1. Introduction
Parasitic nematodes cause diseases that produce substantial mortality and morbidity among humans across the globe. These pathogens also reduce food production in livestock and plants that, in turn, differentially restricts the nutritional resources of the global population with the lowest incomes. Anthelmintic treatments are heavily relied upon in efforts to control and treat infections caused by nematode pathogens. Impeding these efforts is the limited selection of efficacious anthelmintics coupled with emerging anthelmintic resistance among these pathogens [
1]. While the situation identifies a need to expand the number and kind of available anthelmintics, technical advances that enhance research capabilities are also needed to accelerate the accomplishment of this goal.
To advance research on anthelmintics and experimental capabilities on nematode pathogens, we established an approach with a design for pan-Nematoda applications that has a primary focus on a single-parasite tissue, the intestine. The rationale for this focus relates mainly to its apparent hypersensitivity to some anthelmintics and relative tractability to experimental approaches compared with other tissues, as described elsewhere [
2]. Capitalizing on a pan-Nematoda multiomics database for the intestine, combined with other biologic and pharmacological considerations, we experimentally derived a set of drug-like molecules referred to as nematode intestinal toxins/toxicants (NITs). Each NIT caused pathology in intestinal cells of larval stages and, in some cases, adult worms of
Ascaris suum, a model species to study the intestine of parasitic nematode species [
3]. The identification of potential inhibitors of secretory processes was the organizing principle for the
de novo identification of NITs, and multiple NITs demonstrated efficacy against phylogenetically diverse nematode species representing whipworms (Clade I), filarial worms (Clade III), and
C. elegans (Clade V) [
3].
Cellular proteins annotated as targets for each NIT collectively identify a diverse set of potential target proteins of significance in
A. suum intestinal cells [
4]. Although new approaches to kill nematode intestinal cells was a goal of this research, expression in other tissues was expected for many of the potential target proteins. In this context, an approach was developed to rapidly determine the range of cells induced to undergo cell death by experimental treatments, such as NITs, in whole
A. suum larvae in culture. Information of this kind can augment that obtained for prospective anthelmintics identified by using motility assays. The assay utilized propidium iodide (PI), a DNA-binding vital dye to detect the end point of cell death in whole nematodes in culture. Assignment of death to specific cells and organs was facilitated by anatomic details established by using a combination of the cell permeable DNA-binding dye, bisbenzimide (BB), and differential interference contrast (DIC) microscopy. The method rapidly resolved cell death in numerous cells/organs (e.g., intestine, nervous system, excretory system, seam cells, hypodermal cells, and others) of live larvae while in culture, and the results highlighted differences among cell death profiles induced by NITs. Thus, NITs represent tools to investigate cellular mechanisms leading to death in many cells of parasitic nematodes, and the PI cell death assay provides a read out for related experimental manipulations in this context. Related applications have been achieved with the non-parasitic nematode
Caenorhabditis elegans [
5,
6].
Components of early gene response patterns (GRPs) induced by NITs may reflect elements of pathways involved with initiating or mediating cell death responses. RNA-seq analysis following treatment with NITs identified early GRPs in
A. suum larvae that informed on potential target proteins and pathways (e.g.,dihydroorotate dehydrogenase (uridine synthesis) and RAB GTPase(s) (vesicle transport)) that may relate to initiator events leading to cell death induced by selected NITs [
4]. Information of this kind can aid in the design of experiments to manipulate cell death outcomes initiated by NITs and assessed by the PI cell death assay.
The overall design of our larger research effort was intended to advance knowledge in a pan-Nematoda context involving three phylogenetically diverse parasitic species, each with distinct evolutionary pathways to parasitism (
Haemonchus contortus, clade V;
A. suum, clade III; and
Trichuris suis, clade I). These three species were used for the initial computational prioritization of NITs [
3], and they collectively represent much of the phylogenetic distance in the Nematoda, which includes parasites of many species including humans, animals, and plants (where parasitism is defined as requiring a host to complete at least one stage of its life cycle, as opposed to non-parasitic “free-living” species such as
C. elegans which does not infect any hosts [
7]). Although
A. suum figured centrally in the development of this pan-Nematoda approach [
3,
4], we here sought to determine if NITs and the PI cell death assay have applications to rapidly detect death induced by NITs in cells and organs of H. contortus, leaving
T. suis the only one of the three for which applications have yet to be investigated.
H. contortus, an abomasal parasite of small ruminants, was chosen due to its veterinary importance and a number of experimental advantages it offers, including availability of large numbers of infective L3 and the ability to culture exsheathed L3 (xL3) to “parasitic” L4s.
H. contortus feeds on blood and is one of the most detrimental pathogens of small ruminants, which are important resources for nutrition and commerce in many developing regions of the world [
8,
9,
10]. The propensity of
H. contortus to rapidly acquire anthelmintic resistance makes advances to control this parasite highly important. Thus, new tools and biological knowledge related to cellular effects of experimental toxins may promote advances in anthelmintic research on
H. contortus. Although PI has been used in a general application to monitor anthelmintic effects on
H. contortus larvae [
11], resolution of individual cells and organs relative to cell death outcomes and nuances of a system to do so have not.
In research reported here, we sought to determine if NITs are toxic to H. contortus larval stages, if they cause cell death detectable by the PI assay, the identity of cells and organs that are susceptible to NIT-induced cell death, and the GRPs affected early by selected NIT treatments. The results indicated that, even with limitations stemming from interspecies biological differences, the approach has applications to H. contortus, providing new pathological and molecular tools that can be integrated into research on this model species, the progress of which will contribute to the study of nematode pathogens of humans.
