
Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives

 , ,
, , 
Abstract
:1. Introduction
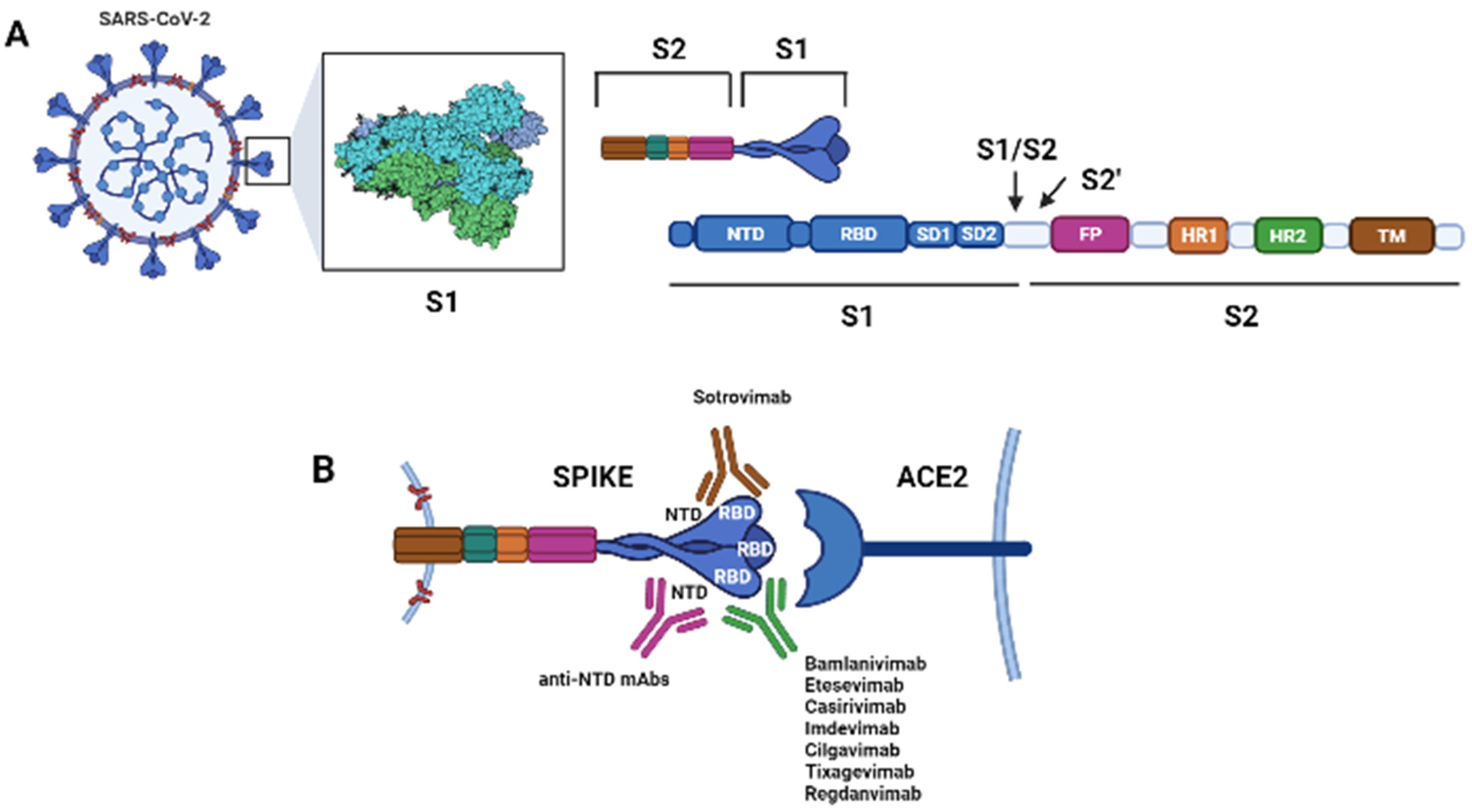
2. Antibodies Targeting SARS-CoV-2
2.1. Principal mAbs against SARS-CoV-2
2.1.1. Bamlanivimab
2.1.2. Bamlanivimab and Etesevimab
2.1.3. Casirivimab and Imdevimab
2.1.4. Sotrovimab
2.1.5. Cilgavimab and Tixagevimab
2.1.6. Regdanvimab
2.2. MAbs Approved against SARS-CoV-2 by Emergency Use Authorizations (EUAs)
- Bamlanivimab 700 mg plus etesevimab 1400 mg (intravenous infusion, (iv));
- Casirivimab 600 mg plus imdevimab 600 mg (iv or subcutaneous injection);
- Sotrovimab 500 mg (iv).
