
Etanercept Ameliorates Cardiac Fibrosis in Rats with Diet-Induced Obesity


 and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Etanercept on Changes in Body Weight, Blood Lipids Levels and other Parameters in HFD-Fed Rats
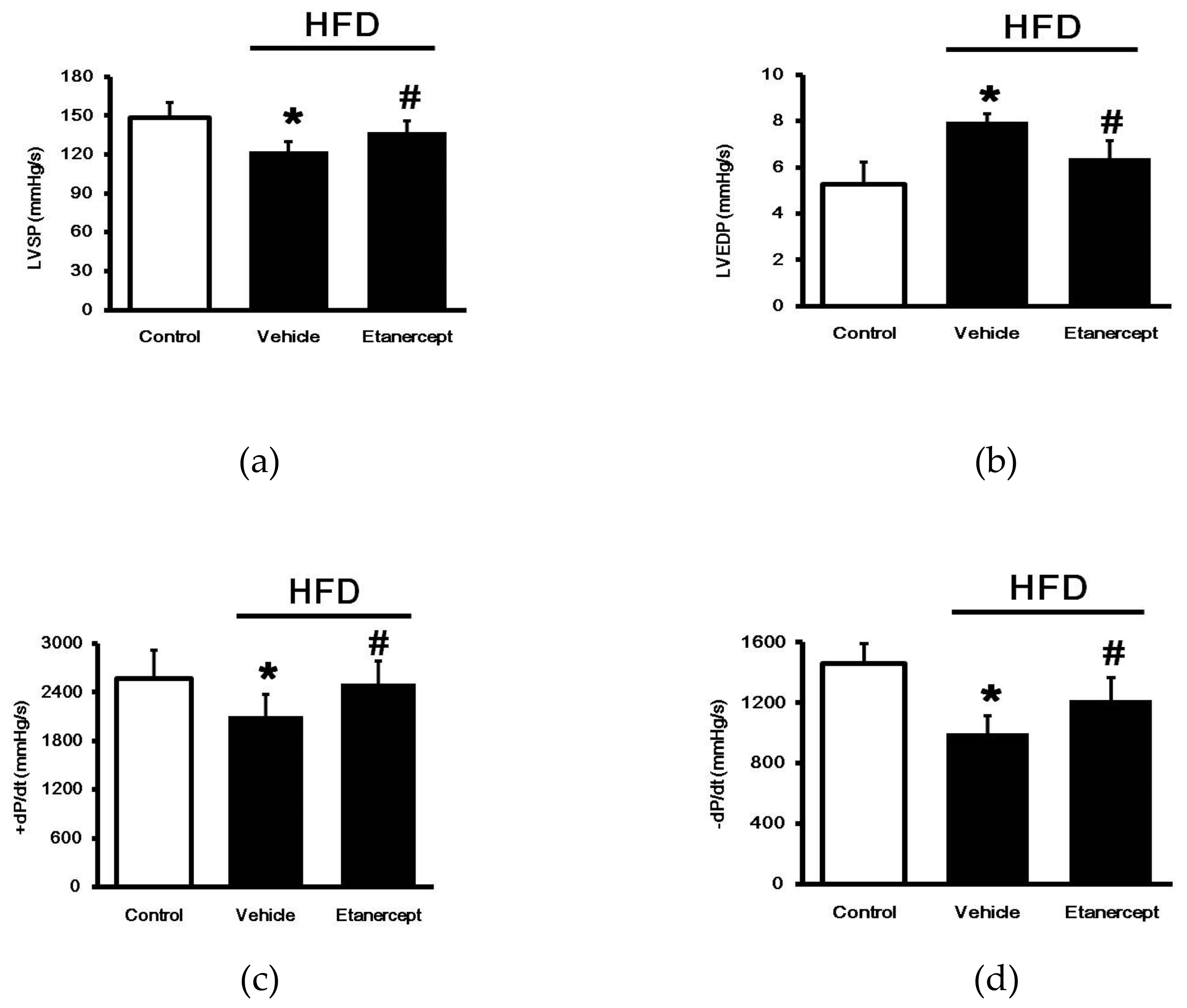
2.2. Effects of Etanercept on Cardiac Functions
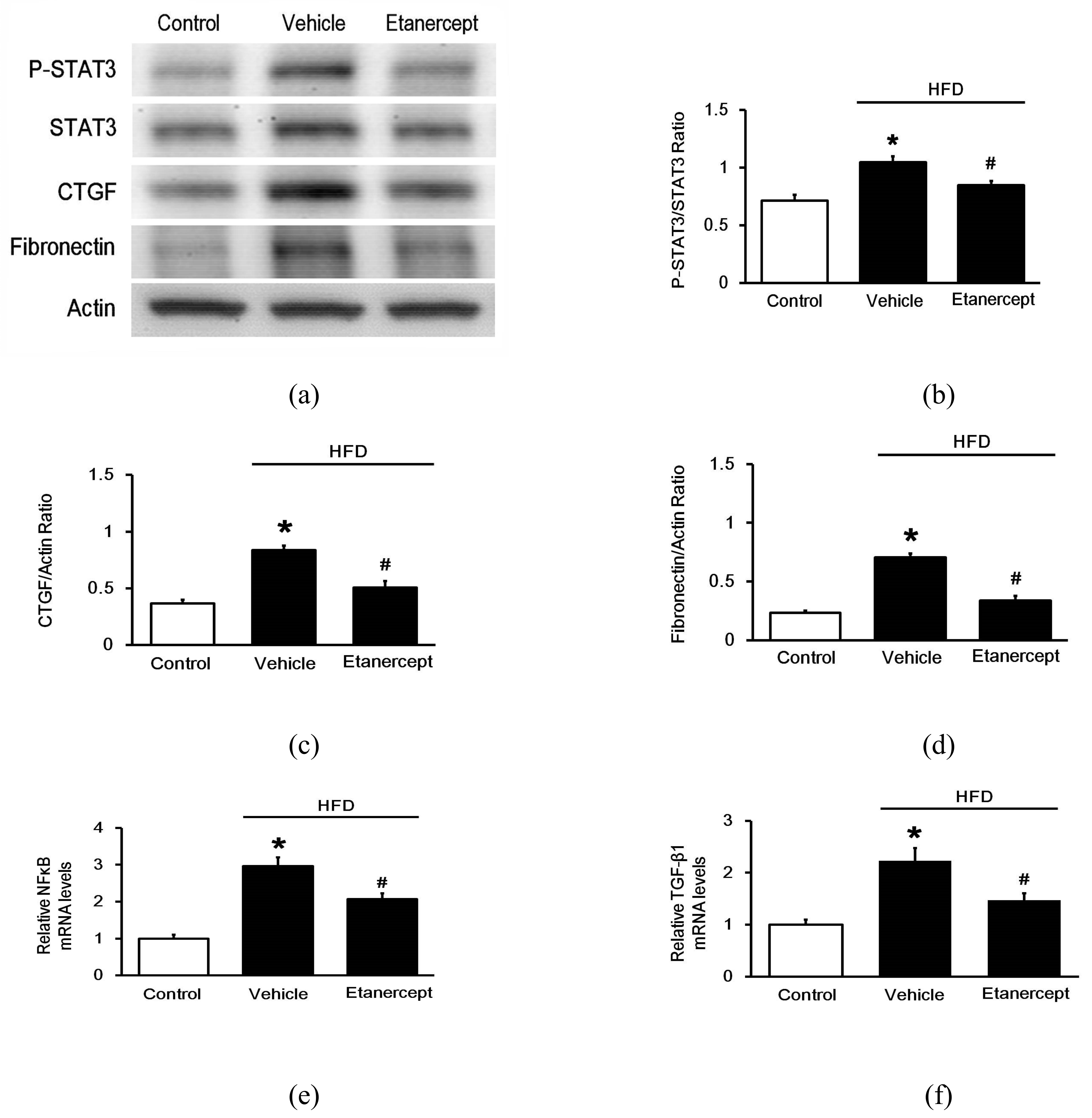
2.3. Effects of Etanercept on Cardiac Fibrosis in HFD-Fed Rats
2.4. Effects of Etanercept on Fibrosis-Related Genes in the Hearts of HFD-Fed Rats
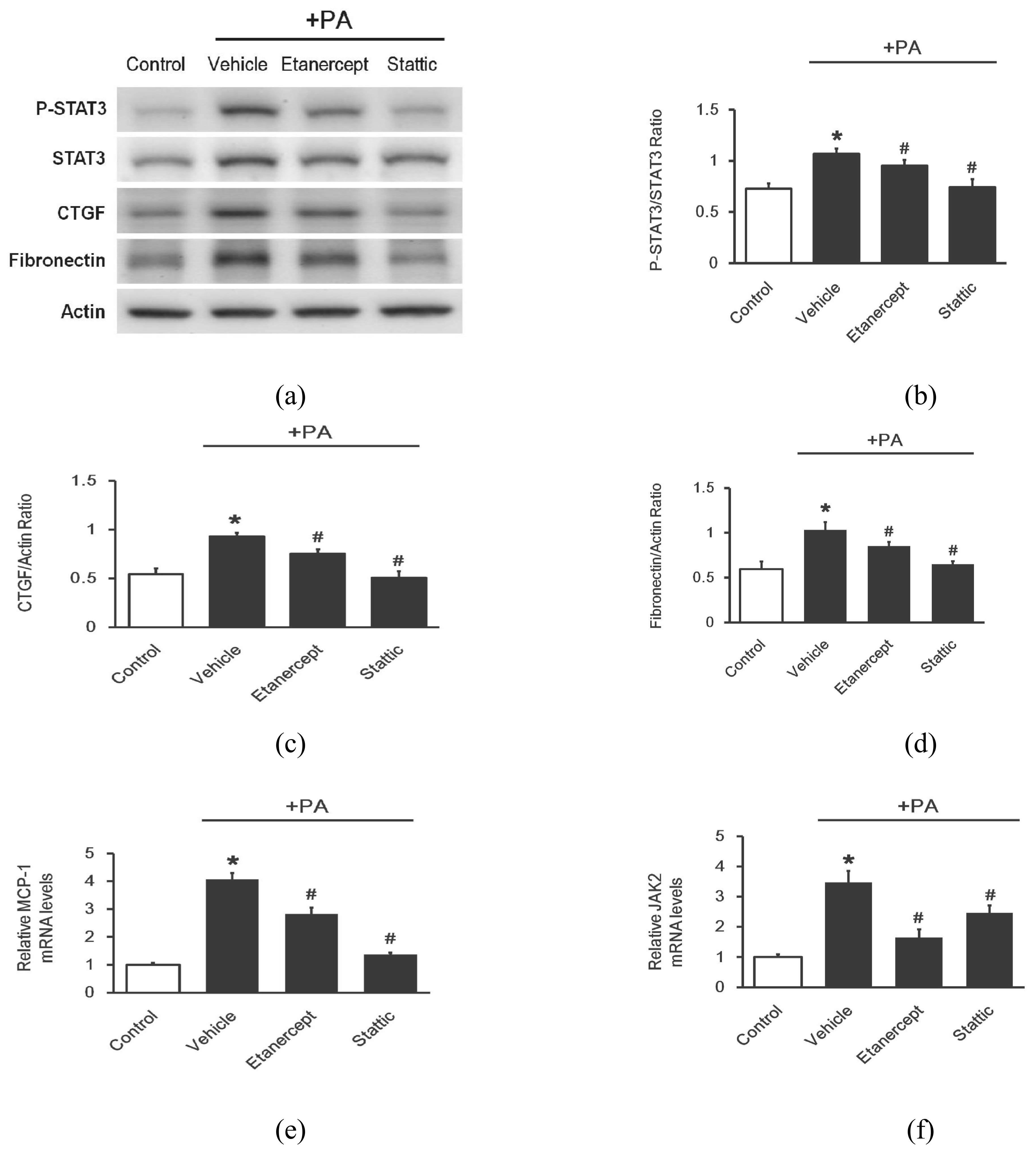
2.5. Effects of Etanercept on Palmitate-Stimulated Fibrosis-Related Gene Expression in Cultured Cardio Myoblast H9c2 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Model
4.3. Analysis of Plasma Lipids
4.4. Measurement of Blood Biomarkers
4.5. Cardiac Performance Measurements
4.6. Morphological Evaluation
4.7. Cell Culture
4.8. Real-Time Quantitative PCR
4.9. Western Blotting Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Omran, Z. Obesity: Current Treatment and Future Horizons. Mini Rev. Med. Chem. 2017, 17, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Popkin, B.M.; Du, S.; Green, W.D.; Beck, M.A.; Algaith, T.; Herbst, C.H.; Alsukait, R.F.; Alluhidan, M.; Alazemi, N.; Shekar, M. Individuals with obesity and COVID-19: A global perspective on the epidemiology and biological relationships. Obes. Rev. 2020, 21, e13128. [Google Scholar] [CrossRef]
- Castillo, J.J.; Orlando, R.A.; Garver, W.S. Gene-nutrient interactions and susceptibility to human obesity. Genes Nutr. 2017, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Lopez, O.; Milagro, F.I.; Allayee, H.; Chmurzynska, A.; Choi, M.S.; Curi, R.; De Caterina, R.; Ferguson, L.R.; Goni, L.; Kang, J.X.; et al. Guide for Current Nutrigenetic, Nutrigenomic, and Nutriepigenetic Approaches for Precision Nutrition Involving the Prevention and Management of Chronic Diseases Associated with Obesity. J. Nutr. Nutr. 2017, 10, 43–62. [Google Scholar] [CrossRef]
- Dai, S.; Huang, B.; Zou, Y.; Liu, Y. Associations of dipping and non-dipping hypertension with cardiovascular diseases in patients with dyslipidemia. Arch. Med. Sci. 2019, 15, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef]
- Axen, K.V.; Dikeakos, A.; Sclafani, A. High dietary fat promotes syndrome X in nonobese rats. J. Nutr. 2003, 133, 2244–2249. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic acid improves high fat diet-induced hepatic steatosis and insulin resistance in mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef]
