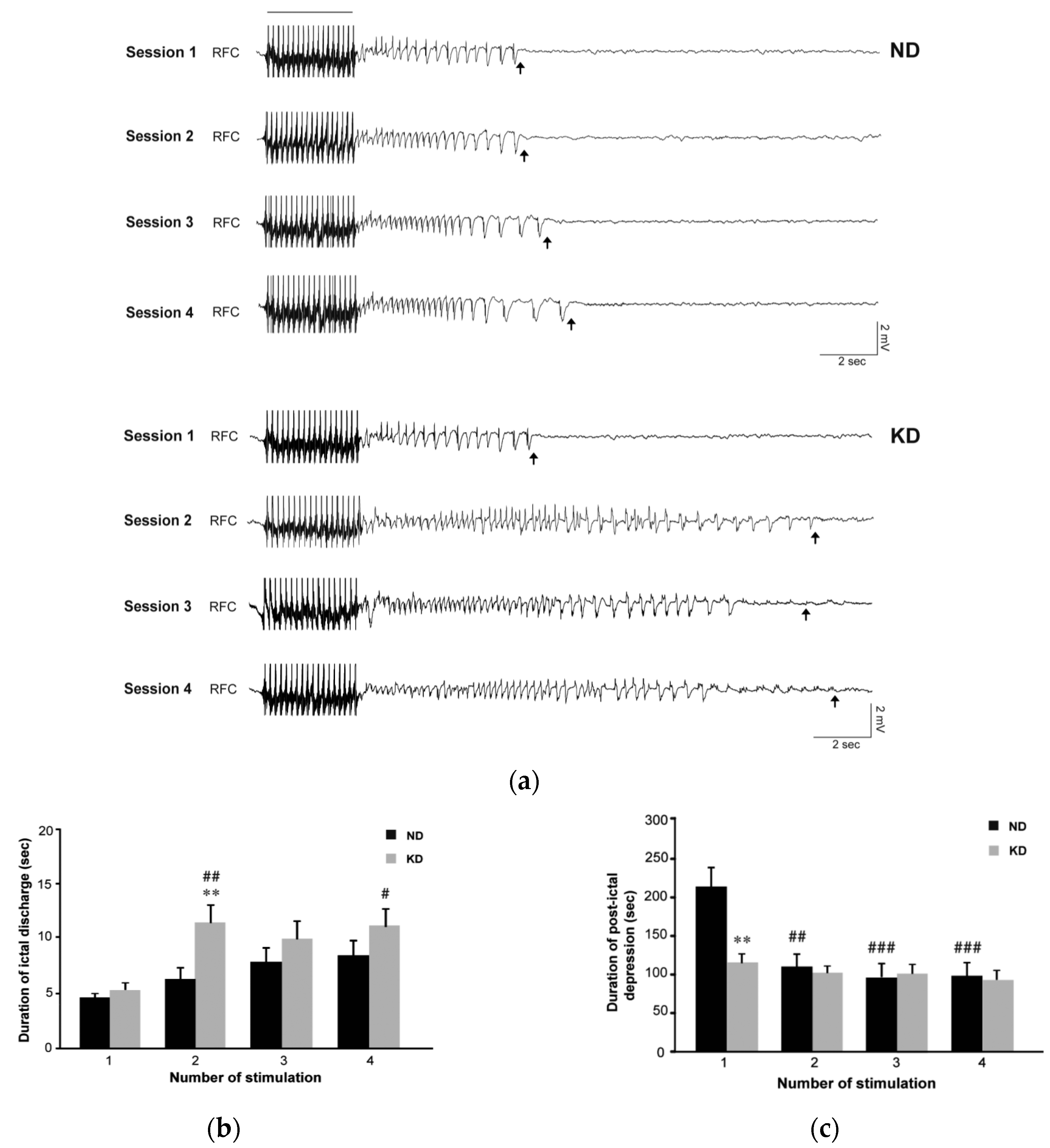
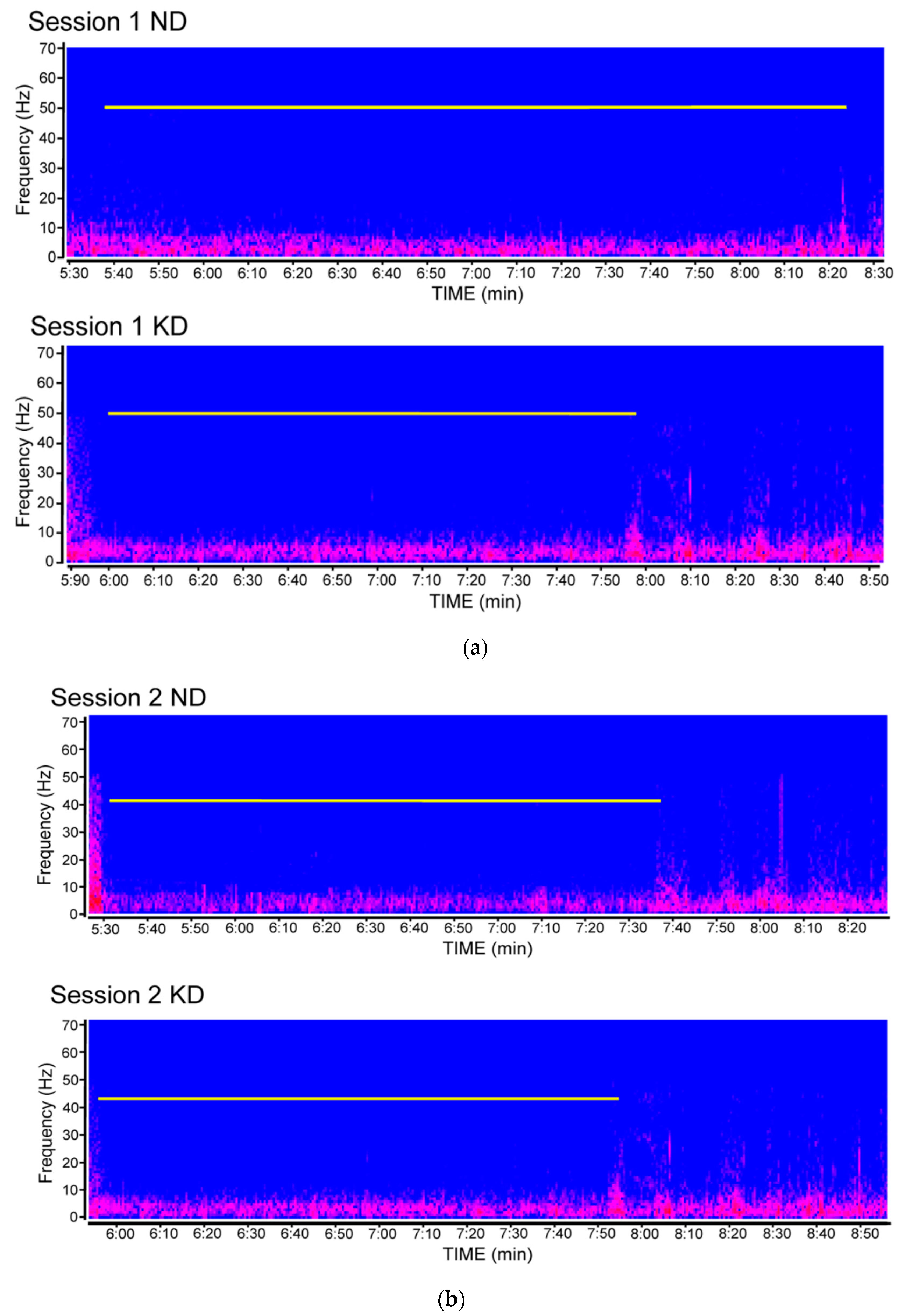
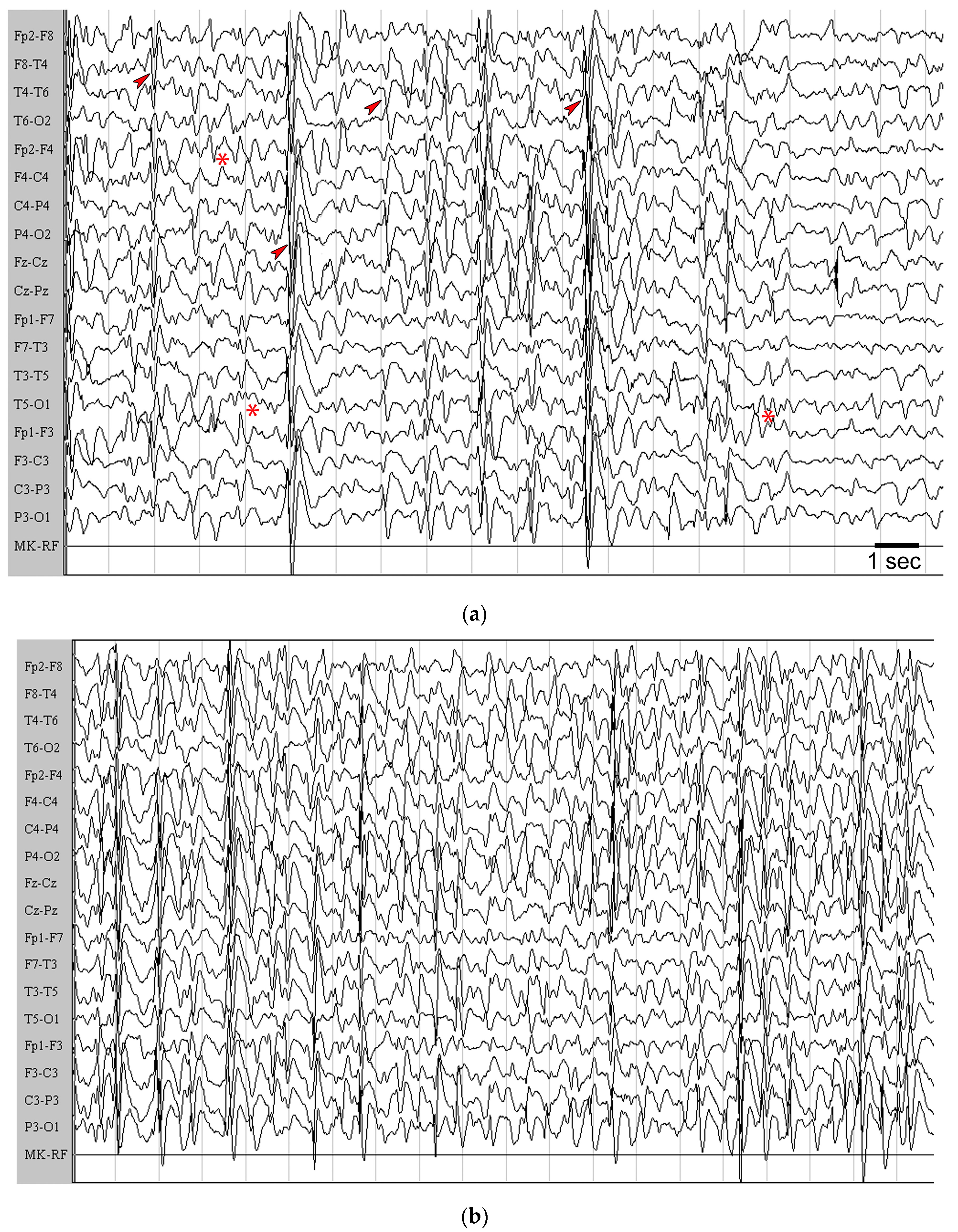
2.1. Seizure Worsening after an Initial Beneficial Effect of KD Administration in Mice
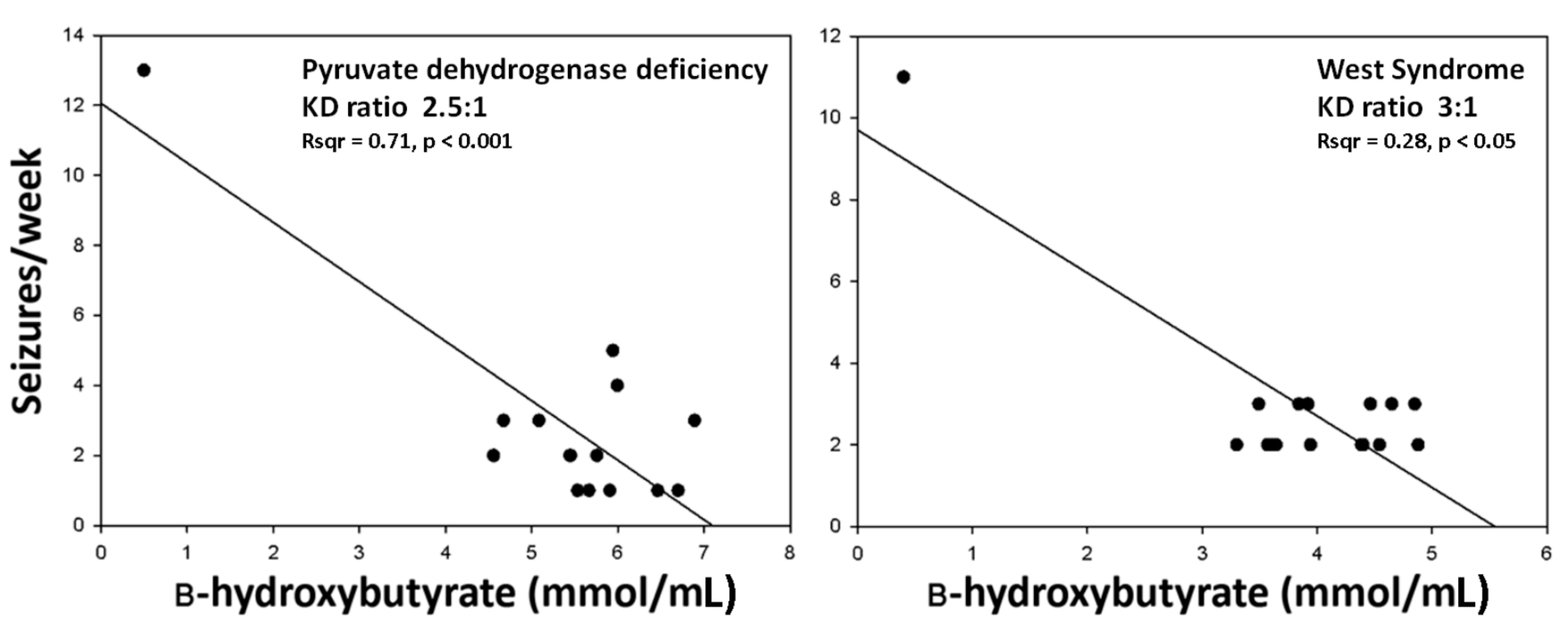
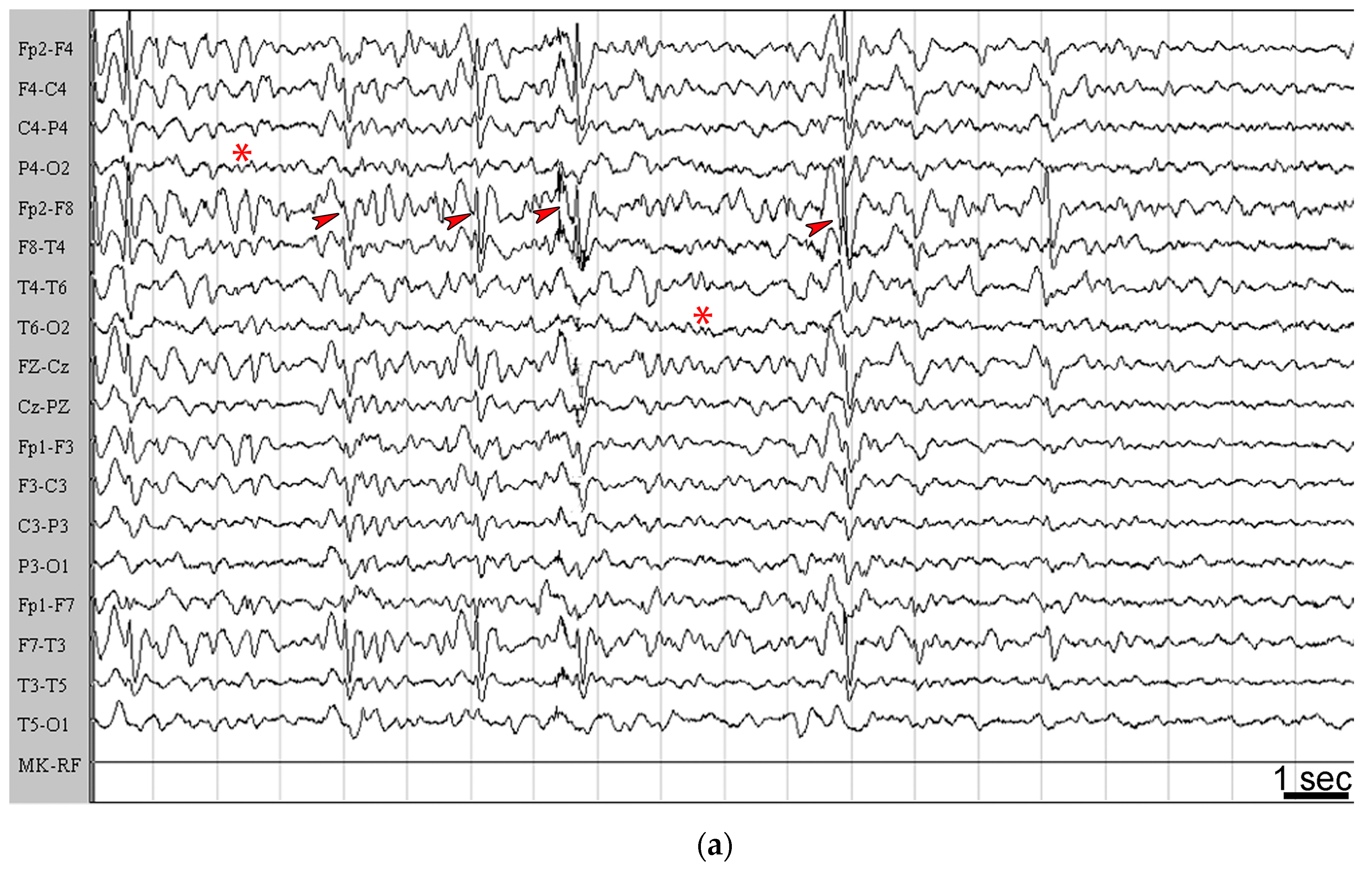
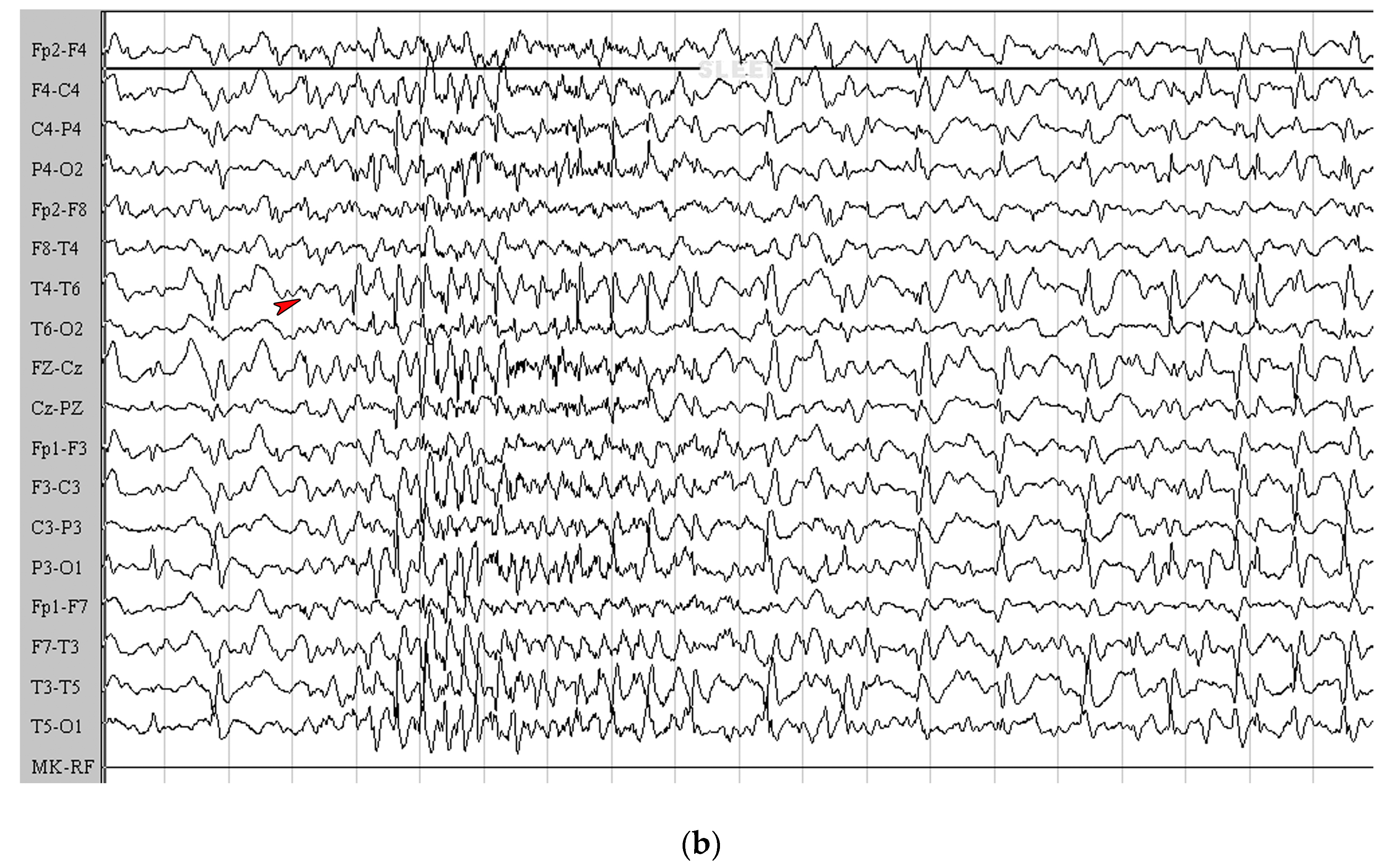
2.2. Seizures Worsened in Three Patients Maintained on the KD
3. Discussion
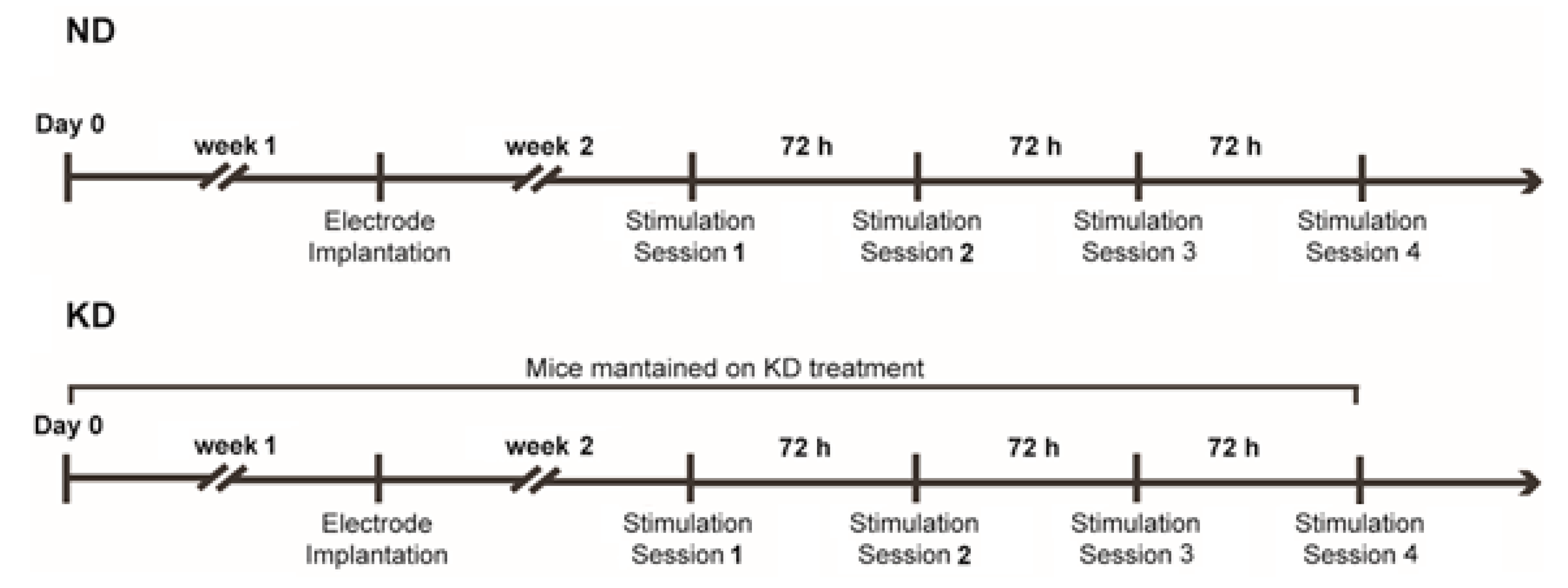
4. Materials and Methods
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, A.; Trevick, S. The epidemiology of global epilepsy. Neurol. Clin. 2016, 34, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.; Choi, H.; Schlesinger, M.; Rodemer, W.; Pollard, J.; Hesdorffer, D.C.; Hauser, W.A.; French, J. Increased mortality persists in an adult drug-resistant epilepsy prevalence cohort. J. Neurol. Neurosurg. Psychiatr. 2014, 85, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Giordano, C.; Marchiò, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Neal, E.G.; Chaffe, H.; Schwartz, R.H.; Lawson, M.S.; Edwards, N.; Fitzsimmons, G.; Whitney, A.; Cross, J.H. The ketogenic diet for the treatment of childhood epilepsy: A randomised controlled trial. Lancet Neurol. 2008, 7, 500–506. [Google Scholar] [CrossRef]
- Martin, K.; Jackson, C.F.; Levy, R.G.; Cooper, P.N. Ketogenic diet and other dietary treatments for epilepsy. Cochrane Database Syst. Rev. 2012, 3. [Google Scholar] [CrossRef]
