Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters


 , and
, and
Abstract
:1. Introduction
2. Results
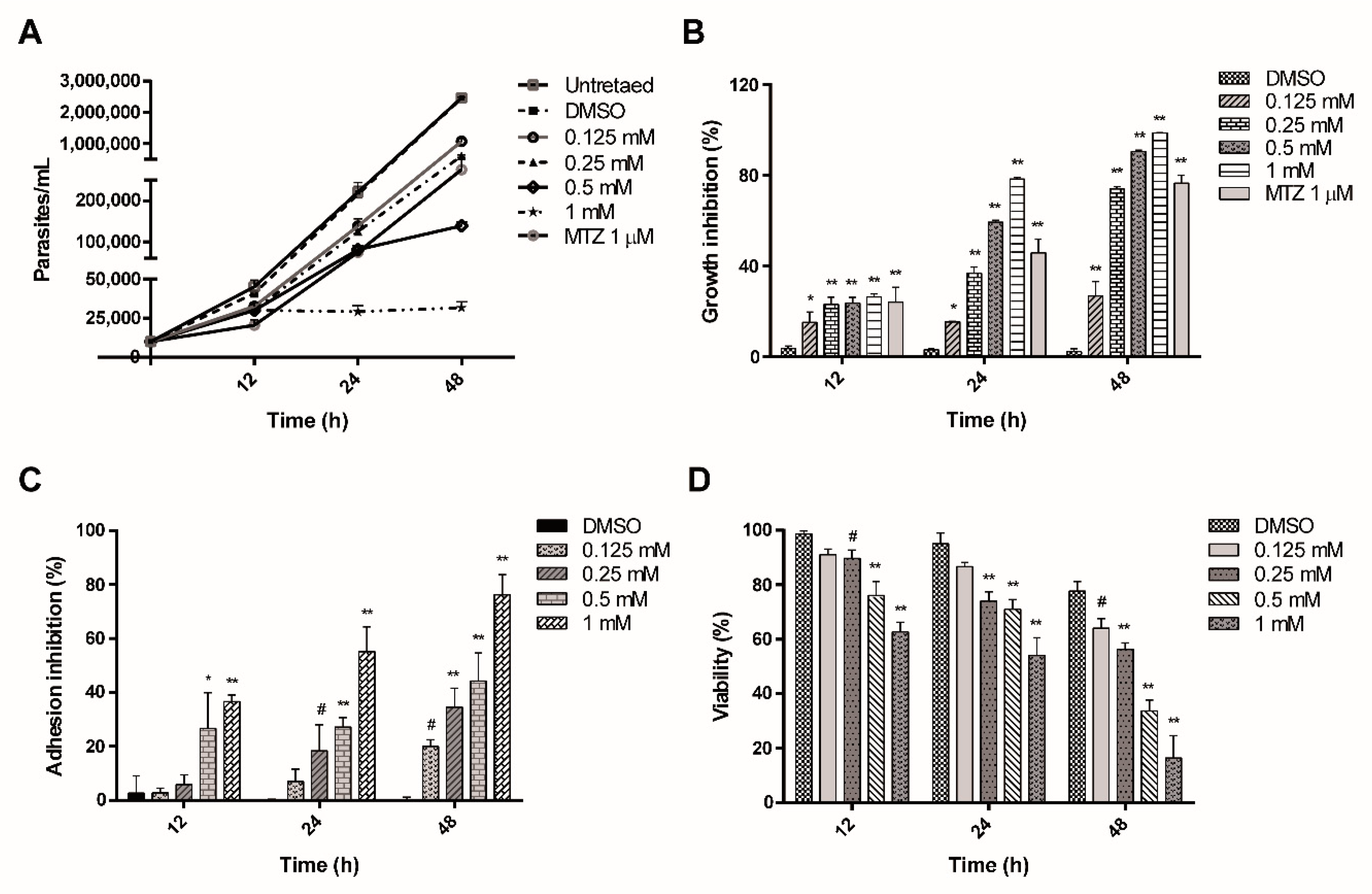
2.1. ASA Inhibits the Growth, Adhesion and Cell Viability of Giardia lamblia Trophozoites
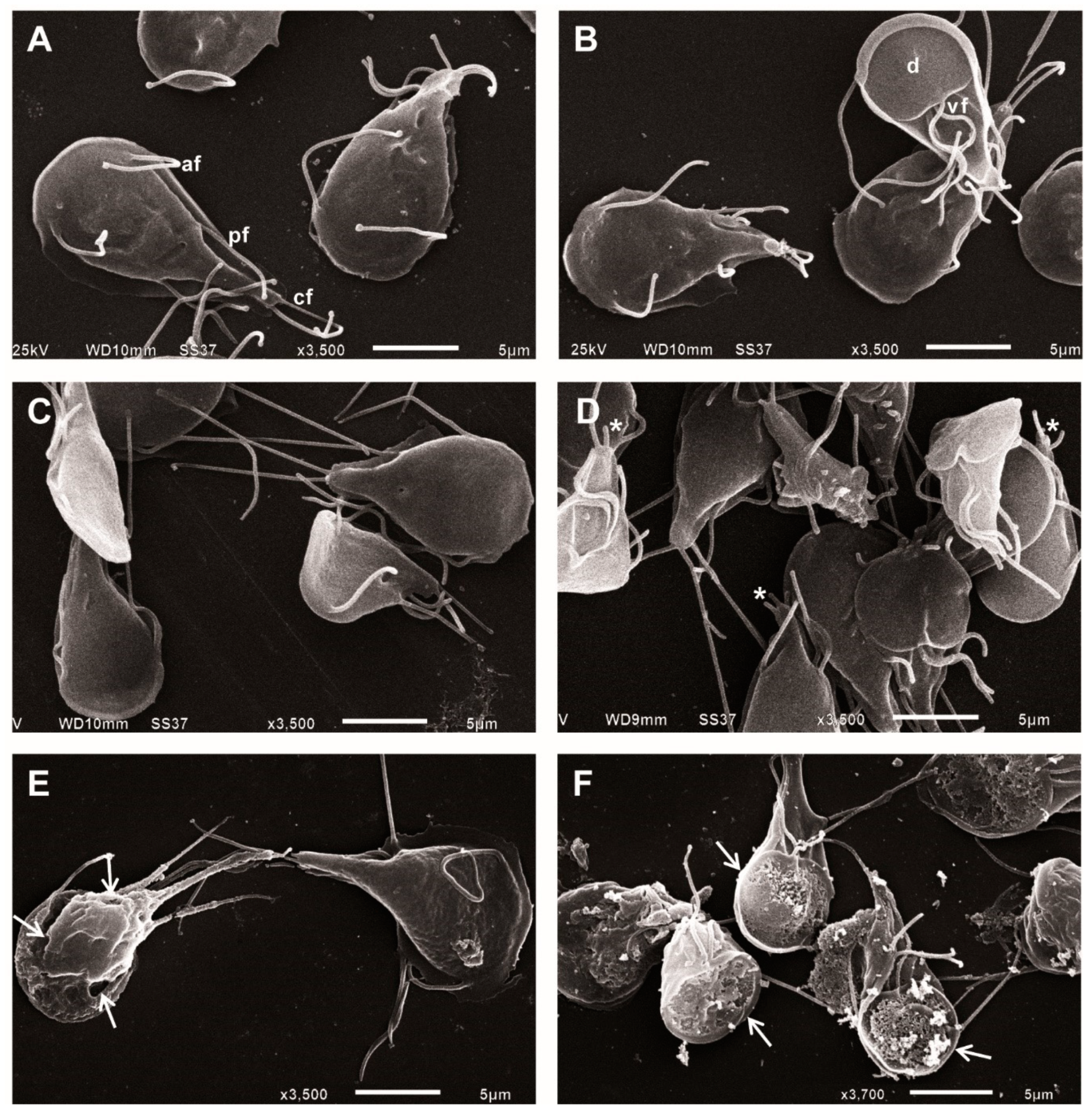
2.2. ASA Alters the Morphology of Giardia lamblia Trophozoites
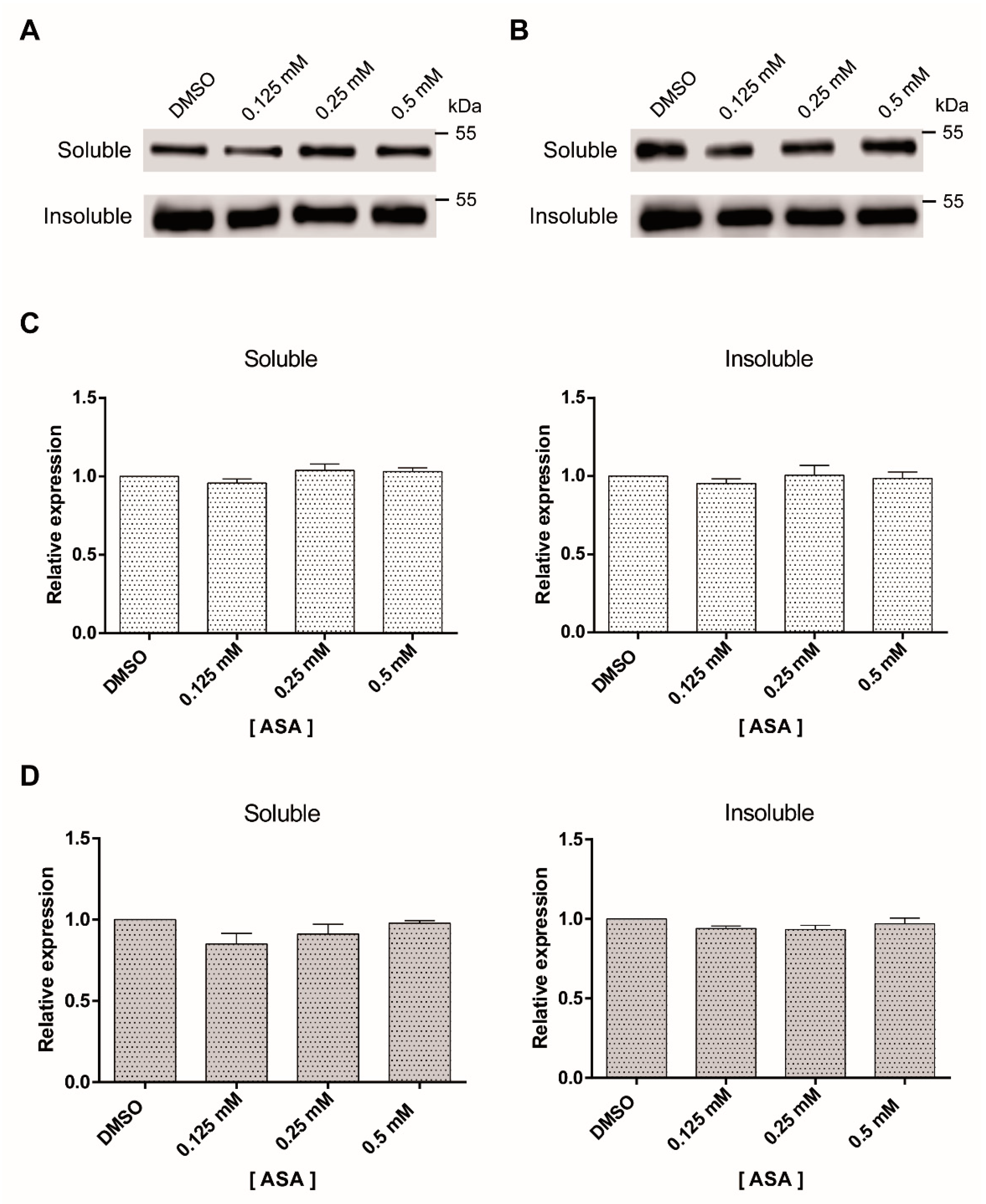
2.3. Tubulin Expression Is Not Modified by ASA Treatment
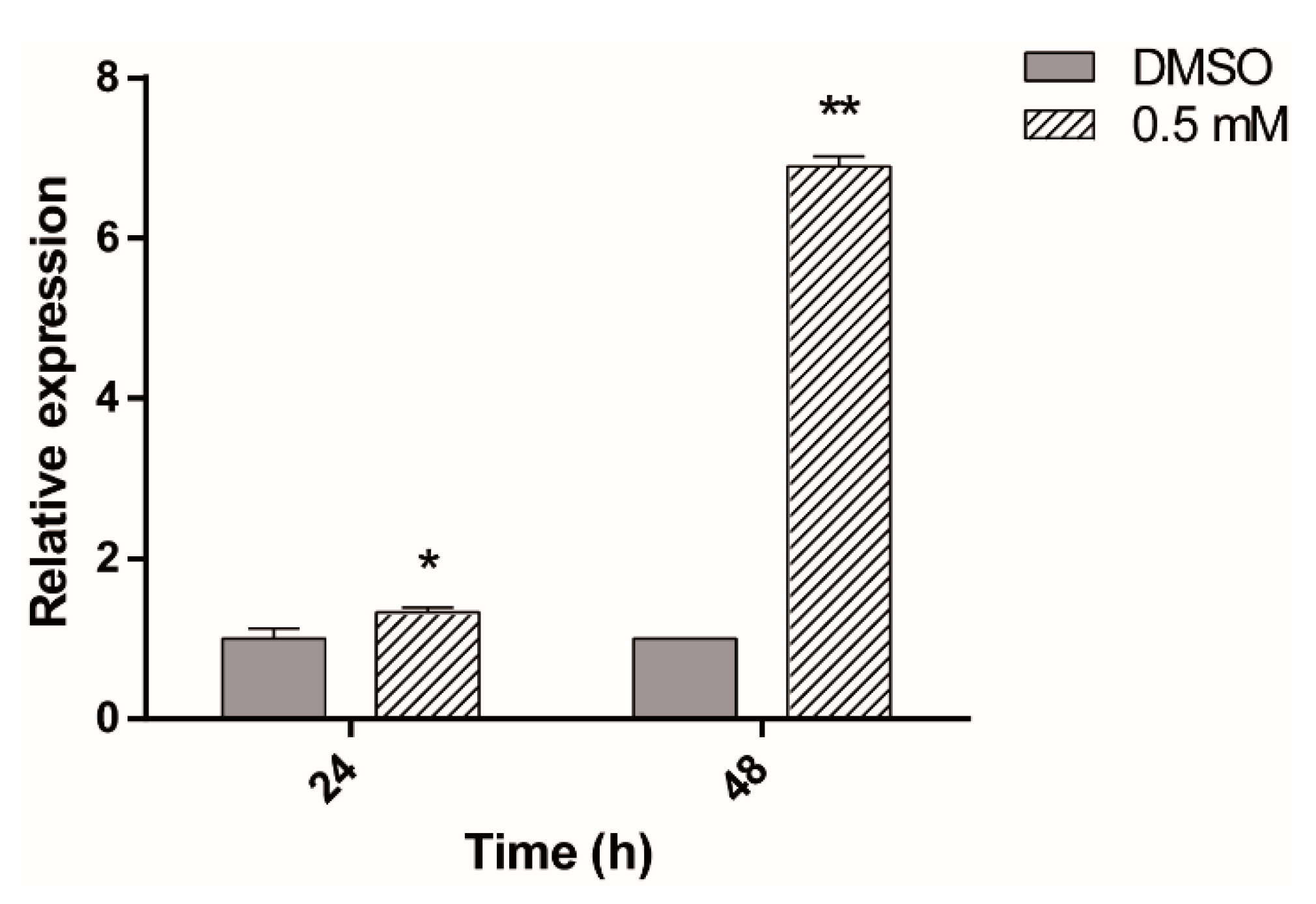
2.4. HSP70 Is Associated with ASA Damage in Giardia
2.5. ASA Induced Hsp70 Overexpression in Giardia lamblia Trophozoites
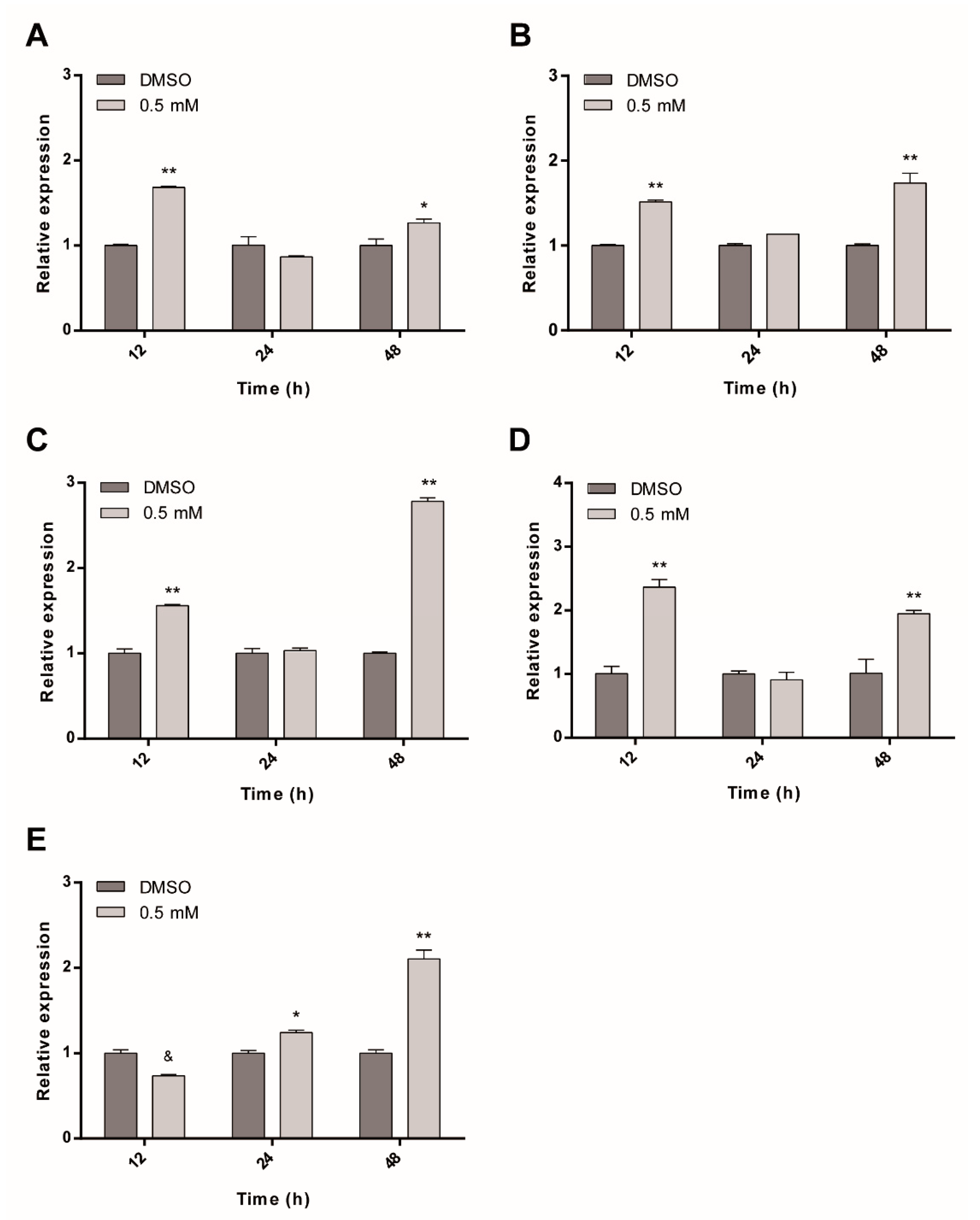
2.6. ASA Modifies the Expression Level of ABC/MDR Transporter Genes
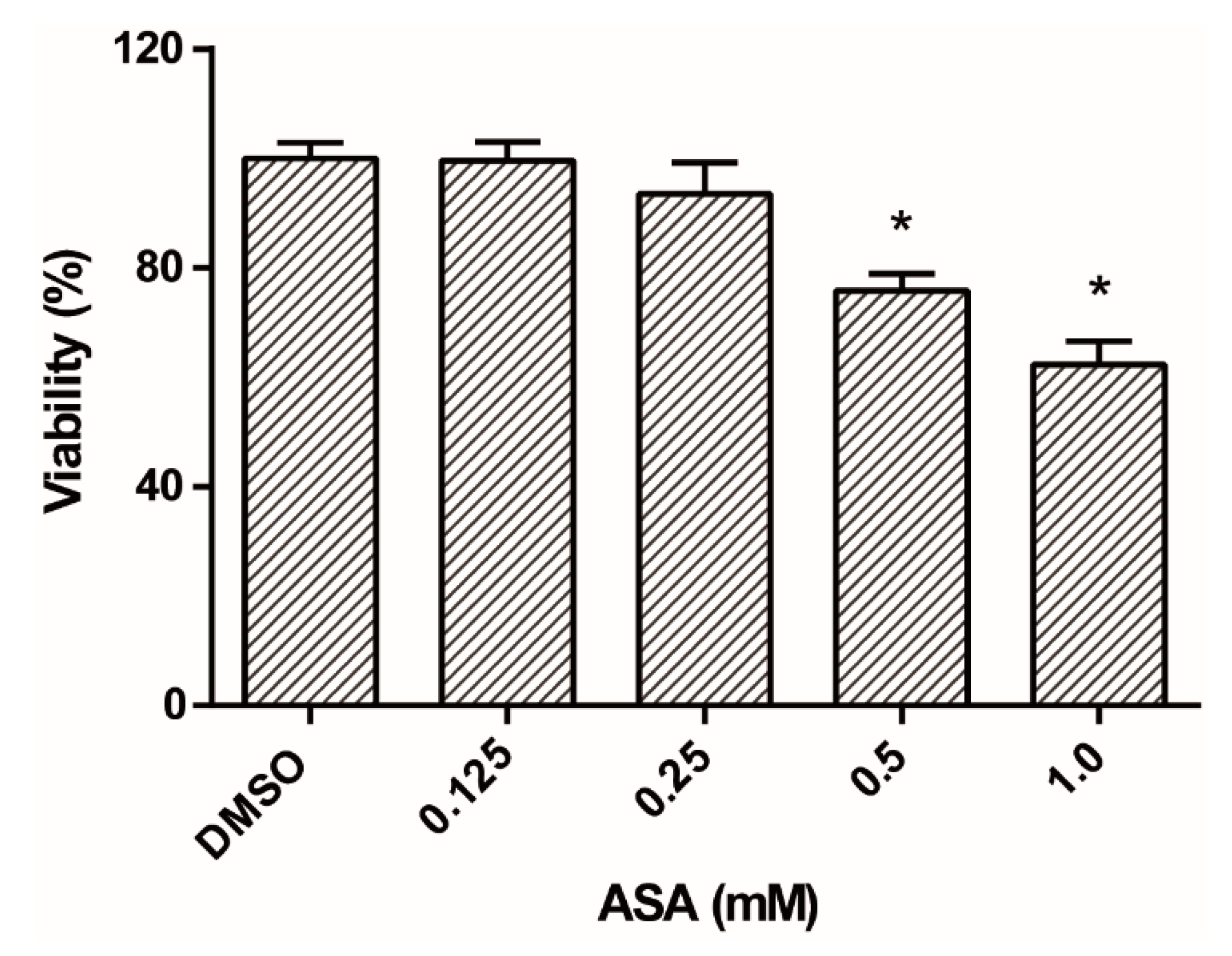
2.7. ASA Is Partially Cytotoxic to Caco-2 Cells at High Doses
3. Discussion
4. Materials and Methods
4.1. Giardia Lamblia Trophozoites Culture
4.2. Growth Inhibition Assay
4.3. Cell Viability Assay
4.4. Adherence Assay
4.5. Scanning Electron Microscopy (SEM)
4.6. Preparation of Protein Fractions and Western Blotting
4.7. Protein Extract and SDS-PAGE
4.8. Tryptic Digest Protocol Subsequent to Coomassie Staining
4.9. Ultra-Performance Liquid Chromatography-Time of Flight Mass Spectrometry (UPLC-TOF/MS)
4.10. Bioinformatics Analysis of Giardia Genome
4.11. Relative-Quantitative RT-PCR
4.12. ASA Selectivity Index Calculation
4.12.1. Culture of Human Intestinal Caco-2 Cells
4.12.2. Cell Viability (MTT Assay)
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leung, A.K.; Leung, A.A.; Wong, A.H.; Sergi, C.M.; Kam, J.K. Giardiasis: An Overview. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 134–143. [Google Scholar] [CrossRef]
- Vivancos, V.; González-Alvarez, I.; Bermejo, M. Giardiasis: Characteristics, Pathogenesis and New Insights About Treatment. Curr. Top. Med. Chem. 2018, 18, 1287–1303. [Google Scholar] [CrossRef]
