Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
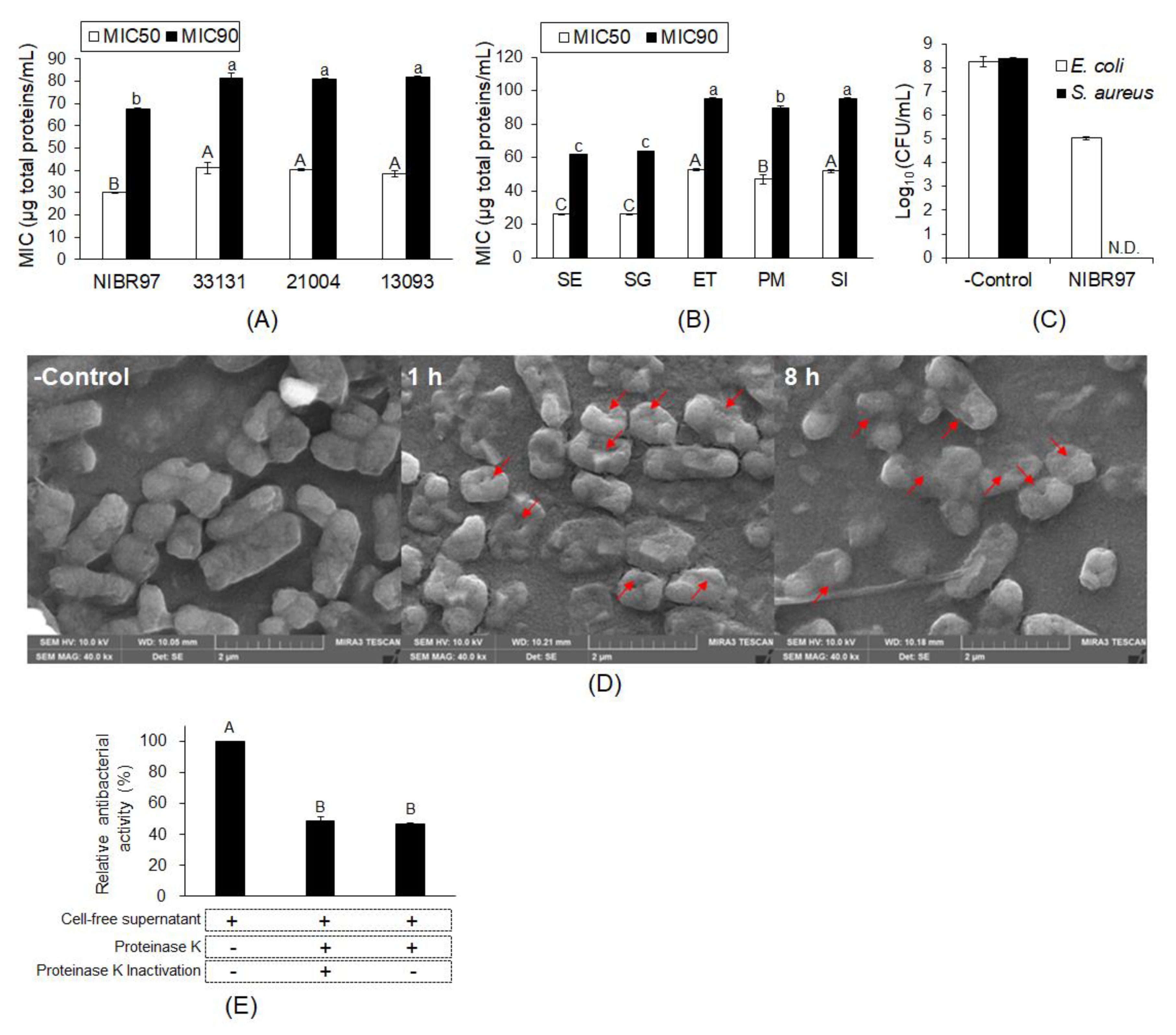
2.1. Antibacterial Activity of Cell-Free Supernatant from L. plantarum NIBR97
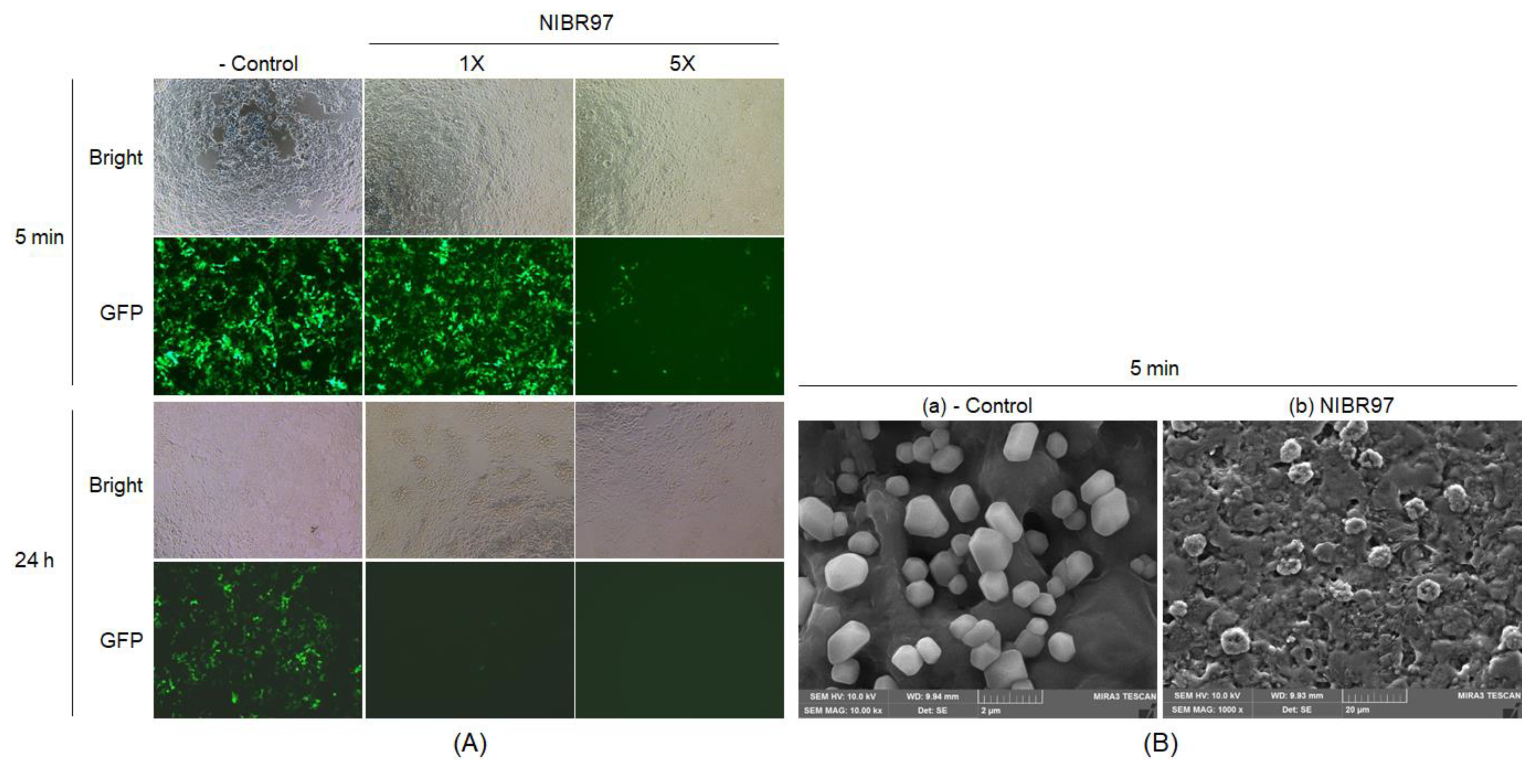
2.2. Antiviral Activity of Cell-Free Supernatant from L. plantarum NIBR97
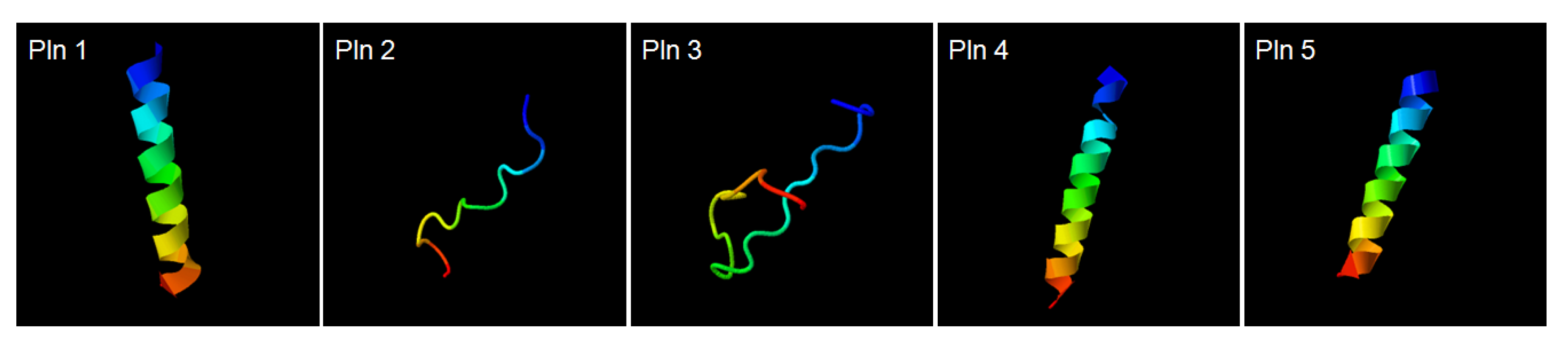
2.3. Discovery of Novel Bacteriocins by the Genomic Analysis of L. plantarum NIBR97
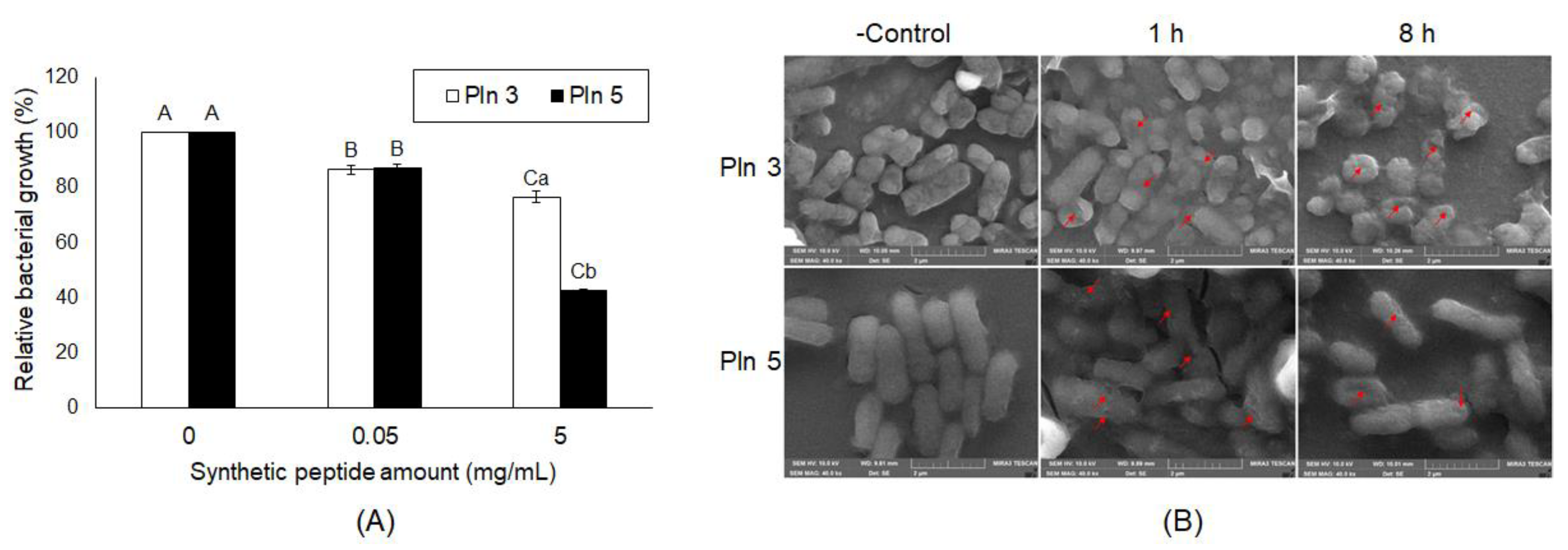
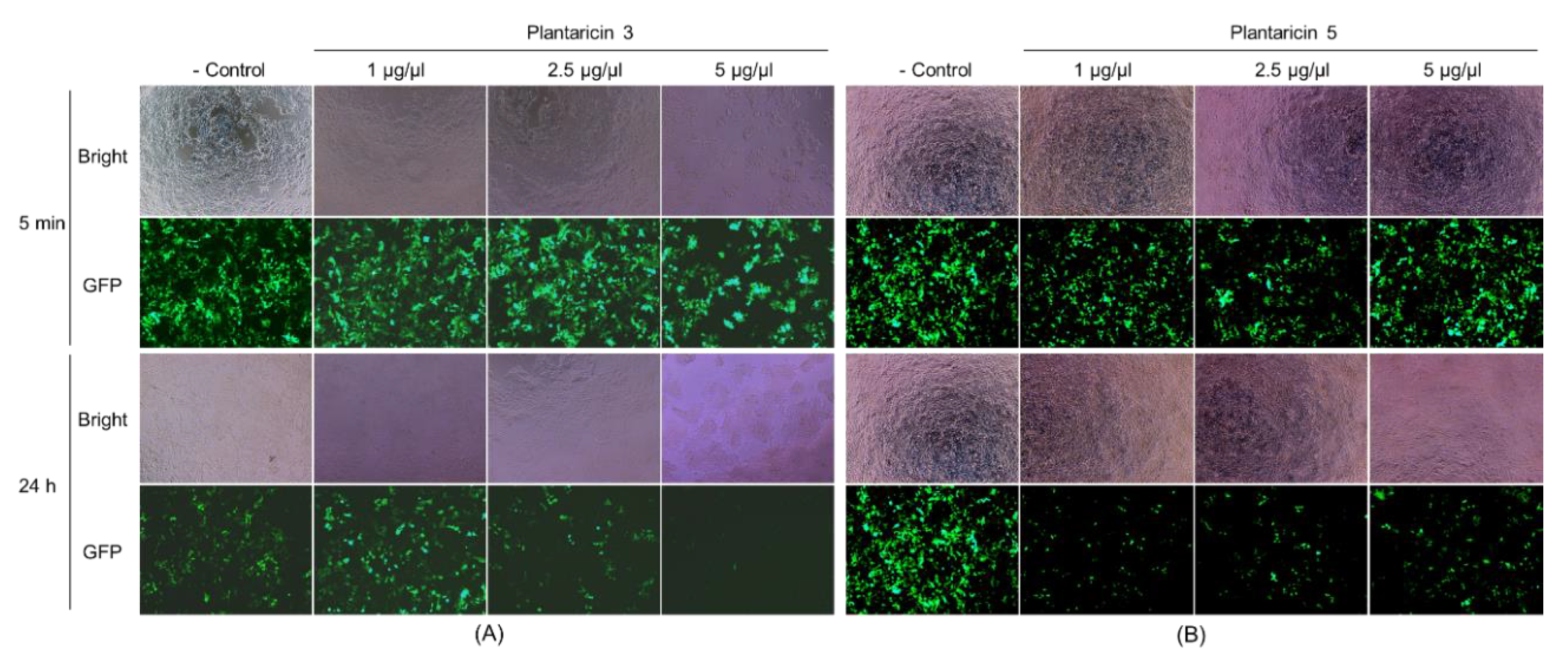
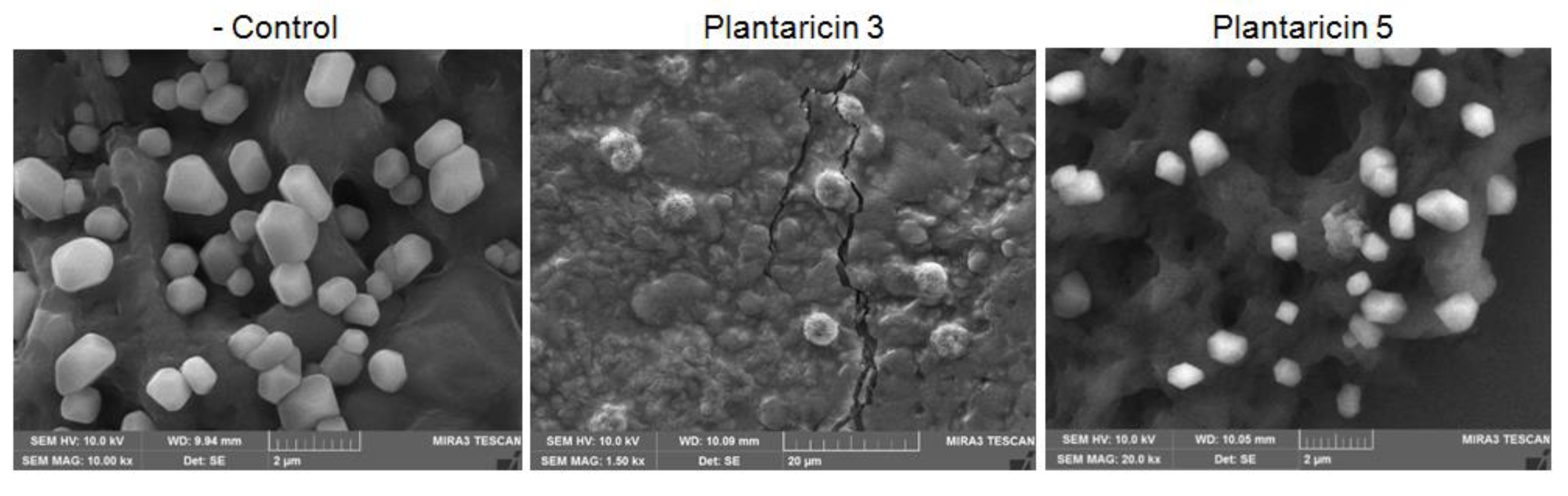
2.4. Antibacterial and Antiviral Activities of Plantaricins from L. plantarum NIBR97
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Analysis of the Minimal Inhibitory Concentration (MIC50 and MIC90)
4.3. Measurement of Bactericidal Activity
4.4. Scanning Electron Microscopy (SEM)
4.5. Antiviral Analysis Against Influenza A/H3N2
4.6. Antiviral Analysis Against GFP-Labeled Lentivirus
4.7. Analysis of the Genome
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Coronavirus Disease (COVID-19): Situation Report, 150; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- World Health Organization. Cleaning and Disinfection of Environmental Surfaces in the Context of COVID-19: Interim