3. Therapeutic and Prophylactic Indications
3.1. Prophylactic Use of mAb against SARS-CoV-2
3.2. Use of mAb in Patient Hospitalized for COVID-19
4. Challenges for Using mAbs against SARS-CoV-2
4.1. Activity against SARS-CoV-2 Variants
- Beta (B.1.351) variant: This VoC includes the E484K and K417N mutations, which results in a reduction in in vitro susceptibility to bamlanivimab and etesevimab [36,65]. In vitro studies also suggest that this variant has markedly reduced susceptibility to casirivimab, although the combination of casirivimab and imdevimab appears to retain activity; sotrovimab appears active as well against this VoC [38,39].
- Gamma (P.1) variant: This VoC includes the E484K and K417T mutations, which results in a marked reduction in in vitro susceptibility to bamlanivimab and etesevimab [36,66]. Additionally, this variant shows reduced susceptibility to casirivimab, although the combination of casirivimab and imdevimab appears to retain activity; sotrovimab appears to retain activity as well [38,39].
- Delta (B.1.617.2) variant: This is the prevalent VoC in the United States. It contains the L452R mutation, which results in a modest decrease in in vitro susceptibility to the combination of bamlanivimab and etesevimab, although the clinical implications of this finding are not fully known. Sotrovimab and casirivimab plus imdevimab appear to maintain activity [38,39,67].
4.2. Innovative Way of mAbs Administration
5. Global Access
6. Conclusions
7. Research Strategy and Selection Criteria
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marston, H.D.; Paules, C.I.; Fauci, A.S. Monoclonal Antibodies for Emerging Infectious Diseases—Borrowing from History. N. Engl. J. Med. 2018, 378, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Guthmiller, J.J.; Dugan, H.L.; Neu, K.E.; Lan, L.Y.; Wilson, P.C. An Efficient Method to Generate Monoclonal Antibodies from Human B Cells. Methods Mol. Biol. 2019, 1904, 109–145. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.; Grzeschik, J.; Hinz, S.C.; Becker, S.; Toleikis, L.; Busch, M.; Kolmar, H.; Krah, S.; Zielonka, S. Facile generation of antibody heavy and light chain diversities for yeast surface display by Golden Gate Cloning. Biol. Chem. 2019, 400, 383–393. [Google Scholar] [CrossRef]
- Burioni, R.; Lang, A.B.; Capra, J.D. Human monoclonal antibodies as a new class of antiinfective compounds. Clin. Dev. Immunol. 2013, 2013, 297120. [Google Scholar] [CrossRef] [Green Version]
- Pelfrene, E.; Mura, M.; Cavaleiro Sanches, A.; Cavaleri, M. Monoclonal antibodies as anti-infective products: A promising future? Clin. Microbiol. Infect. 2019, 25, 60–64. [Google Scholar] [CrossRef]
- IMpact-RSV-Study-Group. Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. Pediatrics 1998, 102, 531–537. [Google Scholar] [CrossRef]
- Zhu, Q.; Patel, N.K.; McAuliffe, J.M.; Zhu, W.; Wachter, L.; McCarthy, M.P.; Suzich, J.A. Natural polymorphisms and resistance-associated mutations in the fusion protein of respiratory syncytial virus (RSV): Effects on RSV susceptibility to palivizumab. J. Infect. Dis. 2012, 205, 635–638. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency; Committee for Medicinal Products for Human Use. ZinplavadEPAR Summary for the Public (EMA/201086/2017). Available online: http://www.ema.europa.eu/docs/en_GB/document_library/EPAR__Summary_for_the_public/human/004136/WC500222645.pdf (accessed on 15 November 2021).
- U.S. Food and Drug Administration. FDA Approved Drug Products: Raxibacumab. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2012/125349s000lbl.pdf (accessed on 15 November 2021).
- U.S. Food and Drug Administration. FDA Approved Drug Products: Anthim. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2016/125509lbl.pdf (accessed on 15 November 2021).
- U.S. Food and Drug Administration. FDA Approved Drug Products: Trogarzo. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/761065lbl.pdf (accessed on 15 November 2021).