- Chen, X.; Yu, W.; Li, W.; Zhang, H.; Huang, W.; Wang, J.; Zhu, W.; Fang, Q.; Chen, C.; Li, X.; et al. An anti-inflammatory chalcone derivative prevents heart and kidney from hyperlipidemia-induced injuries by attenuating inflammation. Toxicol. Appl. Pharmacol. 2018, 338, 43–53. [Google Scholar] [CrossRef]
- Wang, Z.; Li, L.; Zhao, H.; Peng, S.; Zuo, Z. Chronic high fat diet induces cardiac hypertrophy and fibrosis in mice. Metabolism 2015, 64, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Mukhopadhyay, S. The evil axis of obesity, inflammation and type-2 diabetes. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 23–31. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, B.; Yuan, Y.; Hu, J.; Zhao, W.; Jiang, H.; Li, W.; Fan, Z.; Du, Z. Aloe Emodin Reduces Cardiac Inflammation Induced by a High-Fat Diet through the TLR4 Signaling Pathway. Mediat. Inflamm. 2020, 2020, 6318520. [Google Scholar] [CrossRef]
- Borst, S.E.; Conover, C.F. High-fat diet induces increased tissue expression of TNF-alpha. Life Sci. 2005, 77, 2156–2165. [Google Scholar] [CrossRef]
- Meldrum, D.R. Tumor necrosis factor in the heart. Am. J. Physiol. 1998, 274, R577–R595. [Google Scholar] [CrossRef] [PubMed]
- Monden, Y.; Kubota, T.; Inoue, T.; Tsutsumi, T.; Kawano, S.; Ide, T.; Tsutsui, H.; Sunagawa, K. Tumor necrosis factor-alpha is toxic via receptor 1 and protective via receptor 2 in a murine model of myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H743–H753. [Google Scholar] [CrossRef] [PubMed]
- Hamid, T.; Gu, Y.; Ortines, R.V.; Bhattacharya, C.; Wang, G.; Xuan, Y.T.; Prabhu, S.D. Divergent tumor necrosis factor receptor-related remodeling responses in heart failure: Role of nuclear factor-kappaB and inflammatory activation. Circulation 2009, 119, 1386–1397. [Google Scholar] [CrossRef]
- Murdaca, G.; Spano, F.; Contatore, M.; Guastalla, A.; Magnani, O.; Puppo, F. Pharmacogenetics of etanercept: Role of TNF-alpha gene polymorphisms in improving its efficacy. Expert Opin. Drug Metab. Toxicol. 2014, 10, 1703–1710. [Google Scholar] [CrossRef]