Guidance; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Atolani, O.; Baker, M.T.; Adeyemi, O.S.; Olanrewaju, I.R.; Hamid, A.A.; Ameen, O.M.; Oguntoye, S.O.; Usman, L.A. COVID-19: Critical discussion on the applications and implications of chemicals in sanitizers and disinfectants. EXCLI J. 2020, 19, 785. [Google Scholar] [PubMed]
- Pradhan, D.; Biswasroy, P.; Ghosh, G.; Rath, G. A review of current interventions for COVID-19 prevention. Arch. Med. Res. 2020, 51, 363–374. [Google Scholar] [CrossRef]
- Berardi, A.; Perinelli, D.R.; Merchant, H.A.; Bisharat, L.; Basheti, I.A.; Bonacucina, G.; Cespi, M.; Palmieri, G.F. Hand sanitisers amid CoViD-19: A critical review of alcohol-based products on the market and formulation approaches to respond to increasing demand. Int. J. Pharm. 2020, 584, 119431. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, O.O. Classification of Antimicrobial Peptides Bacteriocins, and the Nature of Some Bacteriocins with Potential Applications in Food Safety and Bio-Pharmaceuticals. EC Microbiol. 2019, 15, 591–608. [Google Scholar]
- Stanojević-Nikolić, S.; Dimić, G.; Mojović, L.; Pejin, J.; Djukić-Vuković, A.; Kocić-Tanackov, S. Antimicrobial activity of lactic acid against pathogen and spoilage microorganisms. J. Food Process. Preserv. 2016, 40, 990–998. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Al Karaawi, M.A.; Siddiqui, M.U. Antimicrobial potential of bacteriocins: In therapy, agriculture and food preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef]
- Hashim, H.; Sikandar, S.; Khan, M.A.; Qurashi, A.W. Bacteriocin: The avenues of innovation towards applied microbiology. Pure Appl. Biol. (PAB) 2019, 8, 460–478. [Google Scholar] [CrossRef]
- Cerqueira, J.; Dimitrov, S.; Silva, A.; Augusto, L. Inhibition of Herpes simplex virus 1 and Poliovirus (PV-1) by bacteriocins from Lactococcus lactis subsp. lactis and Enterococcus durans strains isolated from goat milk. Int. J. Antimicrob. Agents 2018, 51, 33–37. [Google Scholar]
- Ermolenko, E.; Desheva, Y.; Kolobov, A.; Kotyleva, M.; Sychev, I.; Suvorov, A. Anti–Influenza Activity of Enterocin B In vitro and Protective Effect of Bacteriocinogenic Enterococcal Probiotic Strain on Influenza Infection in Mouse Model. Probiotics Antimicrob. Proteins 2019, 11, 705–712. [Google Scholar] [CrossRef]
- Park, S.-C.; Park, Y.; Hahm, K.-S. The role of antimicrobial peptides in preventing multidrug-resistant bacterial infections and biofilm formation. Int. J. Mol. Sci. 2011, 12, 5971–5992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, P.M.; O’Shea, E.F.; Cotter, P.D.; Hill, C.; Ross, R.P. The