- Mulangu, S.; Dodd, L.E.; Davey, R.T.; Tshiani Mbaya, O.; Proschan, M.; Mukadi, D.; Lusakibanza Manzo, M.; Nzolo, D.; Tshomba Oloma, A.; Ibanda, A.; et al. A randomized, controlled trial of Ebola virus disease therapeutics. N. Engl. J. Med. 2019, 381, 2293–2303. [Google Scholar] [CrossRef]
- Callaway, B.E.; Cyranoski, D.; Mallapaty, S.; Stoye, E.; Tollefson, J. Coronavirus by the numbers. Nature 2020, 579, 482–483. [Google Scholar] [CrossRef] [Green Version]
- Kreuzberger, N.; Hirsch, C.; Chai, K.L.; Tomlinson, E.; Khosravi, Z.; Popp, M.; Neidhardt, M.; Piechotta, V.; Salomon, S.; Valk, S.J.; et al. SARS-CoV-2-neutralising monoclonal antibodies for treatment of COVID-19. Cochrane Database Syst. Rev. 2021, 9, CD013825. [Google Scholar] [CrossRef]
- Kumar, D.; Gauthami, S.; Bayry, J.; Kaveri, S.V.; Hegde, N.R. Antibody Therapy: From Diphtheria to Cancer, COVID-19, and Beyond. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 36–49. [Google Scholar] [CrossRef]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; Van Haperen, R.; Osterhaus, A.; Van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.J. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, R.; Pan, Z.; Qian, C.; Yang, Y.; You, R.; Zhao, J.; Liu, P.; Gao, L.; Li, Z.; et al. Human monoclonal antibodies block the binding of SARS-CoV-2 spike protein to angiotensin converting enzyme 2 receptor. Cell Mol. Immunol. 2020, 17, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Noy-Porat, T.; Makdasi, E.; Alcalay, R.; Mechaly, A.; Levy, Y.; Bercovich-Kinori, A.; Zauberman, A.; Tamir, H.; Yahalom-Ronen, Y.; Israeli, M.; et al. A panel of human neutralizing mAbs targeting SARSCoV-2 spike at multiple epitopes. Nat. Commun. 2020, 11, 4303. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Lv, Z.; Deng, Y.-Q.; Ye, Q.; Cao, L.; Sun, C.-Y.; Fan, C.; Huang, W.; Sun, S.; Sun, Y.; Zhu, L.; et al. Structural basis for neutralization of SARSCoV-2 and SARS-CoV by a potent therapeutic antibody. Science 2020, 369, 1505–1509. [Google Scholar] [CrossRef]
- Iyer, A.S.; Jones, F.K.; Nodoushani, A.; Kelly, M.; Becker, M.; Slater, D.; Mills, R.; Teng, E.; Kamruzzaman, M.; Garcia-Beltran, W.F.; et al. Persistence and decay of human antibody responses to the receptor binding domain of SARS-CoV-2 spike protein in COVID-19 patients. Sci. Immunol. 2020, 5, eabe0367. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Abernathy, M.E.; Dam, K.A.; Esswein, S.R.; Jette, C.A.; Bjorkman, P.J. How Antibodies Recognize Pathogenic Viruses: Structural Correlates of Antibody Neutralization of HIV-1, SARS-CoV-2, and Zika. Viruses 2021, 13, 2106. [Google Scholar] [CrossRef]
- Winkler, E.S.; Gilchuk, P.; Yu, J.; Bailey, A.L.; Chen, R.E.; Zost, S.J.; Jang, H.; Huang, Y.; Allen, J.D.; Case, J.B.; et al. Human neutralizing antibodies against SARS-CoV-2 require intact Fc effector functions for optimal therapeutic protection. Cell 2021, 184, 1804–1820. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond binding: Antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Gupta, A.; Ravetch, J.V. The role of IgG Fc receptors in antibody-dependent enhancement. Nat. Rev. Immunol. 2020, 20, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Cao, Y. In Translation: FcRn across the Therapeutic Spectrum. Int. J. Mol. Sci. 2021, 22, 3048. [Google Scholar] [CrossRef] [PubMed]
- Bamlanivimab U.S. Food and Drug Administration. Fact Sheet for Health Care Providers: Emergency Use Authorization (EUA) of Bamlanivimab. Available online: https://www.fda.gov/media/143603/download (accessed on 15 November 2021).
- U.S. Food and Drug Administration. Coronavirus (COVID-19) Update: FDA Revokes Emergency Use Authorization for Monoclonal Antibody Bamlanivimab. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-revokes-emergency-use-authorization-monoclonal-antibody-bamlanivimab (accessed on 15 November 2021).