- Waters, J.P.; Pober, J.S.; Bradley, J.R. Tumour necrosis factor and cancer. J. Pathol. 2013, 230, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Westermann, D.; Van Linthout, S.; Dhayat, S.; Dhayat, N.; Schmidt, A.; Noutsias, M.; Song, X.Y.; Spillmann, F.; Riad, A.; Schultheiss, H.P.; et al. Tumor necrosis factor-alpha antagonism protects from myocardial inflammation and fibrosis in experimental diabetic cardiomyopathy. Basic Res. Cardiol. 2007, 102, 500–507. [Google Scholar] [CrossRef]
- Sivasubramanian, N.; Coker, M.L.; Kurrelmeyer, K.M.; MacLellan, W.R.; DeMayo, F.J.; Spinale, F.G.; Mann, D.L. Left ventricular remodeling in transgenic mice with cardiac restricted overexpression of tumor necrosis factor. Circulation 2001, 104, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.H.; Li, Y.; Cheng, K.C.; Niu, C.S.; Cheng, J.T.; Niu, H.S. Ursolic acid activates the TGR5 receptor to enhance GLP-1 secretion in type 1-like diabetic rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2017, 390, 1097–1104. [Google Scholar] [CrossRef]
- Jaikumkao, K.; Pongchaidecha, A.; Chueakula, N.; Thongnak, L.O.; Wanchai, K.; Chatsudthipong, V.; Chattipakorn, N.; Lungkaphin, A. Dapagliflozin, a sodium-glucose co-transporter-2 inhibitor, slows the progression of renal complications through the suppression of renal inflammation, endoplasmic reticulum stress and apoptosis in prediabetic rats. Diabetes Obes. Metab. 2018, 20, 2617–2626. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Hsu, C.T.; Niu, H.S.; Chang, C.H.; Cheng, J.T.; Shieh, J.M. Lung damage induced by hyperglycemia in diabetic rats: The role of signal transducer and activator of transcription 3 (STAT3). J. Diabetes Complicat. 2016, 30, 1426–1433. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Ku, P.M.; Cheng, Y.Z.; Li, Y.; Cheng, J.T.; Niu, H.S. Phosphorylation of signal transducer and activator of transcription 3 induced by hyperglycemia is different with that induced by lipopolysaccharide or erythropoietin via receptorcoupled signaling in cardiac cells. Mol. Med. Rep. 2018, 17, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Czarzasta, K.; Koperski, L.; Segiet, A.; Janiszewski, M.; Kuch, M.; Gornicka, B.; Cudnoch-Jedrzejewska, A. The role of high fat diet in the regulation of MAP kinases activity in left ventricular fibrosis. Acta Histochem. 2019, 121, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Eid, R.A.; Alkhateeb, M.A.; El-Kott, A.F.; Eleawa, S.M.; Zaki, M.S.A.; Alaboodi, S.A.; Salem Al-Shudiefat, A.A.; Aldera, H.; Alnamar, N.M.; Alassiri, M.; et al. A high-fat diet rich in corn oil induces cardiac fibrosis in rats by activating JAK2/STAT3 and subsequent activation of ANG II/TGF-1beta/Smad3 pathway: The role of ROS and IL-6 trans-signaling. J. Food Biochem. 