potency of the broad spectrum bacteriocin, bactofencin A, against staphylococci is highly dependent on primary structure, N-terminal charge and disulphide formation. Sci. Rep. 2018, 8, 1–8. [Google Scholar]
- Vescovo, M.; Bottazzi, V.; Torriani, S.; Dellaglio, F. Basic characteristics, ecology and application of Lactobacillus plantarum [in the production of fermented foods of animal and plant origin]: A review. Ann. Microbiol. Enzimol. (Italy) 1993, 43, 261–284. [Google Scholar]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial activity of Lactobacillus plantarum strains isolated from different environments: A preliminary study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Dinev, T.; Beev, G.; Tzanova, M.; Denev, S.; Dermendzhieva, D.; Stoyanova, A. Antimicrobial activity of Lactobacillus plantarum against pathogenic and food spoilage microorganisms: A review. Bulg. J. Vet. Med. 2018, 21, 253–268. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Diep, D.B.; Håvarstein, L.S.; Nes, I.F. Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11. J. Bacteriol. 1996, 178, 4472–4483. [Google Scholar] [CrossRef] [Green Version]
- Young, R. Bacteriophage holins: Deadly diversity. J. Mol. Microbiol. Biotechnol. 2002, 4, 21–36. [Google Scholar]
- Nissilä, E.; Douillard, F.P.; Ritari, J.; Paulin, L.; Järvinen, H.M.; Rasinkangas, P.; Haapasalo, K.; Meri, S.; Jarva, H.; De Vos, W.M. Genotypic and phenotypic diversity of Lactobacillus rhamnosus clinical isolates, their comparison with strain GG and their recognition by complement system. PLoS ONE 2017, 12, e0176739. [Google Scholar]
- Karpiński, T.M. Efficacy of octenidine against Pseudomonas aeruginosa strains. Eur. J. Biolog. Res. 2019, 9, 135–140. [Google Scholar]
- Hsieh, I.-N.; Hartshorn, K.L. The role of antimicrobial peptides in influenza virus infection and their potential as antiviral and immunomodulatory therapy. Pharmaceuticals 2016, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human antimicrobial peptides as therapeutics for viral infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desriac, N.; Broussolle, V.; Postollec, F.; Mathot, A.-G.; Sohier, D.; Coroller, L.; Leguerinel, I. Bacillus cereus cell response upon exposure to acid environment: Toward the identification of potential biomarkers. Front. Microbiol. 2013, 4, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Ha, Y.J.; Bang, K.H.; Lee, S.; Yeo, J.-H.; Yang, H.-S.; Kim, T.-W.; Lee, K.P.; Bang, W.Y. Potential of Bacteriocins from Lactobacillus taiwanensis for Producing Bacterial Ghosts as a Next Generation Vaccine. Toxins 2020, 12, 432. [Google Scholar] [CrossRef]