- Banlanivimab and Etesevimab EUA. Available online: https://www.covid19.lilly.com/bam-ete (accessed on 15 November 2021).
- U.S. Food and Drug Administration. Fact Sheet for Health Care Providers: Emergency Use Authorization (EUA) of Bamlanivimab and Etesevimab. Available online: https://www.fda.gov/media/145802/download (accessed on 15 November 2021).
- Casirivimab and Imdevimab. Available online: https://www.regeneron.com/medicines/casirivimab-imdevimab (accessed on 15 November 2021).
- U.S. Food and Drug Administration. Fact Sheet for Health Care Providers: Emergency Use Authorization (EUA) of Casirivimab and Imdevimab. Available online: https://www.fda.gov/media/145611/download (accessed on 15 November 2021).
- U.S. Food and Drug Administration. Frequently Asked Questions on the Emergency Use Authorization of Sotrovimab. Available online: https://www.fda.gov/media/149534/download (accessed on 15 November 2021).
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.E.; Nkolola, J.P.; Schäfer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef]
- Zost, S.J.; Gilchuk, P.; Chen, R.E.; Case, J.B.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; Sutton, R.E.; Suryadevara, N.; Chen, E.C.; et al. Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. Nat. Med. 2020, 26, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zost, S.J.; Greaney, A.J.; Starr, T.N.; Dingens, A.S.; Chen, E.C.; Chen, R.E.; Case, J.B.; Sutton, R.E.; Gilchuk, P.; et al. Genetic and structural basis for SARS-CoV-2 variant neutralization by a two-antibody cocktail. Nat. Microbiol. 2021, 6, 1233–1244. [Google Scholar] [CrossRef]
- Kim, C.; Ryu, D.K.; Lee, J.; Kim, Y.I.; Seo, J.M.; Kim, Y.G.; Jeong, J.H.; Kim, M.; Kim, J.I.; Kim, P.; et al. A therapeutic neutralizing antibody targeting receptor binding domain of SARS-CoV-2 spike protein. Nat. Commun. 2021, 12, 288. [Google Scholar] [CrossRef]
- Celltrion’s Regdanvimab (CT-P59) Becomes the First Authorized COVID-19 Treatment Approved from the Korean MFDS. Available online: https://www.celltrionhealthcare.com/en-us/board/newsdetail?modify_key=526&pagenumber=1&keyword=&keyword_type= (accessed on 15 November 2021).
- EMA Receives Application for Marketing Authorisation for Regkirona (regdanvimab) for Treating Patients with COVID-19. Available online: https://www.ema.europa.eu/en/news/ema-receives-application-marketing-authorisation-regkirona-regdanvimab-treating-patients-covid-19 (accessed on 15 November 2021).
- Lilly, Vir Biotechnology and GSK Announce Positive Topline Data from the Phase 2 BLAZE-4 Trial Evaluating Bamlanivimab with VIR-7831 in Low-Risk Adults with COVID-19. Available online: https://investor.lilly.com/news-releases/news-release-details/lilly-vir-biotechnology-and-gsk-announce-positive-topline-data (accessed on 15 November 2021).
- Aleem, A.; Slenker, A.K. Monoclonal Antibody Therapy For High-Risk Coronavirus (COVID 19) Patients with mild to Moderate Disease Presentations; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Dougan, M.; Nirula, A.; Azizad, M.; Mocherla, B.; Gottlieb, R.L.; Chen, P.; Hebert, C.; Perry, R.; Boscia, J.; Helleret, B.; et al. Bamlanivimab plus etesevimab in mild or moderate COVID-19. N. Engl. J. Med. 2021, 385, 1382–1392, published online ahead of print. Available online: https://www.ncbi.nlm.nih.gov/pubmed/34260849 (accessed on 15 November 2021). [CrossRef]
- Regeneron. COV-2067 Phase 3 Trial in High-Risk Outpatients Shows that REGEN-COV (2400 mg and 1200 mg IV doses) Significantly Reduces Risk of Hospitalization or Death While Also Shortening Symptom Duration. 2021. Available online: https://newsroom.regeneron.com/index.php/static-files/a7173b5a-28f3-45d4-bede-b97370bd03f8 (accessed on 15 November 2021).