- Cunnane, S.C.; Courchesne-Loyer, A.; St-Pierre, V.; Vandenberghe, C.; Pierotti, T.; Fortier, M.; Croteau, E.; Castellano, C.A. Can ketones compensate for deteriorating brain glucose uptake during aging? Implications for the risk and treatment of Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2016, 1367, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kossoff, E.H.; McGrogan, J.R. Worldwide use of the ketogenic diet. Epilepsia 2005, 46, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Hallböök, T.; Sjölander, A.; Åmark, P.; Miranda, M.; Bjurulf, B.; Dahlin, M. Effectiveness of the ketogenic diet used to treat resistant childhood epilepsy in Scandinavia. Eur. J. Paediatr. Neurol. 2015, 19, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, F.; Aronson, N. Ketogenic diet for the treatment of refractory epilepsy in children: A systematic review of efficacy. Pediatrics 2000, 105, E46. [Google Scholar] [CrossRef] [PubMed]
- Perucca, E.; Gram, L.; Avanzini, G.; Dulac, O. Antiepileptic drugs as a cause of worsening seizures. Epilepsia 1998, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Uhlemann, E.R.; Neims, A.H. Anticonvulsant properties of the ketogenic diet in mice. J. Pharmacol. Exp. Ther. 1972, 180, 231–238. [Google Scholar] [PubMed]
- Stafstrom, C.E. Animal models of the ketogenic diet: What have we learned, what can we learn? Epilepsy Res. 1999, 37, 241–259. [Google Scholar] [CrossRef]
- Likhodii, S.S.; Musa, K.; Mendonca, A.; Dell, C.; Burnham, W.M.; Cunnane, S.C. Dietary fat, ketosis, and seizure resistance in rats on the ketogenic diet. Epilepsia 2000, 41, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Thavendiranathan, P.; Chow, C.; Cunnane, S.; McIntyre, B.W. The effect of the classic ketogenic diet on animal seizure models. Brain Res. 2003, 959, 206–213. [Google Scholar] [CrossRef]