- Dunn, N.; Juergens, A.L. Giardiasis. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ansell, B.R.; McConville, M.J.; Ma’Ayeh, S.Y.; Dagley, M.J.; Gasser, R.B.; Svärd, S.G.; Jex, A.R. Drug resistance in Giardia duodenalis. Biotechnol. Adv. 2015, 33, 888–901. [Google Scholar] [CrossRef]
- Leitsch, D. Drug Resistance in the Microaerophilic Parasite Giardia lamblia. Curr. Trop. Med. Rep. 2015, 2, 128–135. [Google Scholar] [CrossRef] [Green Version]
- DiMasi, J.A.; Hansen, R.W.; Grabowski, H.G. The price of innovation: New estimates of drug development costs. J. Health Econ. 2003, 22, 151–185. [Google Scholar] [CrossRef] [Green Version]
- DiMasi, J.A. Risks in new drug development: Approval success rates for investigational drugs. Clin. Pharmacol. Ther. 2001, 69, 297–307. [Google Scholar] [CrossRef]
- Lei, J.; Zhou, Y.; Xie, D.; Zhang, Y. Mechanistic Insights into a Classic Wonder Drug—Aspirin. J. Am. Chem. Soc. 2014, 137, 70–73. [Google Scholar] [CrossRef] [Green Version]
- Flossmann, E.; Rothwell, P.M. Effect of aspirin on long-term risk of colorectal cancer: Consistent evidence from randomised and observational studies. Lancet 2007, 369, 1603–1613. [Google Scholar] [CrossRef]
- Chan, A.T.; Giovannucci, E.L. Primary prevention of colorectal cancer. Gastroenterology 2010, 138, 2029–2043.e10. [Google Scholar] [CrossRef] [Green Version]
- Bhat, G.; Marimuthu, S.; Chivukula, R.S.; Alfonso, L.F.; Moridani, M.; Hagen, F.K. Aspirin acetylates multiple cellular proteins in HCT-116 colon cancer cells: Identification of novel targets. Int. J. Oncol. 2011, 39, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Rathaur, S. Combination of DEC plus aspirin induced mitochondrial mediated apoptosis in filarial parasite Setaria cervi. Biochimie 2010, 92, 894–900. [Google Scholar] [CrossRef]