3. Discussion
We explored the application of methods established with
A. suum to rapidly determine cell death in otherwise intact
H. contortus larvae in culture following treatments with seven different NITs. To the extent accomplished, the application has now been established for representatives of two phylogenetic lineages (
H. contortus, clade V;
A. suum, Clade III) of the Nematoda, while also revealing multiple differences in the performance of the assay between the two different parasites, as will be discussed. The term nematode intestinal toxins/toxicants (NITs) reflects the original focus of research to establish practical approaches that kill intestinal cells of parasitic nematodes [
3]. However, and not surprisingly, results obtained with
A. suum and now
H. contortus indicated that multiple NITs individually cause cell death in many tissues and cells beyond the intestinal tract, which demonstrates broader application of the original research focus. To investigate potential cellular target proteins and early pathways that may converge on cell death pathways, GRPs of
H. contortus larvae to three selected NITs were evaluated. Some
H. contortus gene responses resembled those found in
A. suum and suggested potential cell targets, particularly for leflunomide.
3.1. Rapid Determination of Cell Death in H. contortus
The methods used to detect cell death and assign dead cells to specific cell types or organ systems has clear application to
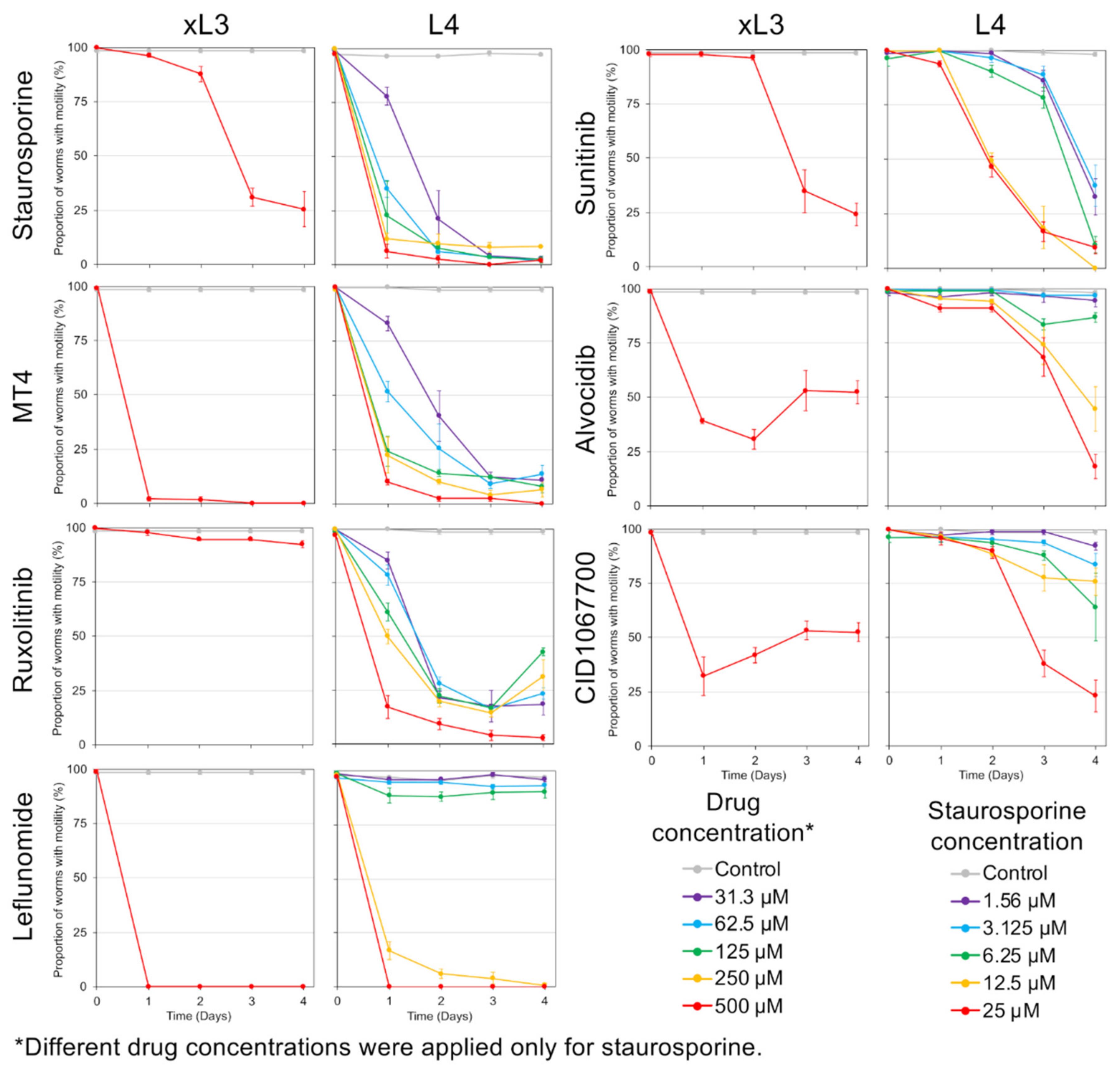
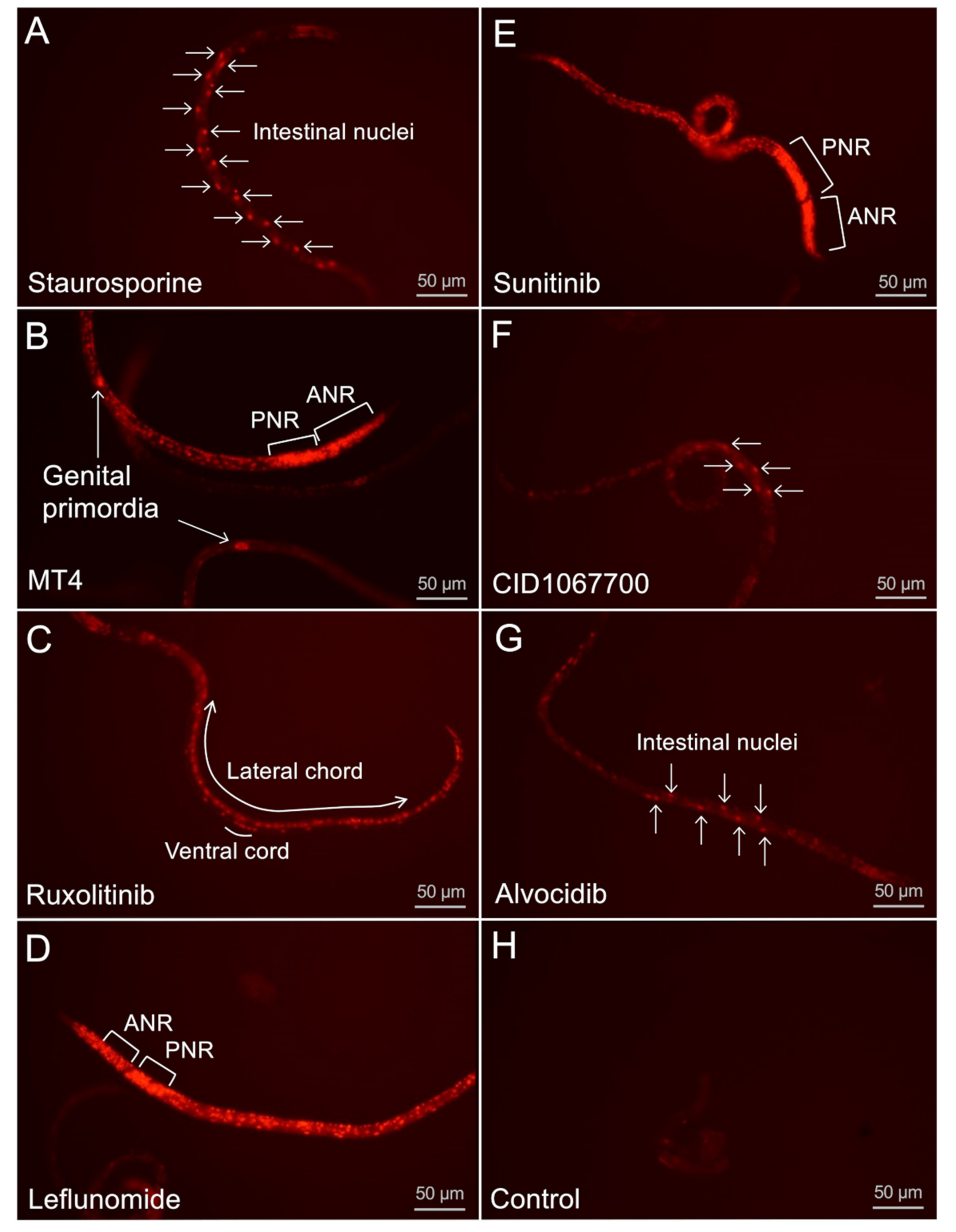
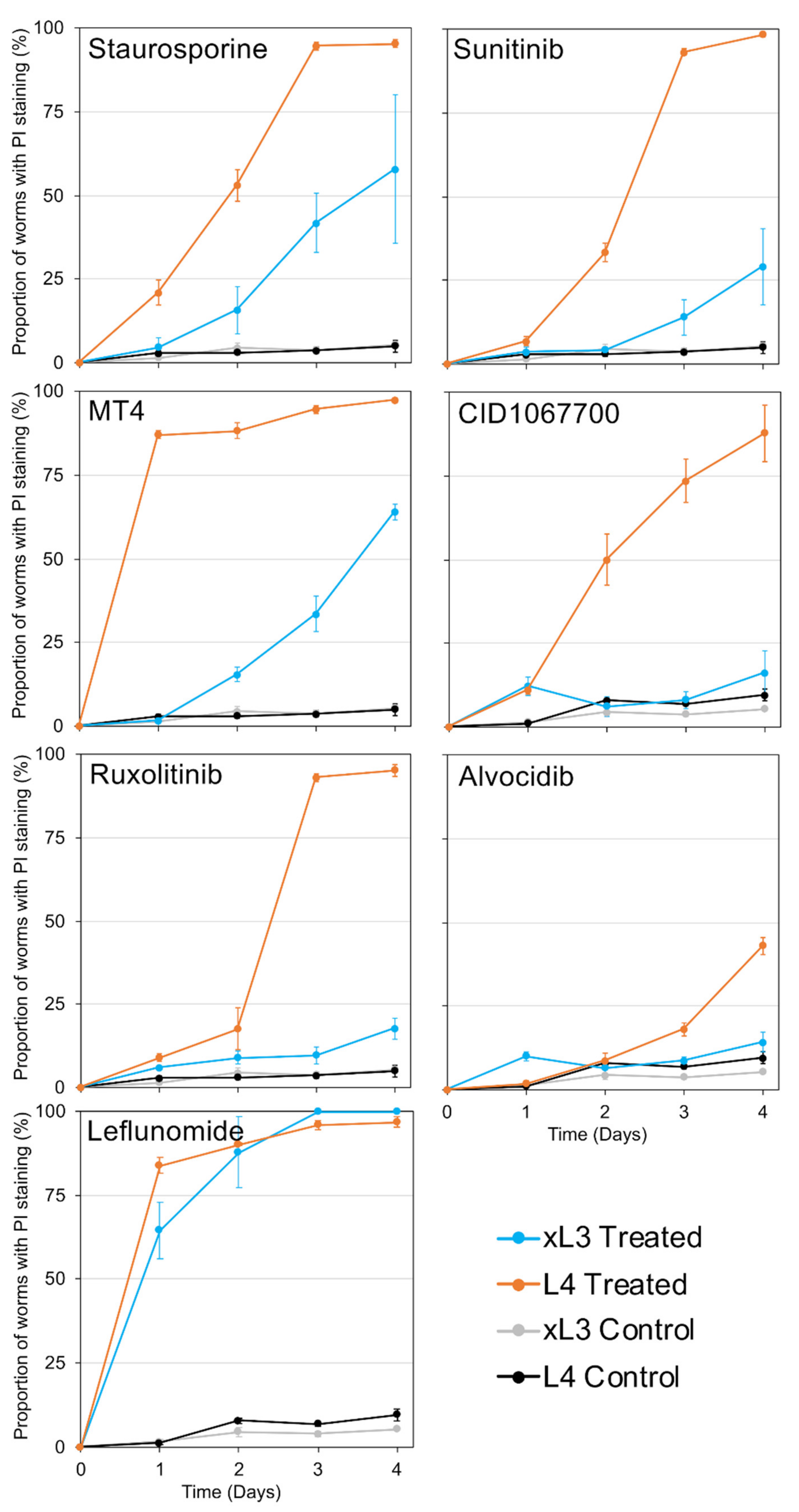
H. contortus in otherwise intact L4 and xL3 maintained in culture. The collection of cells documented as undergoing cell death varied among NITs, indicating specificity of outcome attributable to each NIT. For instance, treatments with some NITs led to PI nuclear staining primarily in intestinal cells [
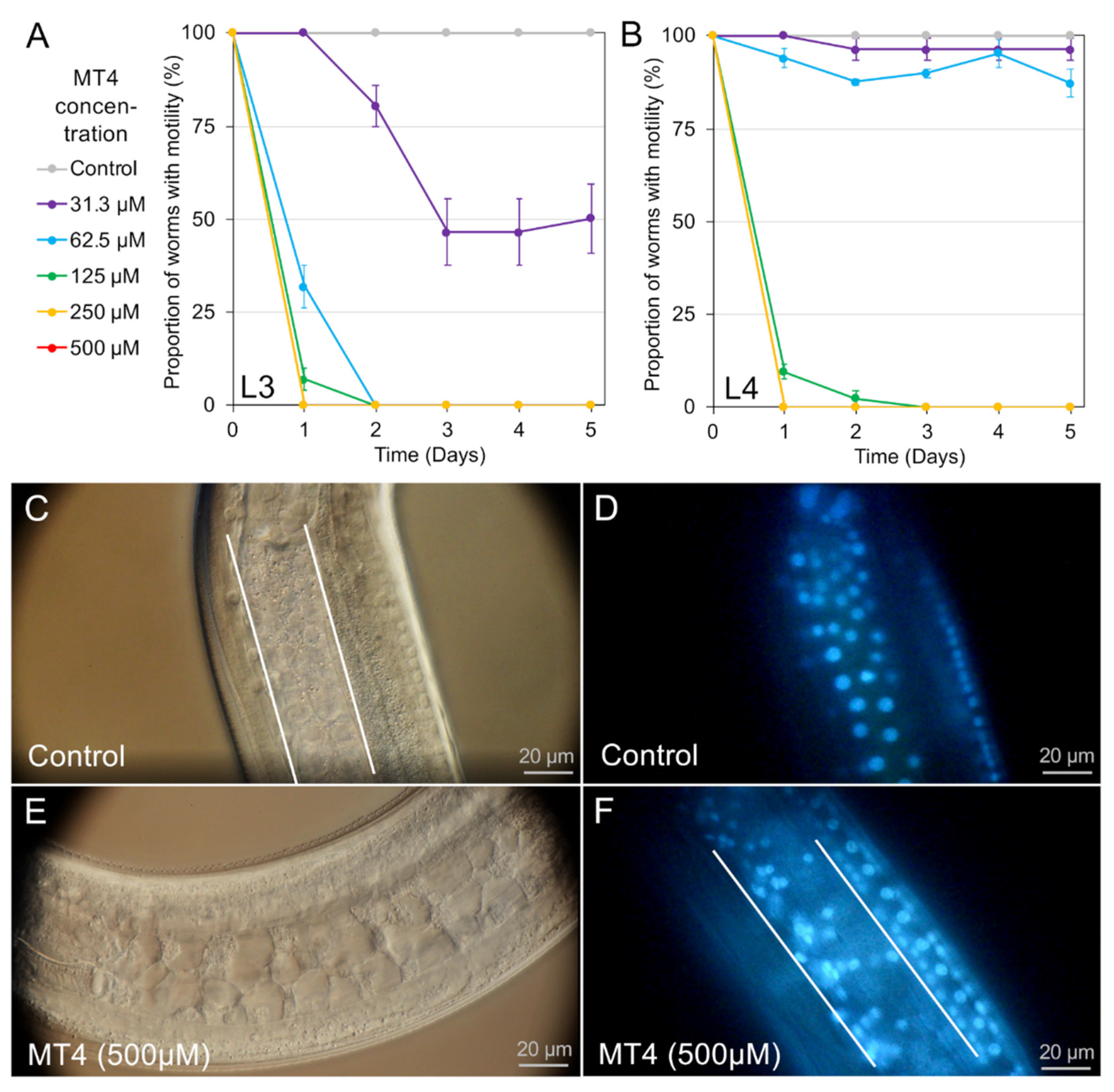
4], while for others (for example, leflunomide, sunitinib, and MT4) cell death was detected in many tissues and cells. As with
A. suum, PI nuclear staining in whole
H. contortus xL3 and L4 indicates: (1) the ability of a given treatment to induce cell death, for which rapid methods to resolve individual cells in whole worms were only recently described for parasitic nematodes [
4]; and (2) the capacity of individual cells to undergo cell death, which, once achieved, is a terminal status and one of interest for anthelmintic research. The very broad extent of PI labeling induced by treatments with leflunomide, sunitinib, and MT4 establishes that many if not all cells of
H. contortus L4, in particular, are susceptible to pharmacologic inducement of cell death pathways, making knowledge of mechanisms involved relevant to anthelmintic research. Treatment with other NITs cause more restricted PI staining, thus potentially providing experimental tools selective for more restricted cell and organ populations. The intestine, the target tissue for the initial prioritization of the NITs, appears to be more specifically targeted by the most effective NIT, staurosporine, as well as by CID1067700 and alvocidib. That individual cells or organs warrant attention for anthelmintic strategies is supported by evidence that excretory/secretory cells in microfilaria of
Brugia pahangi are an apparent individual cell target for ivermectin [
19], and intestinal cells of multiple parasitic nematodes are crucial anthelmintic targets for benzimidazole anthelmintics [
2]. Another notable finding is that some treatments caused apparent neuronal degeneration, which is a more extreme pathologic response than interference with neurotransmission, an endpoint achieved by numerous extant anthelmintics. A similar end point for neurons was noted for
A. suum [
4]. Thus, with specific neurons identified coupled with treatments that cause their degeneration and a means to detect that endpoint, valuable tools are now available to investigate mechanisms involved. Similar logic is applicable to cells identified as undergoing cell death in other organ systems.