- Thavendiranathan, P.; Mendonca, A.; Dell, C.; Likhodii, S.S.; Musa, K.; Iracleous, C.; Cunnane, S.C.; Burnham, W.M. The MCT ketogenic diet: Effects on animal seizure models. Exp. Neurol. 2000, 161, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Hartman, A.L.; Lyle, M.; Rogawski, M.A.; Gasior, M. Efficacy of the ketogenic diet in the 6-Hz seizure test. Epilepsia 2008, 49, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Samala, R.; Willis, S.; Borges, K. Anticonvulsant profile of a balanced ketogenic diet in acute mouse seizure models. Epilepsy Res. 2008, 81, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Potschka, H.; Löscher, W. Corneal kindling in mice: Behavioral and pharmacological differences to conventional kindling. Epilepsy Res. 1999, 37, 109–120. [Google Scholar] [CrossRef]
- Leclercq, K.; Matagne, A.; Kaminski, R.M. Low potency and limited efficacy of antiepileptic drugs in the mouse 6 Hz corneal kindling model. Epilepsy Res. 2014, 108, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W. Fit for purpose application of currently existing animal models in the discovery of novel epilepsy therapies. Epilepsy Res. 2016, 126, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, Y.; Wang, S.; Ding, Y.; Guo, Y.; Zhang, M.M.; Wen, S.Q.; Ding, M.P. Ketogenic diet protects against epileptogenesis as well as neuronal loss in amygdaloid-kindling seizures. Neurosci. Lett. 