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Gonzalez-Rojas, Y.; Juarez, E.; Crespo Casal, M.; Moya, J.; Falci, D.R.; Sarkis, E.; Solis, J.; Zheng, H.; Scott, N.; et al. Early Treatment for Covid-19 with SARS-CoV-2 Neutralizing Antibody Sotrovimab. N. Engl. J. Med. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Vladislav, I.; Matlock, A.J.; Sawalha, A.H. COVID-19: A review of therapeutic strategies and vaccine candidates. Clin. Immunol. 2021, 222, 108634. [Google Scholar] [CrossRef]
- Corey, L.; Mascola, J.R.; Fauci, A.S.; Collins, F.S. A strategic approach to COVID-19 vaccine R&D. Science 2020, 368, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Wheatley, A.K. Neutralizing Antibody Therapeutics for COVID-19. Viruses 2021, 13, 628. [Google Scholar] [CrossRef] [PubMed]
- Marovich, M.; Mascola, J.R.; Cohen, M.S. Monoclonal Antibodies for Prevention and Treatment of COVID-19. JAMA 2020, 324, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Nirula, A.; Mulligan, M.J.; Novak, R.M.; Marovich, M.; Yen, C.; Stemer, A.; Mayer, S.M.; Wohl, D.; Brengle, B.; et al. Effect of Bamlanivimab vs Placebo on Incidence of COVID-19 Among Residents and Staff of Skilled Nursing and Assisted Living Facilities: A Randomized Clinical Trial. JAMA 2021, 326, 46–55. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.P.; Forleo-Neto, E.; Musser, B.J.; Isa, F.; Chan, K.C.; Sarkar, N.; Bar, K.J.; Barnabas, R.V.; Barouch, D.H.; Cohen, M.S.; et al. Subcutaneous REGEN-COV Antibody Combination to Prevent Covid-19. N. Engl. J. Med. 2021, 385, 1184–1195. [Google Scholar] [CrossRef] [PubMed]
- CDC Quarantine and Isolation. Available online: https://www.cdc.gov/coronavirus/2019-ncov/your-health/quarantine-isolation.html?CDC_AA_refVal=https%3A%2F%2Fwww.cdc.gov%2Fcoronavirus%2F2019-ncov%2Fif-you-are-sick%2Fquarantine.html (accessed on 15 November 2021).
- Update on AZD7442 STORM CHASER Trial in Post-Exposure Prevention of Symptomatic COVID-19. Available online: https://www.astrazeneca.com/media-centre/press-releases/2021/update-on-azd7442-storm-chaser-trial.html (accessed on 15 November 2021).
- AZD7442 PROVENT Phase III Prophylaxis Trial Met Primary Endpoint in Preventing COVID-19. Available online: https://www.astrazeneca.com/media-centre/press-releases/2021/azd7442-prophylaxis-trial-met-primary-endpoint.html (accessed on 15 November 2021).
- RECOVERY Collaborative Group. Casirivimab and imdevimab in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. medRxiv. 2021. preprint. Available online: https://www.medrxiv.org/content/10.1101/2021.06.15.21258542v1.full (accessed on 15 November 2021).
- Focosi, D.; Maggi, F. Neutralising antibody escape of SARS-CoV-2 spike protein: Risk assessment for antibody-based COVID-19 therapeutics and vaccines. Rev. Med. Virol. 2021, 1–21. [Google Scholar] [CrossRef]
- Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants (accessed on 15 November 2021).
- FDA (US Food and Drug Administration). Development of Monoclonal Antibody Products Targeting SARS-CoV-2, Including Addressing the Impact of Emerging Variants, During the COVID 19 Public Health Emergency. 2021. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/development-monoclonal-antibody-products-targeting-sars-cov-2-includingaddressing-impact-emerging (accessed on 15 November 2021).