However, an important caveat to use of PI on whole nematodes rather than cells in culture relates to potential false negative PI staining. In this case, cells may die but go undetected by PI staining due to the lack of access of PI to the damaged cell in whole worms, as discussed previously [
4]. PI must gain access across the cuticle, gut, or other access points in order to stain nuclei of dead cells within the nematode. With this caveat in mind, care is needed relative to questions investigated, experimental designs, and interpretations of results with PI staining in whole nematodes. Equally important, though, is that the caveat does not apply to positive results, which confirms access of PI to the affected cell by virtue of nuclear labeling. With an assay that detects cell death, cellular mechanisms (targets and pathways) responsible for this outcome might be investigated with other treatments (inhibitors) designed to prevent cell death based on targets annotated for NITs, e.g., cell pathways impacted by NITs (RNAseq) and mediators of cell death known in other organisms, including nematodes [
20]. A reason for interest in these mechanisms stems from the fact that cell death is pharmacologically targeted in treatments of other diseases [
21,
22] and might provide useful targets in anthelmintic strategies.
3.2. Comparisons of Microscopy and NIT Responses between H. contortus and A. suum
3.2.1. Cell and Organ Assessments
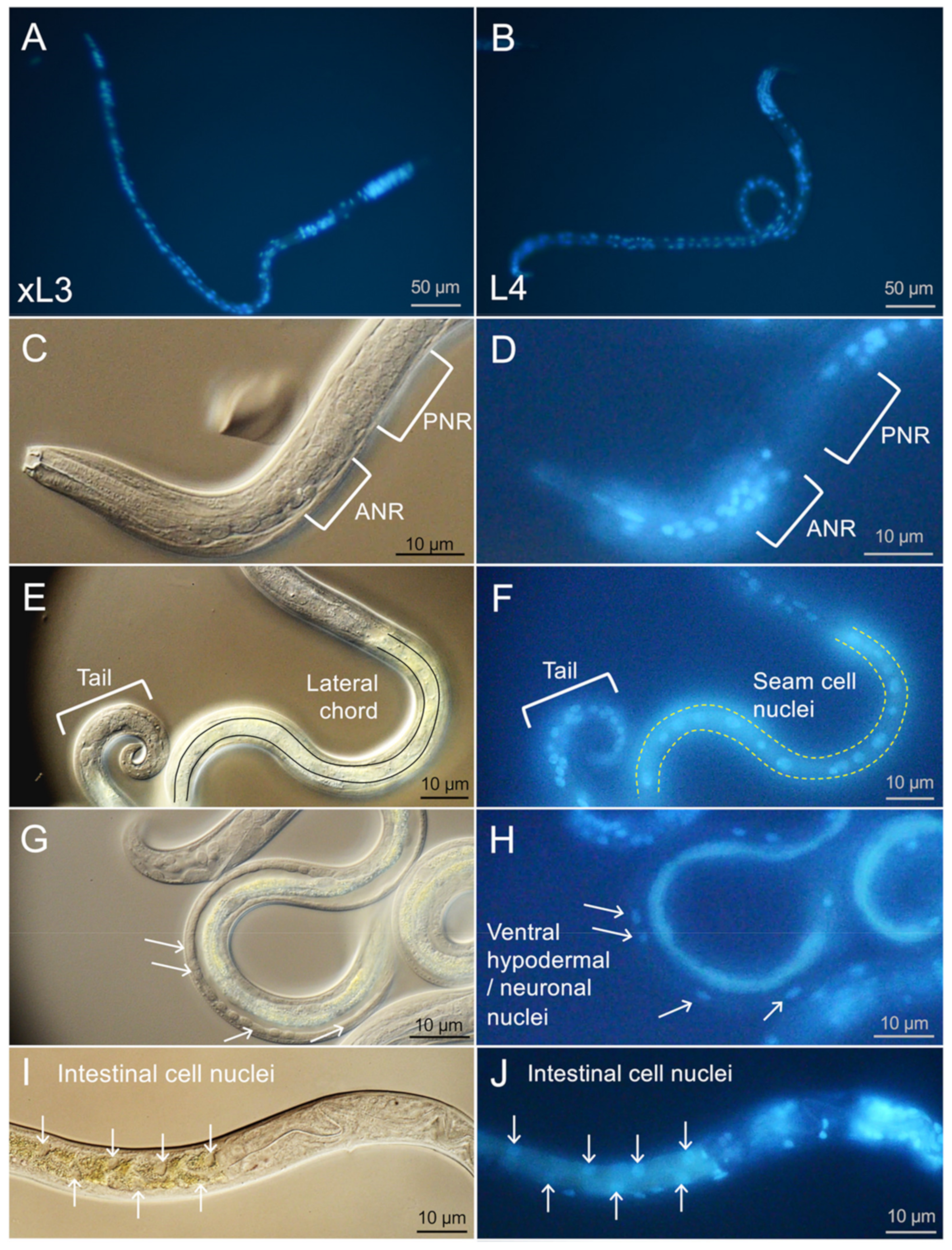
Despite this successful application of new methods to H. contortus, interspecies comparisons indicated that characteristics of A. suum larvae supported better resolution of details on pathologic effects. As one example, better performance was evident in the far more comprehensive live BB staining and DIC resolution of organs and cells in A. suum L3 and L4. The number and range of nuclei labeled by BB in live H. contortus was more restricted and similar in pattern for both xL3 and L4. Notable absences of, or restrictions on, staining in H. contortus larvae included nuclei of cells posterior to the nerve ring, intestinal cells, and many ventral chord nuclei of prospective hypodermal cells and neurons. A. suum L3 and L4 both feed, and while only H. contortus L4 feed (xL3 do not), restrictions on staining were similar for both H. contortus stages. Thus, differences in feeding competence, and internal delivery of BB by ingestion, does not alone explain the differences in BB staining between the two species. The results suggested that unknown physical differences possibly related to cuticular and/or cell membrane constituents differentially impede BB access to specific sites in H. contortus compared with A. suum larvae.