2012, 508, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Lusardi, T.A.; Akula, K.K.; Coffman, S.Q.; Ruskin, D.N.; Masino, S.A.; Boison, D. Ketogenic diet prevents epileptogenesis and disease progression in adult mice and rats. Neuropharmacology 2015, 99, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Todorova, M.T.; Tandon, P.; Madore, R.A.; Stafstrom, C.E.; Seyfried, T.N. The ketogenic diet inhibits epileptogenesis in EL mice: A genetic model for idiopathic epilepsy. Epilepsia 2000, 41, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Linard, B.; Ferrandon, A.; Koning, E.; Nehlig, A.; Raffo, E. Ketogenic diet exhibits neuroprotective effects in hippocampus but fails to prevent epileptogenesis in the lithium-pilocarpine model of mesial temporal lobe epilepsy in adult rats. Epilepsia 2010, 51, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Giordano, C.; Vinet, J.; Curia, G.; Biagini, G. Repeated 6-Hz corneal stimulation progressively increases FosB/ΔFosB levels in the lateral amygdala and induces seizure generalization to the hippocampus. PLoS ONE 2015, 10, e0141221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, C.; Costa, A.M.; Lucchi, C.; Leo, G.; Brunel, L.; Fehrentz, J.A.; Martinez, J.; Torsello, A.; Biagini, G. Progressive seizure aggravation in the repeated 6-Hz corneal stimulation model is accompanied by marked increase in hippocampal p-ERK1/2 immunoreactivity in neurons. Front. Cell. Neurosci. 