- Ho, D.; Wang, P.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.; et al. Increased Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7 to Antibody Neutralization. Res. Sq. 2021, 29, 747–751. [Google Scholar] [CrossRef]
- Wang, P.; Wang, P.; Casner, R.G.; Nair, M.S.; Wang, M.; Yu, J.; Cerutti, G.; Liu, L.; Kwong, P.D.; Huang, Y.; et al. Increased resistance of SARS-CoV-2 variant P.1 to antibody neutralization. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ginn, H.M.; Dejnirattisai, W.; Supasa, P.; Wang, B.; Tuekprakhon, A.; Nutalai, R.; Zhou, D.; Mentzer, A.J.; Zhao, Y.; et al. Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum. Cell 2021, 184, 4220–4236.e13, published online ahead of print. Available online: https://www.ncbi.nlm.nih.gov/pubmed/34242578 (accessed on 15 November 2021). [CrossRef]
- Vellas, C.; Del Bello, A.; Debard, A.; Steinmeyer, Z.; Tribaudeau, L.; Ranger, N.; Jeanne, N.; Martin-Blondel, G.; Delobel, P.; Kamar, N.; et al. Influence of treatment with neutralizing monoclonal antibodies on the SARS-CoV-2 nasopharyngeal load and quasispecies. Clin. Microbiol. Infect. 2021. [Google Scholar] [CrossRef]
- Corti, D.; Purcell, L.A.; Snell, G.; Veesler, D. Tackling COVID-19 with neutralizing monoclonal antibodies. Cell 2021, 184, 3086–3108. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Xiang, J.; Cui, Z.; Liu, J.; Liu, C. Intranasal administration of SARS-CoV-2 neutralizing human antibody prevents infection in mice. bioRxiv 2020. [Google Scholar] [CrossRef]
- Nishat, F.; Vichitra, K.; Amjad, A. A Narrative Review of a Pulmonary Aerosolized Formulation or a Nasal Drop Using Sera Containing Neutralizing Antibodies Collected from COVID-19-Recovered Patients as a Probable Therapy for COVID-19. Iran. J. Med. Sci. 2021, 46, 151–168. [Google Scholar] [CrossRef]
- Vivek, P.C.; Lalitkumar, K.V.; Anjali, K.P.; Vandana, B.P. Intranasal vaccines for SARS-CoV-2: From challenges to potential in COVID-19 management. Drug Discov. Today 2021, 26, 2619–2636. [Google Scholar] [CrossRef]
- Tiboni, M.; Casettari, L.; Illum, L. Nasal vaccination against SARS-CoV-2: Synergistic or alternative to intramuscular vaccines? Int. J. Pharm. 2021, 603, 120686. [Google Scholar] [CrossRef]
- Du, Y.; Xu, Y.; Feng, J.; Hu, L.; Zhang, Y.; Zhang, B.; Guo, W.; Mai, R.; Chen, L.; Fang, J.; et al. Intranasal administration of a recombinant RBD vaccine induced protective immunity against SARS-CoV-2 in mouse. Vaccine 2021, 39, 2280–2287. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.D.; Schmitz, K.S.; Bovier, F.T.; Predella, C.; Khao, J.; Noack, D.; Haagmans, B.L.; Herfst, S.; Stearns, K.N.; Drew-Bear, J.; et al. Intranasal fusion inhibitory lipopeptide prevents direct contact SARSCoV-2 transmission in ferrets. Science 2021, 371, 1379–1382. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Sun, S.; Tai, W.; Chen, J.; Geng, Q.; He, L.; Chen, Y.; Wu, J.; Shi, Z.; et al. Molecular mechanism for antibody-dependent enhancement of coronavirus entry. J. Virol. 2020, 94, e02015-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parray, H.A.; Shukla, S.; Perween, R.; Khatri, R.; Shrivastava, T.; Singh, V.; Murugavelu, P.; Ahmed, S.; Samal, S.; Sharma, C.; et al. Inhalation monoclonal antibody therapy: A new way to treat and manage respiratory infections. Appl. Microbiol. Biotechnol. 2021, 105, 6315–6332. [Google Scholar] [CrossRef] [PubMed]
- Piepenbrink, M.S.; Park, J.G.; Oladunni, F.S.; Deshpande, A.; Basu, M.; Sarkar, S.; Loos, A.; Woo, J.; Lovalenti, P.; Sloan, D.; et al. Therapeutic activity of an inhaled potent SARS-CoV-2 neutralizing human monoclonal antibody in hamsters. Cell Rep. Med. 2021, 2, 100218. [Google Scholar] [CrossRef]
- In-patient COVID-19 Study of Intranasal Foralumab. Available online: https://clinicaltrials.gov/ct2/show/record/NCT04983446?term=foralumab&cond=COVID-19&draw=2&rank=1 (accessed on 15 November 2021).