- Jang, Y.; Shin, J.S.; Lee, J.-Y.; Shin, H.; Kim, S.J.; Kim, M. In Vitro and In Vivo Antiviral Activity of Nylidrin by Targeting the Hemagglutinin 2-Mediated Membrane Fusion of Influenza A Virus. Viruses 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.-S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biolog. 2004, 5, R12. [Google Scholar] [CrossRef] [Green Version]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagesen, K.; Hallin, P.; Rødland, E.; Stærfeldt, H.; Rognes, T.; Ussery, D. RNammer: Consistent annotation of rRNA genes in genomic sequences. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments 1 | 10 min 1 | 30 min 1 | 18 h 1 | |||
|---|---|---|---|---|---|---|
| Titer 2 | Reduction 3 | Titer 2 | Reduction 3 | Titer 2 | Reduction 3 | |
| Water | 5.66 | 0 | 5.45 | 0 | 5.34 | 0.21 |
| NIBR97 | 3.27 | 99.594 | <0.51 | >99.999 | <0.51 | >99.999 |
| Items | Contig 1 | Contig 2 | Contig 3 | Contig 4 | Contig 5 |
|---|---|---|---|---|---|
| Form | A circular chromosome | A circular plasmid | A circular plasmid | A linear plasmid | A linear plasmid |
| Length 1 | 3,022,780 | 61,378 | 32,520 | 7394 | 6876 |