2016, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, C.; Costa, A.M.; Giordano, C.; Curia, G.; Piat, M.; Leo, G.; Vinet, J.; Fehrentz, J.A.; Brunel, L.; Martinez, J.; et al. Involvement of PPARγ in the anticonvulsant activity of EP-80317, a ghrelin receptor antagonist. Front. Pharmacol. 2017, 8, 676. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.E.; Klein, B.D.; Wolf, H.H.; White, H.S. Pharmacological characterization of the 6 Hz psychomotor seizure model of partial epilepsy. Epilepsy Res. 2001, 47, 217–227. [Google Scholar] [CrossRef]
- Hartman, A.L.; Zheng, X.; Bergbower, E.; Kennedy, M.; Hardwick, J.M. Seizure tests distinguish intermittent fasting from the ketogenic diet. Epilepsia 2010, 51, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Wlaź, P.; Socała, K.; Nieoczym, D.; Łuszczki, J.J.; Zarnowska, I.; Zarnowski, T.; Czuczwar, S.J.; Gasior, M. Anticonvulsant profile of caprylic acid, a main constituent of the medium-chain triglyceride (MCT) ketogenic diet, in mice. Neuropharmacology 2012, 62, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Ruskin, D.N.; Svedova, J.; Cote, J.L.; Sandau, U.; Rho, J.M.; Kawamura, M., Jr. Ketogenic diet improves core symptoms of autism in BTBR mice. PLoS ONE 2013, 8, e65021. [Google Scholar] [CrossRef] [PubMed]
- Matagne, A.; Klitgaard, H. Validation of corneally kindled mice: A sensitive screening model for partial epilepsy in man. Epilepsy Res. 1998, 31, 59–71. [Google Scholar] [CrossRef]
- Morimoto, K.; Fahnestock, M.; Racine, R.J. Kindling and status epilepticus models of epilepsy: Rewiring the brain. Prog. Neurobiol. 2004, 73, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, A.; Lukasiuk, K. Mechanisms of epileptogenesis and potential treatment targets. Lancet Neurol. 2011, 10, 173–186. [Google Scholar] [CrossRef]