- Contreras, L.L.; Hernández-Ramírez, V.I.; Lagunes-Guillén, A.E.; Montaño, S.; Chávez-Munguía, B.; Sánchez-Ramírez, B.; Talamás-Rohana, P. Exploring the Possible Role of Lysine Acetylation on Entamoeba histolytica Virulence: A Focus on the Dynamics of the Actin Cytoskeleton. BioMed Res. Int. 2013, 2013, 757392. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.-Y.; Liu, M.-S.; Zhang, C.; Cai, P.-C.; Xiao, Z.-H.; Wang, F.-F. Aspirin enhances the sensitivity of hepatocellular carcinoma side population cells to doxorubicin via miR-491/ABCG2. Biosci. Rep. 2018, 38, 20180854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugai, M.; Uchiyama, K.; Tsuji, T.; Yoriki, H.; Fukui, A.; Qin, Y.; Higashimura, Y.; Mizushima, K.; Yoshida, N.; Katada, K.; et al. MDR1 is Related to Intestinal Epithelial Injury Induced by Acetylsalicylic Acid. Cell. Physiol. Biochem. 2013, 32, 942–950. [Google Scholar] [CrossRef]
- Massimi, I.; Ciuffetta, A.; Temperilli, F.; Ferrandino, F.; Zicari, A.; Pulcinelli, F.M.; Felli, M.P. Multidrug Resistance Protein-4 Influences Aspirin Toxicity in Human Cell Line. Mediat. Inflamm. 2015, 2015, 607957. [Google Scholar] [CrossRef] [Green Version]
- Klokouzas, A. ABC transporters and drug resistance in parasitic protozoa. Int. J. Antimicrob. Agents 2003, 22, 301–317. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Yeo, G.-S.; Lim, Y.-S.; Kang, C.-D.; Kim, C.-M.; Chung, B.-S. Suppression of multidrug resistance via inhibition of heat shock factor by quercetin in MDR cells. Exp. Mol. Med. 1998, 30, 87–92. [Google Scholar] [CrossRef]
- Vilaboa, N.E.; Galán, A.; Troyano, A.; De Blas, E.; Aller, P. Regulation of Multidrug Resistance 1 (MDR1)/P-glycoprotein Gene Expression and Activity by Heat-Shock Transcription Factor 1 (HSF1). J. Biol. Chem. 2000, 275, 24970–24976. [Google Scholar] [CrossRef] [Green Version]