2019, 43, e12952. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chen, J.; Zhao, J.; Meng, M. Etanercept attenuates myocardial ischemia/reperfusion injury by decreasing inflammation and oxidative stress. PLoS ONE 2014, 9, e108024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackey, D.E.; Lazaro, R.G.; Li, P.; Johnson, A.; Hernandez-Carretero, A.; Weber, N.; Vorobyova, I.; Tsukomoto, H.; Osborn, O. The role of dietary fat in obesity-induced insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E989–E997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, M.A.; Murray, A.J.; Cochlin, L.E.; Heather, L.C.; McAleese, S.; Knight, N.S.; Sutton, E.; Jamil, A.A.; Parassol, N.; Clarke, K. A high fat diet increases mitochondrial fatty acid oxidation and uncoupling to decrease efficiency in rat heart. Basic Res. Cardiol. 2011, 106, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Yida, Z.; Imam, M.U.; Ismail, M.; Ismail, N.; Ideris, A.; Abdullah, M.A. High fat diet-induced inflammation and oxidative stress are attenuated by N-acetylneuraminic acid in rats. J. Biomed. Sci. 2015, 22, 96. [Google Scholar] [CrossRef]
- Speyer, C.L.; Ward, P.A. Role of endothelial chemokines and their receptors during inflammation. J. Investig. Surg. 2011, 24, 18–27. [Google Scholar] [CrossRef]
- Khimenko, P.L.; Bagby, G.J.; Fuseler, J.; Taylor, A.E. Tumor necrosis factor-alpha in ischemia and reperfusion injury in rat lungs. J. Appl. Physiol. 1998, 85, 2005–2011. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, M.; Kennedy, R.H.; Liu, S.J. TNF-alpha-induced impairment of mitochondrial integrity and apoptosis mediated by caspase-8 in adult ventricular myocytes. Cytokine 2006, 34, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Apaijai, N.; Inthachai, T.; Lekawanvijit, S.; Chattipakorn, S.C.; Chattipakorn, N. Effects of dipeptidyl peptidase-4 inhibitor in insulin-resistant rats with myocardial infarction. J. Endocrinol. 2016, 229, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margoni, A.; Perrea, D.N.; Vlachos, I.; Prokopaki, G.; Pantopoulou, A.; Fotis, L.; Kostaki, M.; Papavassiliou, A.G. Serum leptin, adiponectin and tumor necrosis factor-alpha in hyperlipidemic rats with/without concomitant diabetes mellitus. Mol. Med. 2011, 17, 36–40. [Google Scholar] [CrossRef]