- Valdez-Cruz, N.A.; García-Hernández, E.; Espitia, C.; Cobos-Marín, L.; Altamirano, C.; Bando-Campos, C.G.; Cofas-Vargas, L.F.; Coronado-Aceves, E.W.; González-Hernández, R.A.; Hernández-Peralta, P.; et al. Integrative overview of antibodies against SARS-CoV-2 and their possible applications in COVID-19 prophylaxis and treatment. Microb. Cell Fact. 2021, 20, 88. [Google Scholar] [CrossRef]
- Kaplon, H.; Reichert, J.M. Antibodies to Watch in 2021. MAbs 2021, 13, 1860476. [Google Scholar] [CrossRef]
- Jaworski, J.P. Neutralizing monoclonal antibodies for COVID-19 treatment and prevention. Biomed. J. 2021, 44, 7–17. [Google Scholar] [CrossRef]
- Wellcome Trust. Expanding Access to Monoclonal Antibody-Based Products. Available online: https://wellcome.org/sites/default/files/expanding-access-to-monoclonal-antibody-based-products.pdf (accessed on 15 November 2021).
- Tuccori, M.; Ferraro, S.; Convertino, I.; Cappello, E.; Valdiserra, G.; Blandizzi, C.; Maggi, F.; Focosi, D. Anti-SARS-CoV-2 neutralizing monoclonal antibodies: Clinical pipeline. MAbs 2020, 12, 1854149. [Google Scholar] [CrossRef]
- CDC COVID Data Tracker. Available online: https://www.cdc.gov/coronavirus/2019-ncov/covid-data/covidview/index.html (accessed on 15 November 2021).
- WHO Coronavirus (COVID-19) Dashboard|WHO Coronavirus (COVID-19) Dashboard with Vaccination Data. Available online: https://covid19.who.int (accessed on 15 November 2021).


{kind=link}
| Age ≥ 65 years |
| Obesity or being overweight (e.g., adults with BMI > 25 kg/m2, or if age 12–17, have BMI ≥85th percentile for their age and gender based on CDC growth charts) |
| Pregnancy |
| Chronic kidney diseases |
| Diabetes |
| Immunosuppressive diseases or immunosuppressive treatment |
| Cardiovascular disease (including congenital heart disease) or hypertension |
| Chronic lung diseases (e.g., chronic obstructive pulmonary disease, asthma, interstitial lung disease, cystic fibrosis and pulmonary hypertension) |
| Sickle cell disease |
| Neurodevelopmental disorders (e.g., cerebral palsy) or other conditions that confer medical complexity (for example, genetic or metabolic syndromes and severe congenital anomalies) |
| Medical-related technological dependence (e.g., tracheostomy, gastrostomy, or positive pressure ventilation (not related to COVID-19)) |
| mAb | Study Design | Methods | Results |
|---|---|---|---|
| Bamlanivimab plus etesevimab (Trial Number NCT04427501) | Double-blind, phase 3 randomized clinical trial in outpatients with mild to moderate COVID-19 who are at high risk for progressing to severe COVID-19 and/or hospitalization [48]. | Intervention: Single intravenous infusion of: - bamlanivimab 2800 mg + etesevimab 2800 mg - Placebo Primary endpoint: Proportion of participants with COVID-19 related hospitalization or death by any cause by day 29 | Number of Participants: - bamlanivimab + etesevimab (n = 518) - placebo (n = 517) Primary outcome: Proportion of participants with COVID-19 related hospitalization or death by any cause by Day 29: 11 of 518 participants (2.1%) in the bamlanivimab + etesevimab arm vs. 36 of 517 (7.0%) in the placebo arm (absolute risk difference, −4.8 percentage points; 95% confidence interval (CI), −7.4 to −2.3; relative risk difference, 70%; p < 0.001). Proportion of participants who had died from any cause by Day 29: 0 of 518 participants (0%) in the bamlanivimab + etesevimab arm vs. 10 of 517 (1.9%) in the placebo arm (p< 0.001). |