- Kanagasabai, R.; Krishnamurthy, K.; Druhan, L.J.; Ilangovan, G. Forced Expression of Heat Shock Protein 27 (Hsp27) Reverses P-Glycoprotein (ABCB1)-mediated Drug Efflux andMDR1Gene Expression in Adriamycin-resistant Human Breast Cancer Cells. J. Biol. Chem. 2011, 286, 33289–33300. [Google Scholar] [CrossRef] [Green Version]
- Tchénio, T.; Havard, M.; Martinez, L.A.; Dautry, F. Heat Shock-Independent Induction of Multidrug Resistance by Heat Shock Factor 1. Mol. Cell. Biol. 2006, 26, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Granados, C.E.; Reveiz, L.; Uribe, L.G.; Criollo, C.P. Drugs for treating giardiasis. Cochrane Database Syst. Rev. 2012, 12, CD007787. [Google Scholar] [CrossRef]
- Lalle, M.; Hanevik, K. Treatment-refractory giardiasis: Challenges and solutions. Infect. Drug Resist. 2018, 11, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Lalle, M. Giardiasis in the Post Genomic Era: Treatment, Drug Resistance and Novel Therapeutic Perspectives. Infect. Disord. Drug Targets 2010, 10, 283–294. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bakri, A.; Othman, G.; Bustanji, Y. The assessment of the antibacterial and antifungal activities of aspirin, EDTA and aspirin-EDTA combination and their effectiveness as antibiofilm agents. J. Appl. Microbiol. 2009, 107, 280–286. [Google Scholar] [CrossRef]
- López-Muñoz, R.; Faundez, M.; Klein, S.; Escanilla, S.; Torres, G.; Lee-Liu, D.; Ferreira, J.; Kemmerling, U.; Orellana, M.; Morello, A.; et al. Trypanosoma cruzi: In vitro effect of aspirin with nifurtimox and benznidazole. Exp. Parasitol. 2010, 124, 167–171. [Google Scholar] [CrossRef]
- De Souza, N.D.; Belin, B.S.; Massocatto, C.L.; De Araújo, S.M.; Sant’Ana, D.M.; Araújo, E.J.; Filho, P.P.; Nihei, O.K.; Moreira, N.M. Effect of acetylsalicylic acid on total myenteric neurons in mice experimentally infected with Trypanosoma cruzi. Anais da Academia Brasileira de Ciências 2019, 91, e20180389. [Google Scholar] [CrossRef]