- Thammongkol, S.; Vears, D.F.; Bicknell-Royle, J.; Nation, J.; Draffin, K.; Stewart, K.G.; Scheffer, I.E.; Mackay, M.T. Efficacy of the ketogenic diet: Which epilepsies respond? Epilepsia 2012, 53, e55–e59. [Google Scholar] [CrossRef] [PubMed]
- Castel-Branco, M.M.; Alves, G.L.; Figueiredo, I.V.; Falcão, A.C.; Caramona, M.M. The maximal electroshock seizure (MES) model in the preclinical assessment of potential new antiepileptic drugs. Methods Find. Exp. Clin. Pharmacol. 2009, 31, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Van der Louw, E.; Van den Hurk, D.; Neal, E.; Leiendecker, B.; Fitzsimmon, G.; Dority, L.; Thompson, L.; Marchió, M.; Dudzińska, M.; Dressler, A.; et al. Ketogenic diet guidelines for infants with refractory epilepsy. Eur. J. Paediatr. Neurol. 2016, 20, 798–809. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Gender | Age | Diagnosis | Type of Seizure | AEDs | Type of KD | Response (Reduction in Seizure Frequency) | Transient Side Effects |
|---|---|---|---|---|---|---|---|---|
| #1 | F | 5 | Respiratory chain defect I-III complex | Spasm | CBZ, NZP, LEV | Classic 2:1 | 88% | Hypercholesterolemia, hyperoxaluria |
| #2 | F | 4 | Suspected PDCD | Tonic-clonic, spasm | NZP, LEV | Classic 3:1 | 92% | Nausea, vomiting |
| #3 | F | 9.6 | Lennox–Gastaut syndrome | Spasm, drop attack, tonic-clonic | LEV, RFN, NZP | Classic 4:1 | Transient worsening, then 78% reduction | Hypertriglyceridemia, acidosis |