- Bayramgurler, D.; Karson, A.; Ozer, C.; Utkan, T. Effects of long-term etanercept treatment on anxiety- and depression-like neurobehaviors in rats. Physiol. Behav. 2013, 119, 145–148. [Google Scholar] [CrossRef]
- Fan, E.W.; Chen, L.J.; Cheng, J.T.; Tong, Y.C. Changes of urinary bladder contractility in high-fat diet-fed mice: The role of tumor necrosis factor-alpha. Int. J. Urol. 2014, 21, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, J.; Sauve, M.; Zhao, W.; Stacey, H.M.; Wiber, S.C.; Bolz, S.S.; Brubaker, P.L. Chronic Exposure to TNFalpha Impairs Secretion of Glucagon-Like Peptide-1. Endocrinology 2015, 156, 3950–3960. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, R.A.; Al-Mahdy, D.A.; Mouneir, S.M.; Hifnawy, M.S.; Abdel-Sattar, E.A. Anti-obesity effect of argel (Solenostemma argel) on obese rats fed a high fat diet. J. Ethnopharmacol. 2019, 238, 111893. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, Y.C.; Wu, C.C.; Huang, M.Y.; Tsai, W.C.; Hung, C.H.; Kuo, P.L. The immunomodulatory effects of TNF-alpha inhibitors on human Th17 cells via RORgammat histone acetylation. Oncotarget 2017, 8, 7559–7571. [Google Scholar] [CrossRef] [Green Version]
- Harhous, Z.; Booz, G.W.; Ovize, M.; Bidaux, G.; Kurdi, M. An Update on the Multifaceted Roles of STAT3 in the Heart. Front. Cardiovasc. Med. 2019, 6, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zgheib, C.; Zouein, F.A.; Kurdi, M.; Booz, G.W. Differential STAT3 signaling in the heart: Impact of concurrent signals and oxidative stress. Jak-Stat 2012, 1, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Fang, Q.; Zhong, P.; Chen, L.; Wang, L.; Zhang, Y.; Wang, J.; Li, X.; Wang, Y.; Wang, J.; et al. EGFR Inhibition Blocks Palmitic Acid-induced inflammation in cardiomyocytes and Prevents Hyperlipidemia-induced Cardiac Injury in Mice. Sci. Rep. 2016, 6, 24580. [Google Scholar] [CrossRef] [PubMed]
- Defer, N.; Azroyan, A.; Pecker, F.; Pavoine, C. TNFR1 and TNFR2 signaling interplay in cardiac myocytes. J. Biol. Chem. 2007, 282, 35564–35573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howerton, E.; Tarzami, S. Tumor Necrosis Factor-Alpha and Inflammation-Mediated Cardiac Injury. J. Cell Sci. Ther. 2017, 8. [Google Scholar] [CrossRef]