- Dovizio, M.; Bruno, A.; Tacconelli, S.; Patrignani, P. Mode of Action of Aspirin as a Chemopreventive Agent. In Prospects for Chemoprevention of Colorectal Neoplasia; Springer: Berlin/Heidelberg, Germany, 2013; pp. 39–65. [Google Scholar]
- Bahadur, V.; Mastronicola, D.; Tiwari, H.K.; Kumar, Y.; Falabella, M.; Pucillo, L.P.; Sarti, P.; Giuffrè, A.; Singh, B.K. O2-Dependent Efficacy of Novel Piperidine- and Piperazine-Based Chalcones against the Human Parasite Giardia intestinalis. Antimicrob. Agents Chemother. 2013, 58, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Bahadur, V.; Mastronicola, D.; Singh, A.K.; Tiwari, H.K.; Pucillo, L.P.; Sarti, P.; Singh, B.K.; Giuffrè, A. Antigiardial activity of novel triazolyl-quinolone-based chalcone derivatives: When oxygen makes the difference. Front. Microbiol. 2015, 6, 256. [Google Scholar] [CrossRef]
- Riches, A.; Hart, C.; Trenholme, K.; Skinner-Adams, T.S. Anti-Giardia drug discovery: Current status and gut feelings. J. Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Ge, J. Clinical Use of Aspirin in Treatment and Prevention of Cardiovascular Disease. Thrombosis 2012, 2012, 245037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forder, S.; Voelker, M.; Lanas, A. Gastrointestinal Safety of Aspirin for a High-Dose, Multiple-Day Treatment Regimen: A Meta-Analysis of Three Randomized Controlled Trials. Drugs R&D 2016, 16, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Nosala, C.; Hagen, K.D.; Hilton, N.; Chase, T.M.; Jones, K.; Loudermilk, R.; Nguyen, K.; Dawson, S.C. Disc-associated proteins mediate the unusual hyperstability of the ventral disc in Giardia lamblia. J. Cell Sci. 2020, 133, jcs227355. [Google Scholar] [CrossRef]