| Casirivimab plus imdevimab (Trial Number NCT04425629) | Double-blind, Phase 3 RCT in outpatients with mild to moderate COVID-19 [49,50]. | Intervention: Single intravenous infusion of: - casirivimab 600 mg + imdevimab 600 mg - casirivimab 1200 mg + imdevimab 1200 mg - placebo Endpoint: Proportion of patients with COVID-19-related hospitalization or all-cause death through Day 29 | Number of participants: - casirivimab 600 mg + imdevimab 600 mg (n = 736) vs. placebo (n = 748) - casirivimab 1200 mg + imdevimab 1200 mg (n = 1355) vs. placebo (n = 1341) Primary outcomes: COVID-19-related hospitalization or all-cause death through Day 29: - 7 of 736 (1.0%) in casirivimab 600 mg plus imdevimab 600 mg arm vs. 24 of 748 (3.2%) in placebo arm (relative risk reduction, 70.4%; 95% CI, 31.6 to 87.1; p = 0.0024) - 18 of 1355 (1.3%) in casirivimab 1200 mg plus imdevimab 1200 mg arm vs. 62 of 1341 (4.6%) in placebo arm (relative risk reduction 73%; CI 95%, 51.7 to 82.9; p < 0.001). |
| Sotrovimab (Trial number NCT04545060) | Double-blind, Phase 1/2/3 RCT in outpatients with mild to moderate COVID-19 [51]. | Interventions: - sotrovimab 500 mg IV - Placebo Primary Endpoint: Proportion of patients with hospitalization or death from any cause by Day 29 | Number of Participants: - sotrovimab (n = 291) - placebo (n = 292) Primary outcome: There an 85% relative risk reduction in all-cause hospitalizations or deaths in patients who received sotrovimab compared to those who received placebo. (All-cause hospitalization or death by Day 29: 3 of 291 (1%) in sotrovimab arm vs. 21 of 292 (7%) in placebo arm (p = 0.002)) |
| WHO Label | Pango Lineage * | GISAID Clade | Nextstrain Clade | Additional Aamino Acid Changes Monitored ° | Earliest Documented Samples | Date of Designation |
|---|---|---|---|---|---|---|
| Alpha | B.1.1.7 # | GRY | 20I (V1) | +S:484K +S:452R | United Kingdom, Sep-2020 | 18 December 2020 |
| Beta | B.1.351 | GH/501Y.V2 | 20H (V2) | +S:L18F | South Africa, May-2020 | 18 December 2020 |
| Gamma | P.1 | GR/501Y.V3 | 20J (V3) | +S:681H | Brazil, Nov-2020 | 11 January 2021 |
| Delta | B.1.617.2 § | G/478K.V1 | 21A, 21I, 21J | +S:417N | India, Oct-2020 | VOI: 4 April 2021 VOC: 11 May 2021 |
| WHO Label § | Pango Lineage * | GISAID Clade | Nextstrain Clade | Earliest Documented Samples | Date of Designation |
|---|---|---|---|---|---|
| Lambda | C.37 | GR/452Q.V1 | 21G | Peru, December 2020 | 14 June 2021 |
| Mu | B.1.621 | GH | 21H | Colombia, January 2021 | 30 August 2021 |
| Casirivimab Indevimab | Bamlanivimab Etesevimab | Sotrovimab | Cilgavimab Tixagevimab | Regdanvimab | |
|---|---|---|---|---|---|
| B.1.1.7 (UK) | S S | S S | S | S S | S |
| B.1.351 (South Africa) | R S | R R | S | S S | I/R |
| P.1 (Brazil) | R S | R R | S | S S | Pot I/R |
| B.1.429 (California) | S S | R S | S | S S | I/R |
| B.1.1.258 (Scotland) | S R | S U | S | U U | Pot S |
| B.1.525 (Nigeria) | Pot I/R Pot S | Pot I/R Pot S | S | Pot S Pot S | U |
| B.1.526 (New York) | Pot I/R Pot S | Pot I/R Pot S | S | Pot S Pot S | U |
| B.1.617.1 (India) | S S | R S | S | Pot S Pot S | U |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiros-Roldan, E.; Amadasi, S.; Zanella, I.; Degli Antoni, M.; Storti, S.; Tiecco, G.; Castelli, F. Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives. Pharmaceuticals 2021, 14, 1272. https://doi.org/10.3390/ph14121272
Quiros-Roldan E, Amadasi S, Zanella I, Degli Antoni M, Storti S, Tiecco G, Castelli F. Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives. Pharmaceuticals. 2021; 14(12):1272. https://doi.org/10.3390/ph14121272
Chicago/Turabian StyleQuiros-Roldan, Eugenia, Silvia Amadasi, Isabella Zanella, Melania Degli Antoni, Samuele Storti, Giorgio Tiecco, and Francesco Castelli. 2021. "Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives" Pharmaceuticals 14, no. 12: 1272. https://doi.org/10.3390/ph14121272
APA StyleQuiros-Roldan, E., Amadasi, S., Zanella, I., Degli Antoni, M., Storti, S., Tiecco, G., & Castelli, F. (2021). Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives. Pharmaceuticals, 14(12), 1272. https://doi.org/10.3390/ph14121272