Resolution of cell and organ anatomy by DIC was also somewhat compromised in
H. contortus larvae. BB was used in conjunction with DIC to resolve anatomy with substantial clarity in
A. suum larvae. Thus, the limited scope of BB staining, light refraction by intestinal inclusions that obscured detail for DIC microscopy, and a perceived less-distinct differentiation of cells by DIC with
H. contortus larvae together translated into comparatively lower resolution of anatomical structures. Coupled with these challenges, the smaller larvae of
H. contortus also lessened confidence in assigning PI labeled nuclei to specific cells and organs in many cases (although not all, see
Figure 5) when viewed at a magnification conducive for efficient counting of tens of larvae in replicate wells (20× objective, inverted microscope). In sum, while the success in application of the methods to
H. contortus offers optimism for utility with other parasitic nematodes, the interspecies differences also indicate that specifics of the application could vary considerably among species.
3.2.2. NITs
We extended the list of NITs to include an inhibitor of p38 MAP kinases (MT4), which inhibited larval movement in
A. suum and caused demonstrable intestinal cell damage, thus meeting the definition of a NIT established in this species. MT4 also was effective against
H. contortus in motility and PI nuclear staining assays. That p38 MAP kinases are candidate targets for anthelmintics was also established in
Brugia malayi [
23], but not in relation to specific nematode cells or organs.
The target proteins ascribed for each NIT in mammalian cells are largely distinct among the NITs (summarized in [
3]). Detrimental effects resulted from treatment of
H. contortus larvae by way of immotility and cell death for each NIT, thus suggesting that these outcomes can be initiated through disruption of multiple distinct targets of NITs in nematode cells. A balancing view is that the actual targets of NITs in
A. suum and
H. contortus remain unknown and NITs have regularly been used in our experiments at relatively high concentrations to obtain positive results from which to work toward mechanisms of anthelmintic effects. Target proteins of several of the NITs include kinases and elements of signal transduction pathways, and categorically similar inhibitors have shown efficacy against
H. contortus larvae [
24], inclusive of demonstrable phenotypes beyond motility [
25]. It is also unclear if the target for an individual NIT is the same among all affected cells, such as those identified by PI staining. Still, differences in PI staining patterns observed among different NIT treatments support that perturbation of multiple different targets can lead to the outcome of cell death in both
H. contortus and
A. suum. In this regard, while NITs may or may not represent prospective anthelmintics, it seems clear that they do represent experimental tools to investigate nematode cellular targets and pathways that lead to the anthelmintic outcome of cell death in two phylogenetically distant species,
H. contortus and
A. suum.
3.2.3. Gene Response Patterns
Gene response patterns (GRPs) can provide information relevant to target proteins and pathways influenced by toxic treatments of cells [
4]. GRPs to NIT treatments also support that different NITs initiate effects via different targets and cell pathways. For instance, GRPs to selected NITs (leflunomide, staurosporine, and CID1067700) showed marked differences in
A. suum L4 [
4].
In humans, leflunomide targets only dihydroorotate dehydrogenase (DHODH) at lower doses, with some inhibition of tyrosine kinases at higher doses [
26]. Among the pathways upregulated by leflunomide in both xL3 and L4 of
H. contortus, the most significant enriched pathway was “citrate cycle (TCA cycle)”, which was also identified as the second-most enriched pathway after 2 h of leflunomide exposure in L3
A. suum [
4]. DHODH links cellular respiration (including the TCA cycle) with pyrimidine synthesis [
27], so this pathway may indicate compensation for leflunomide’s indirect downstream effects. Several pathways were enriched both among the genes most significantly upregulated and among the genes most significantly downregulated with leflunomide, including “Protein processing in endoplasmic reticulum”, a pathway previously shown to be disrupted and differentially expressed in response to leflunomide treatment in cancer cells [
28], along with the activation of “MAPK signaling”, which we identified here as being upregulated by both leflunomide and staurosporine, indicating more general NIT responsiveness. The same study [
28] also identified enrichment of the Jak-STAT signaling pathway following leflunomide treatment in cancer cells, which was the second-most upregulated pathway in this study. In addition, the pathways “thermogenesis” and “valine, leucine and isoleucine degradation” identified as being upregulated in response to leflunomide were also upregulated in our previous study of L3
A. suum [
4]. Although no direct link is obvious between these pathways and leflunomide activity, “valine, leucine and isoleucine degradation” may be linked to the described alterations in the TCA cycle, due to its production of propionyl CoA which enters the TCA cycle at the succinyl CoA step [
29]. Overall, the pathways identified here indicated that leflunomide has similar transcriptional-level effects on
H. contortus as for
A. suum, and as previously described in mammalian cell models. These results, again, are consistent with DHODH as a candidate target whose perturbation by leflunomide might initiate cellular pathways leading to cell death. The ability to induce RNAi with dsRNA treatments (reviewed in [
30]) could facilitate investigations on this hypothesis.
The most significantly enriched KEGG pathway among the genes upregulated by staurosporine treatment (
Table 3) was “signaling pathways regulating pluripotency of stem cells”, a mechanism directly supported by previous research showing that staurosporine regulates lineage choice in pluripotent cells by acting as a broad-spectrum inducer of molecular gastrulation [
31]. The other pathways only enriched among genes upregulated by staurosporine treatment include “Phosphatidylinositol signaling system” and “Inositol phosphate metabolism”, pathways which were shown to be affected by staurosporine in human platelets through the hydrolysis of phosphatidyl inositol 4,5-bisphosphate [
32], as well as “mTOR signaling pathway”, which was previously shown to be affected by staurosporine through the inhibition of phosphorylation of mTOR transcriptional regulators [
33]. Interestingly, “synaptic vesicle cycle” was upregulated by staurosporine, which contains the “exocytosis” target pathway for which staurosporine was originally prioritized as a NIT [
3]. The pathways enriched only among the genes downregulated by staurosporine (
Table 4) include “Fatty acid elongation” and “Ether lipid metabolism”, which may be related to the previously-described differential regulation of sphingolipids in response to staurosporine treatment of cancer cells [
34]. Here, we described several staurosporine-regulated pathways in
H. contortus that are consistent with results from other studies. Nevertheless, the wide range of cellular effects known to be induced by staurosporine complicates the identification of a specific molecular target.