- Alsop, R.J.; Toppozini, L.; Marquardt, D.; Kucerka, N.; Harroun, T.A.; Rheinstädter, M.C. Aspirin inhibits formation of cholesterol rafts in fluid lipid membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Alsop, R.J.; Barrett, M.A.; Zheng, S.; Dies, H.; Rheinstädter, M.C. Acetylsalicylic acid (ASA) increases the solubility of cholesterol when incorporated in lipid membranes. Soft Matter 2014, 10, 4275–4286. [Google Scholar] [CrossRef] [Green Version]
- Yichoy, M.; Duarte, T.T.; De Chatterjee, A.; Mendez, T.L.; Aguilera, K.Y.; Roy, D.; Roychowdhury, S.; Aley, S.B.; Das, S. Lipid metabolism inGiardia: A post-genomic perspective. Parasitology 2011, 138, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Mendez, T.L.; De Chatterjee, A.; Duarte, T.T.; De Leon, J.; Robles-Martinez, L.; Das, S. Sphingolipids, Lipid Rafts, and Giardial Encystation: The Show Must Go On. Curr. Trop. Med. Rep. 2015, 2, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Efeoğlu, B. Heat shock proteins and heat shock response in plants. Gazi Univ. J. Sci. 2009, 22, 67–75. [Google Scholar]
- Emery, S.J.; Baker, L.; Ansell, B.R.E.; Mirzaei, M.; Haynes, P.A.; McConville, M.J.; Svärd, S.G.; Jex, A.R. Differential protein expression and post-translational modifications in metronidazole-resistant Giardia duodenalis. Gigascience 2018, 7. [Google Scholar] [CrossRef]
- Keister, D.B. Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 487–488. [Google Scholar] [CrossRef]
- Wender, N.; Villalobo, E.; Mirelman, D. EhLimA, a Novel LIM Protein, Localizes to the Plasma Membrane in Entamoeba histolytica. Eukaryot. Cell 2007, 6, 1646–1655. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 on Giardia | CC50 on Caco-2 Cells | Selectivity Index CC50 Caco-2/IC50 Giardia |
|---|---|---|
| 0.29 mM | 2.32 mM | 8 |
| UniProt Access Number | Description | mW (Da) | PLGS Score | Coverage (%) |
|---|---|---|---|---|
| A8BCR6_GIAIC | Cytosolic HSP70 (GL50803_88765) | 71,588 | 5209.85 | 49.4 |
| E2RU36_GIAIC | Arginine deiminase (GL50803_112103) | 64,090 | 1510.245 | 31.4 |
| A8BQS7_GIAIC | HSP90 alpha (GL50803_13864) | 36,966 | 881.7655 | 24.1 |