| GC 2 | 44.74 | 39.22 | 39.59 | 34.33 | 35.67 |
| ORF 3 | 2816 | 60 | 32 | 10 | 9 |
| rRNA 4 | 16 | 0 | 0 | 0 | 0 |
| tRNA 5 | 68 | 0 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.W.; Kang, S.I.; Shin, D.H.; Oh, S.Y.; Lee, C.W.; Yang, Y.; Son, Y.K.; Yang, H.-S.; Lee, B.-H.; An, H.-J.; et al. Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants. Pharmaceuticals 2020, 13, 266. https://doi.org/10.3390/ph13100266
Kim SW, Kang SI, Shin DH, Oh SY, Lee CW, Yang Y, Son YK, Yang H-S, Lee B-H, An H-J, et al. Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants. Pharmaceuticals. 2020; 13(10):266. https://doi.org/10.3390/ph13100266
Chicago/Turabian StyleKim, Sam Woong, Song I. Kang, Da Hye Shin, Se Yun Oh, Chae Won Lee, Yoonyong Yang, Youn Kyoung Son, Hee-Sun Yang, Byoung-Hee Lee, Hee-Jung An, and et al. 2020. "Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants" Pharmaceuticals 13, no. 10: 266. https://doi.org/10.3390/ph13100266
APA StyleKim, S. W., Kang, S. I., Shin, D. H., Oh, S. Y., Lee, C. W., Yang, Y., Son, Y. K., Yang, H. -S., Lee, B. -H., An, H. -J., Jeong, I. S., & Bang, W. Y. (2020). Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants. Pharmaceuticals, 13(10), 266. https://doi.org/10.3390/ph13100266