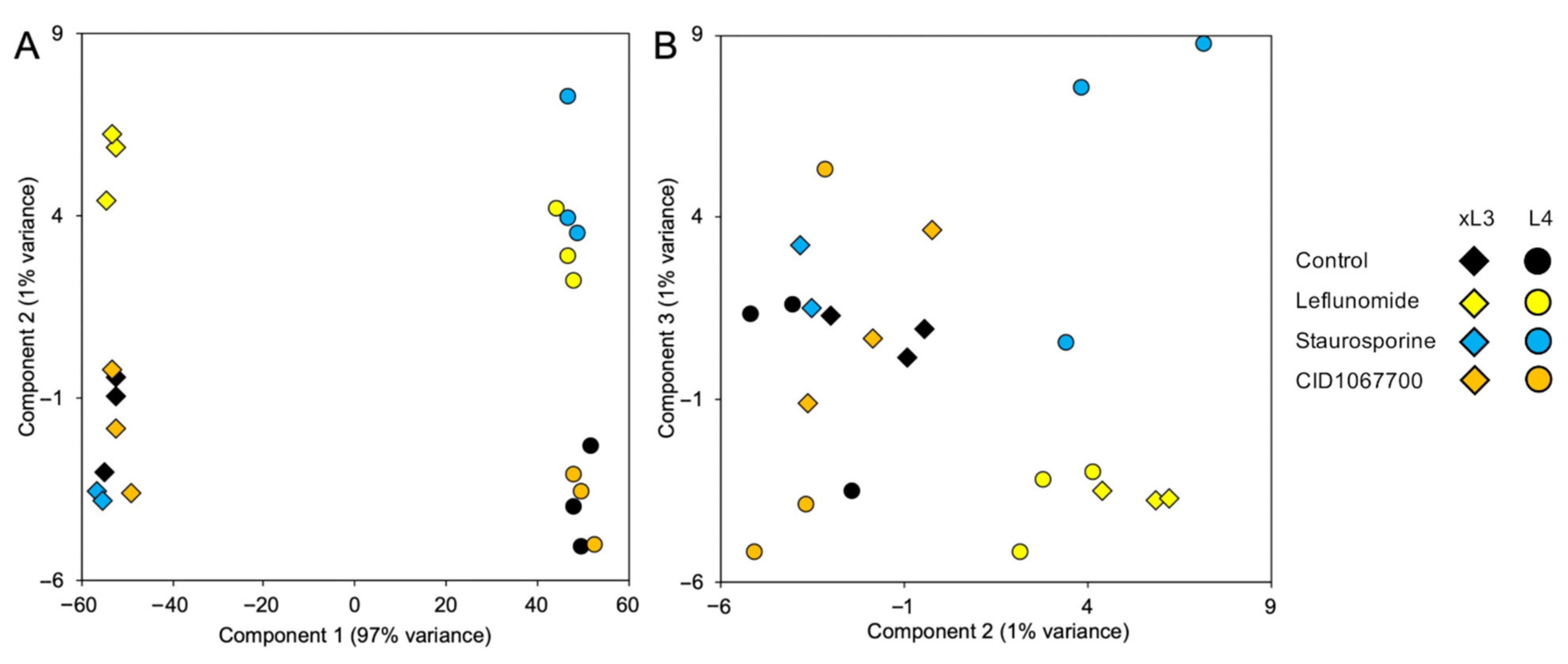
Overall, several NIT-specific and shared GRPs were identified among NITs, which were conserved across both xL3 and L4 stages. These shared pathway responses may reflect more general stress responses, even though the early treatment timepoint (2 h) aimed to minimize these general stress responses and cell death responses, in order to better elucidate the NIT-specific responses. However, since CID1067700 was relatively slow-acting in terms of motility inhibition and cell death (
Figure 3 and
Figure 5), this 2-h timepoint seemed insufficient to observe a consistent cellular GRP response. The identification of multiple unique cell targets and affected pathways reflects the differences in NIT motility responses, as well as the cell death assays showing unique NIT profiles. It should be noted that the
H. contortus xL3 larvae for the GRP analysis were obtained from a different laboratory-passaged strain than the rest of the experimentation, but the results obtained here were consistent with our previous
A. suum study with a similar design, as well as with existing literature describing cellular responses to these NITs, indicating broad phylogenetically consistency in application of methods and responses to NITs. However, the phylogenetic levels at which differences emerge is not known and could include interstrain variability. Overall, our results highlighted that the NITs have unique and complex mechanisms of action that do not simply immediately result in cellular death.
4. Materials and Methods
4.1. Ethics Statement
All animal experiments were carried out under protocols approved by Washington State University Institutional Animal Care and Use Committee, protocol 4097. The protocols meet requirements of AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; Guide for the Care and Use of Laboratory Animals: 2011 Edition, National Research Council, and USA Animal Welfare Act and Animal Welfare Regulations: 2017 Edition (AWA), US Department of Agriculture.