| E2RU18_GIAIC | Phosphomannomutase 2 (GL50803_17254) | 73,882 | 792.8965 | 35.9 |
| E2RTY6_GIAIC | Glucose 6 phosphate isomerase (GL50803_9115) | 64,451 | 691.5236 | 15.1 |
| A8B2H9_GIAIC | Uncharacterized protein (GL50803_10315) | 25,108 | 627.4312 | 20.3 |
| A8BBL2_GIAIC | Uncharacterized protein (GL50803_27947) | 7457 | 626.1924 | 41.5 |
| A8B431_GIAIC | Bip (GL50803_17121) | 74,314 | 586.4369 | 30.1 |
| A8BUY7_GIAIC | AAA family ATPase (GL50803_16867) | 96,298 | 565.1209 | 27.5 |
| A8B820_GIAC | Protein 21.1 (GL50803_17060) | 66,194 | 554.8513 | 27.6 |
| A8BA49_GIAIC | Uncharacterized protein (GL50803_16507) | 98,575 | 517.2396 | 11.9 |
| Giardia MDRs | Homo Sapiens MDRs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pgp/MDR1 | MRP4 | ABCG2 | |||||||
| Bit Score | Identity (%) | E Value | Bit Score | Identity (%) | E Value | Bit Score | Identity (%) | E Value | |
| giABC | 81.6 | 25 | 9 × 10−16 | ||||||
| giABCP | 171 | 24 | 1 × 10−42 | 360 | 31 | 8 × 10−104 | |||
| giMDRP | 140 | 35 | 3 × 10−33 | ||||||
| giMRPL | 129 | 32 | 2 × 10−29 | 450 | 34 | 6 × 10−133 | |||
| giMDRAP1 | 533 | 35 | 8 × 10−163 | ||||||
| GenBank Accession Number | Gene | Primer (5′-3′) |
|---|---|---|
| XM_001707918.1 | HSP70 | F 5′-CAT CGC CAA TGA GCA GGG CGC GTA-3′ R 5′-ATC GCC CTG TTG CTA CCG GAG A-3′ |
| XM_001706171.1 | Multidrug resistance-associated protein 1 (giMDRAP1) | F 5′-AGA CTC GAG CGA CAA GAA CCC CAA CCA CGT T-3′ R 5′-AGA CTC GAG TGA AGA GCT TGA GGT CGG GTA TC-3′ |
| XM_001705432.1 | ABC transporter family protein (giABC) | F 5′-AGA CTC GAG ACA CGA ATA GGT GGT TAG CCG ACT-3′ R 5′-AGA CTC GAG AGA CTG ACC CAC ATA TGC CCG C-3′ |
| XM_001709457.1 | MRP-like ABC transporter (giMRPL) | F 5′-AGA CTC GAG CAA ATA CAA GTC CAG AGA AGC AGG-3′ R 5′-AGA CTC GAG CAG AGA ACC AGT GTC TGT CAA C-3′ |