| #4 | F | 5 | Congenital hydrocephalus | Drop attack, myoclonus, absence | TPM, LEV | Classic 2:1 | 94% | Hypercholesterolemia |
| #5 | M | 5.9 | Suspect of GLUT1 deficiency | Tonic-clonic, spasm | TPM, CZP | Classic 4:1 | 52% | Hyperphosphatemia |
| #6 | M | 5.9 | Epileptic encephalopathy | Tonic-clonic, spasm | GVG, VPA, TPM | MCT 3:1 | 97% | No |
| #7 | F | 5.4 | Partial trisomy for chromosome 13 | Tonic-clonic, drop attack | VPA, NZP, RFN | MCT 3.5:1 | 96% | Nausea, vomiting |
| #8 | M | 5.3 | Epileptic encephalopathy | Tonic-clonic | TPM, ESM | Classic 3:1 | 95% | No |
| #9 | M | 4.7 | Suspected GLUT1 deficiency | Myoclonus, tonic-clonic | GVG, CBZ, NZP | Classic 3:1 | 97% | Hypercholesterolemia |
| #10 | M | 4 | Lissencephaly type 1 | Tonic-clonic, spasm | CZP, PB, CBZ | Classic 4:1 + MCT | Worsened | Nausea, vomiting |
| #11 | F | 3.9 | Dravet syndrome | Myoclonus | TPM, CZP, CLB | Classic 2:1 | 48% | No |
| #12 | F | 9.3 | GLUT1 deficiency | Absence, spasm | VPA | Classic 2.5:1 | 100% | Hypercholesterolemia |
| #13 | M | 3 | Epileptic encephalopathy | Tonic-clonic, drop attack | TPM, CZP, CLB | Classic 4:1 | 45% | No |
| #14 | F | 9.6 | Epileptic encephalopathy | Absence, spasm | VPA, LTG, CLB | Classic 4:1 | 91% | Constipation |
| #15 | M | 8.2 | Suspected PDCD | Tremor | NZP, LEV | Classic 3:1 | 53% | No |
| #16 | F | 6 | Cerebral cortex malformation | Absence, tonic-clonic | LEV, LTG | MCT 3:1 | 63% | Constipation |
| #17 | F | 6.5 | West Syndrome | Tonic-clonic | TAS, CBZ, ESM | Classic 2:1 | 0% | No |