- Kasembeli, M.M.; Bharadwaj, U.; Robinson, P.; Tweardy, D.J. Contribution of STAT3 to Inflammatory and Fibrotic Diseases and Prospects for its Targeting for Treatment. Int. J. Mol. Sci. 2018, 19, 2299. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Sumova, B.; Mallano, T.; Chen, C.W.; Distler, A.; Bergmann, C.; Ludolph, I.; Horch, R.E.; Gelse, K.; Ramming, A.; et al. Activation of STAT3 integrates common profibrotic pathways to promote fibroblast activation and tissue fibrosis. Nat. Commun. 2017, 8, 1130. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, U.; Kasembeli, M.M.; Robinson, P.; Tweardy, D.J. Targeting Janus Kinases and Signal Transducer and Activator of Transcription 3 to Treat Inflammation, Fibrosis, and Cancer: Rationale, Progress, and Caution. Pharmacol. Rev. 2020, 72, 486–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, R.; Heusch, G. Tumor necrosis factor-alpha and its receptors 1 and 2: Yin and Yang in myocardial infarction? Circulation 2009, 119, 1355–1357. [Google Scholar] [CrossRef]
- Kleinbongard, P.; Schulz, R.; Heusch, G. TNFalpha in myocardial ischemia/reperfusion, remodeling and heart failure. Heart Fail. Rev. 2011, 16, 49–69. [Google Scholar] [CrossRef]
- Al-Lamki, R.S.; Brookes, A.P.; Wang, J.; Reid, M.J.; Parameshwar, J.; Goddard, M.J.; Tellides, G.; Wan, T.; Min, W.; Pober, J.S.; et al. TNF receptors differentially signal and are differentially expressed and regulated in the human heart. Am. J. Transplant. 2009, 9, 2679–2696. [Google Scholar] [CrossRef] [Green Version]
- De Cesaris, P.; Starace, D.; Riccioli, A.; Padula, F.; Filippini, A.; Ziparo, E. Tumor necrosis factor-alpha induces interleukin-6 production and integrin ligand expression by distinct transduction pathways. J. Biol. Chem. 1998, 273, 7566–7571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Miyamoto, T.; Yoshida, H.; Asakawa, M.; Kawasumi, M.; Kobayashi, T.; Morioka, H.; Chiba, K.; Toyama, Y.; Yoshimura, A. IL-1β and TNFα-initiated IL-6–STAT3 pathway is critical in mediating inflammatory cytokines and RANKL expression in inflammatory arthritis. Int. Immunol. 2011, 23, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Mustapha, S.; Kirshner, A.; De Moissac, D.; Kirshenbaum, L.A. A direct requirement of nuclear factor-kappa B for suppression of apoptosis in ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H939–H945. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Signaling to NF-kappaB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freund, C.; Schmidt-Ullrich, R.; Baurand, A.; Dunger, S.; Schneider, W.; Loser, P.; El-Jamali, A.; Dietz, R.; Scheidereit, C.; Bergmann, M.W. Requirement of nuclear factor-kappaB in angiotensin II- and isoproterenol-induced cardiac hypertrophy in vivo. Circulation 2005, 111, 2319–2325. [Google Scholar] [CrossRef] [Green Version]
- Kelliher, M.A.; Grimm, S.; Ishida, Y.; Kuo, F.; Stanger, B.Z.; Leder, P. The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity 1998, 8, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.W.; Shaw, J.A.; Kirshenbaum, L.A. Multiple facets of NF-kappaB in the heart: To be or not to NF-kappaB. Circ. Res. 2011, 108, 1122–1132. [Google Scholar] [CrossRef] [Green Version]
- Monaco, C.; Nanchahal, J.; Taylor, P.; Feldmann, M. Anti-TNF therapy: Past, present and future. Int. Immunol. 2015, 27, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A small-molecule inhibitor of STAT3 activation and dimerization. Chem. Biol. 2006, 13, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, K.E.; Simmons, J.G.; Ding, S.; Van Landeghem, L.; Lund, P.K. Cytokine induction of tumor necrosis factor receptor 2 is mediated by STAT3 in colon cancer cells. Mol. Cancer Res. 2011, 9, 1718–1731. [Google Scholar] [CrossRef] [Green Version]
- Orti-Casan, N.; Wu, Y.; Naude, P.J.W.; De Deyn, P.P.; Zuhorn, I.S.; Eisel, U.L.M. Targeting TNFR2 as a Novel Therapeutic Strategy for Alzheimer’s Disease. Front. Neurosci. 2019, 13, 49. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.T.; Liu, I.M.; Chi, T.C.; Tzeng, T.F.; Lu, F.H.; Chang, C.J. Plasma glucose-lowering effect of tramadol in streptozotocin-induced diabetic rats. Diabetes 2001, 50, 2815–2821. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.T.; Cheng, J.T.; Chen, Z.C. Telmisartan improves cardiac fibrosis in diabetes through peroxisome proliferator activated receptor delta (PPARdelta): From bedside to bench. Cardiovasc. Diabetol. 2016, 15, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, S.H.; Hsu, C.T.; Niu, H.S.; Niu, C.S.; Cheng, J.T.; Chen, Z.C. Ginsenoside Rh2 Improves Cardiac Fibrosis via PPARdelta-STAT3 Signaling in Type 1-Like Diabetic Rats. Int. J. Mol. Sci. 2017, 18, 1364. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.C.; Chang, W.T.; Kuo, F.Y.; Chen, Z.C.; Li, Y.; Cheng, J.T. TGR5 activation ameliorates hyperglycemia-induced cardiac hypertrophy in H9c2 cells. Sci. Rep. 2019, 9, 3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetrullo, S.; Tantini, B.; Flamigni, F.; Pazzini, C.; Facchini, A.; Stefanelli, C.; Caldarera, C.M.; Pignatti, C. Antiapoptotic and antiautophagic effects of eicosapentaenoic acid in cardiac myoblasts exposed to palmitic acid. Nutrients 2012, 4, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.C.; Yu, B.C.; Chen, L.J.; Cheng, K.C.; Lin, H.J.; Cheng, J.T. Characterization of the mechanisms of the increase in PPARdelta expression induced by digoxin in the heart using the H9c2 cell line. Br. J. Pharmacol. 2011, 163, 390–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Normal Rat | HFD-fed Rat | +Etanercept |
|---|---|---|---|
| (i) | (ii) | (iii) | |
| Body weight(g) | 447.33 ± 16.49 | 549.50 ± 19.97 * | 463.17 ± 17.09 |
| Heart weight(g) | 2.28 ± 0.08 | 2.74 ± 0.15 | 2.36 ± 0.13 |
| Blood glucose(mg/dL) | 116.50 ± 5.58 | 157.83 ± 7.05 * | 139.00 ± 4.15 # |
| Total Cholesterol(mg/dL) | 76.92 ± 4.73 | 190.14 ± 3.41 * | 169.24 ± 6.59 # |
| Triglyceride(mg/dL) | 86.86 ± 2.13 | 236.71 ± 4.53 * | 212.74 ± 4.86 # |
| HDL (mg/dL) | 27.17 ± 1.74 | 20.46 ± 0.76 * | 25.20 ± 1.07 # |
| LDL (mg/dL) | 32.38 ± 4.59 | 122.34 ± 4.51 * | 101.50 ± 5.56 # |
| AST (U/L) | 114.00 ± 6.20 | 226.50 ± 5.60 * | 168.00 ± 5.60 # |
| ALT (U/L) | 44.20 ± 3.50 | 132.70 ± 3.50 * | 109.30 ± 7.70 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-C.; Li, Y.; Hsu, C.-T.; Cheng, J.-T.; Lin, M.-H.; Cheng, K.-C.; Chen, S.-W. Etanercept Ameliorates Cardiac Fibrosis in Rats with Diet-Induced Obesity. Pharmaceuticals 2021, 14, 320. https://doi.org/10.3390/ph14040320
Hsu C-C, Li Y, Hsu C-T, Cheng J-T, Lin M-H, Cheng K-C, Chen S-W. Etanercept Ameliorates Cardiac Fibrosis in Rats with Diet-Induced Obesity. Pharmaceuticals. 2021; 14(4):320. https://doi.org/10.3390/ph14040320
Chicago/Turabian StyleHsu, Chia-Chen, Yingxiao Li, Chao-Tien Hsu, Juei-Tang Cheng, Mang-Hung Lin, Kai-Chun Cheng, and Shang-Wen Chen. 2021. "Etanercept Ameliorates Cardiac Fibrosis in Rats with Diet-Induced Obesity" Pharmaceuticals 14, no. 4: 320. https://doi.org/10.3390/ph14040320
APA StyleHsu, C. -C., Li, Y., Hsu, C. -T., Cheng, J. -T., Lin, M. -H., Cheng, K. -C., & Chen, S. -W. (2021). Etanercept Ameliorates Cardiac Fibrosis in Rats with Diet-Induced Obesity. Pharmaceuticals, 14(4), 320. https://doi.org/10.3390/ph14040320