| XM_001710125.1 | Multidrug resistance ABC transporter ATP-binding and permease protein (giMDRP) | F 5′-GAG CTC GAG GGT CTA CTT GAG AAG GCC ATT CCA-3′ R 5′-AGA CTC GAG GTC AAC GCT TTT GAA CTT GTG CA-3′ |
| XM_001705989.1 | ABC transporter, putative (giABCP) | F 5′-AGA CTC GAG GGG CTT GCC ACA CTT GTT GGC AGC-3′ R 5′-AGA CTC GAG GAT GCA CTT GAT AGT CAG AGT CGT-3′ |
| XM_001708537.1 | H-shippo 1 | F 5′-CGT CAT CAA CAG GTC CGA-3′ R 5′-CCA GCT CTC CTT GAA CAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochoa-Maganda, V.Y.; Rangel-Castañeda, I.A.; Suárez-Rico, D.O.; Cortés-Zárate, R.; Hernández-Hernández, J.M.; Pérez-Rangel, A.; Chiquete-Félix, N.; León-Ávila, G.; González-Pozos, S.; Gaona-Bernal, J.; et al. Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters. Pharmaceuticals 2020, 13, 440. https://doi.org/10.3390/ph13120440
Ochoa-Maganda VY, Rangel-Castañeda IA, Suárez-Rico DO, Cortés-Zárate R, Hernández-Hernández JM, Pérez-Rangel A, Chiquete-Félix N, León-Ávila G, González-Pozos S, Gaona-Bernal J, et al. Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters. Pharmaceuticals. 2020; 13(12):440. https://doi.org/10.3390/ph13120440
Chicago/Turabian StyleOchoa-Maganda, Verónica Yadira, Itzia Azucena Rangel-Castañeda, Daniel Osmar Suárez-Rico, Rafael Cortés-Zárate, José Manuel Hernández-Hernández, Armando Pérez-Rangel, Natalia Chiquete-Félix, Gloria León-Ávila, Sirenia González-Pozos, Jorge Gaona-Bernal, and et al. 2020. "Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters" Pharmaceuticals 13, no. 12: 440. https://doi.org/10.3390/ph13120440
APA StyleOchoa-Maganda, V. Y., Rangel-Castañeda, I. A., Suárez-Rico, D. O., Cortés-Zárate, R., Hernández-Hernández, J. M., Pérez-Rangel, A., Chiquete-Félix, N., León-Ávila, G., González-Pozos, S., Gaona-Bernal, J., & Castillo-Romero, A. (2020). Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters. Pharmaceuticals, 13(12), 440. https://doi.org/10.3390/ph13120440