| #18 | F | 7.1 | Lissencephaly type 1 | Tonic-clonic, spasm, absence | VPA, CLB, RFN | Classic 2:1 | Worsened | No |
| #19 | F | 14 | Cryptogenic generalized epilepsy | Tonic-clonic, absence | LEV, CZP, PB | Classic 3:1 | 73% | No |
| #20 | F | 14.3 | Epileptic encephalopathy | Tonic-clonic | LEV, ZSN, NZP | Classic 4:1 | 63% | Hypercholesterolemia, weight gain |
| #21 | F | 10 | Epileptic encephalopathy | Tonic-clonic, absence | LTG, LEV | MCT 3:1 | 51% | Vomiting, constipation |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucchi, C.; Marchiò, M.; Caramaschi, E.; Giordano, C.; Giordano, R.; Guerra, A.; Biagini, G. Electrographic Changes Accompanying Recurrent Seizures under Ketogenic Diet Treatment. Pharmaceuticals 2017, 10, 82. https://doi.org/10.3390/ph10040082
Lucchi C, Marchiò M, Caramaschi E, Giordano C, Giordano R, Guerra A, Biagini G. Electrographic Changes Accompanying Recurrent Seizures under Ketogenic Diet Treatment. Pharmaceuticals. 2017; 10(4):82. https://doi.org/10.3390/ph10040082
Chicago/Turabian StyleLucchi, Chiara, Maddalena Marchiò, Elisa Caramaschi, Carmela Giordano, Rocco Giordano, Azzurra Guerra, and Giuseppe Biagini. 2017. "Electrographic Changes Accompanying Recurrent Seizures under Ketogenic Diet Treatment" Pharmaceuticals 10, no. 4: 82. https://doi.org/10.3390/ph10040082
APA StyleLucchi, C., Marchiò, M., Caramaschi, E., Giordano, C., Giordano, R., Guerra, A., & Biagini, G. (2017). Electrographic Changes Accompanying Recurrent Seizures under Ketogenic Diet Treatment. Pharmaceuticals, 10(4), 82. https://doi.org/10.3390/ph10040082