4.2. Haemonchus contortus L3, L4 and Adults
Haemonchus contortus L3 larvae were obtained from two sources; small batches of laboratory-passaged L3 were purchased from Dr. Ray Kaplan (University of Georgia, Department of infectious diseases, Athens, GA, USA) and used in most experiments presented here, and large batches (1 to 2 million) of laboratory-passaged L3 provided by Zoetis (Dr. Debra Woods, Global Parasitology Research at Zoetis, Kalamazoo, MI, USA) were used to determine gene transcript responses to experimental treatments. Isolates from both sources were designated as susceptible to contemporary anthelmintics. Exsheathed L3 (xL3) were produced by incubation of L3 in 0.125% sodium hypochlorite at 37 °C for twenty minutes. xL3 were washed 5 times in sterile phosphate buffered saline (pH 7.4) by pelleting larvae at 600× g for five minutes, discarding the supernatant, and then adding PBS for the next rinse. After the final rinse, pelleted xL3 were resuspended in RPMI medium (R8758, Sigma-Aldrich, St. Louis MO) containing 20% fetal bovine serum, 100 units penicillin, and 100 μg Streptomycin per mL (P0781, Sigma-Aldrich, St. Louis, MO, USA) and incubated at 38 °C with 20% CO2. L4 were generated by incubation of xL3 in this conditioned media for 7 days. Acquisition of L4 characteristics (well-defined mouth) was observed by this time in a mean 95.3% (±1.3%) of larvae. Culture volumes were 100 μL in 96 well plates (3595, Costar, Corning Inc., Corning, NY, USA) and 1 mL in 24 well plates for applications in this research.
4.3. Ascaris suum L3, L4
A. suum lung-stage L3 were obtained as described before [
3]. Briefly, infective larvated eggs were used from batches generated from previous studies [
3,
4]. Then, 4000 eggs were used to orally infect New Zealand white rabbits (5.5 to 6.5 weeks old, Western Oregon Rabbit Company, Philomath, OR, USA) and L3 were collected from the lungs by lavage. Larvae were settled by gravity and then washed in 3 sequential 50 mL volumes of warm PBS followed by 3 sequential 15 mL volumes, with intervening gravity sedimentation and discard of supernatant PBS. Extracted and cleaned larvae were then suspended in RPMI medium (R8758, Sigma-Aldrich, St. Louis, MO, USA, containing 10% swine serum, 100 units penicillin and 100 μg Streptomycin/mL; P0781, Sigma-Aldrich, St. Louis, MO, USA) and then, if used for testing as L3, dispensed into wells of 96-well plates (3595, Costar, Corning Inc., Corning, NY, USA, triplicate wells for each treatment), with a total volume of 100 μL culture medium containing treatments with diluent (DMSO, 922401 JT Baker, Center Valley, PA, USA) at 1%. L3 were then incubated at 37 °C in 5% CO
2 for times prescribed for each experimental design. Alternatively, to obtain L4, L3 were incubated in 1 mL culture medium contained in a 15 mL polypropylene tube (62.554.100, Sarstedt, Newton, NC, USA) with a loosened screw cap for 3 days, and medium was replaced daily with fresh medium. L3 molted to L4 by day 3. L4 generated in this way were dispensed into wells of 96 well plates used for experiments described here and incubated as for L3.
4.4. Fluorescent Marker Labeling
To determine the ability of xL3 and L4
H. contortus to ingest fluorescent probes, two function-based fluorescent probes were used in feeding assays: (1) beta-ala,lys-AMCA (BAL-AMCA), 200 μM (BP0352, BioTrend, Zurich, Switzerland) [
12]; and (2) DQ Green-BSA (D-12050, Molecular Probes, Eugene, OR, USA), 100 μg/mL [
13]. L3 and L4 were cultured in the presence of these probes for 4 h prior to assessment of ingestion. To assess staining of nuclei by bisbenzimide (BB, 10 µg/mL, H 33258, Sigma-Aldrich, St. Louis, MO, USA) live xL3 and L4 were incubated with BB in standard culture conditions for 16 h prior to assessment by fluorescent microscopy. To assess cell death in whole xL3 and L4, propidium iodide (PI, P4170, Sigma-Aldrich, St. Louis, MO, USA) was used at 100 μM, and added to larval cultures 4 h prior to the addition of experimental treatments. Preincubation leading to ingestion of PI was intended to maximize access of tissues to the fluorescent probes in presence of various treatments.
4.5. Microscopy and Imaging
Fluorescent labeling of larvae in wells of 96 well plates was monitored using a Nikon Diaphot 300 inverted microscope equipped with epifluorescence capabilities: UV-2A filter (blue), BB, BAL-AMCA; G1A (red filter), PI; B2A (green filter), DQ-BSA) and recorded with a Nikon D5100 digital camera. Most observations were made using the 20× microscope objective (200× magnification). To better resolve cells and organs in which BB-stained nuclei reside, larvae were transferred from wells and placed onto agar pads on glass microscope slides and viewed using the 60× microscope objective (600× magnification) on a Nikon Optiphot compound microscope equipped with differential interference contrast (DIC) filters, epifluorescence capabilities, and a Nikon D5100 digital camera. To optimize resolution of DIC/fluorescence details, images were captured in movie mode, and then selected screen shots were copied and used to produce final digital images.
4.6. NIT Treatments
A set of chemicals identified as nematode intestinal toxins/toxicants (NITs) [
3] were used in experiments here and included: alvocidib (S1230), sunitinib (S7781), Selleckchem Houston, TX, USA; CID1067700 (SML054), leflunomide (L5025), p38 MAP kinase inhibitor IV (a.k.a. MT4, which was used in this paper, SML0543) Sigma-Aldrich, St. Louis, MO, USA; ruxolitinib (tlrl-rux), InvivoGen, San Diego, CA, USA; staurosporine (S-9300), LC Laboratories, Woburn, MA, USA. DMSO was used as diluent for chemical treatments, except propidium iodide was solubilized in PBS, and concentrations were adjusted to achieve a maximum 1% DMSO after all components were included for each experiment. Control wells were adjusted according to diluent used for treatment wells. With the exception of staurosporine, treatments were initiated at 500 μM (staurosporine, 25 μM) to better ensure the possibility of achieving a positive result with PI labeling based on previous experience [
3]. Dose response experiments to determine effective dose 50 (IC
50) were conducted using two-fold serial dilutions over a range of 5 concentrations. IC
50 values were calculated using AAT Bioquest (Sunnyvale, CA, USA) IC
50 calculator (
https://www.aatbio.com/tools/IC50-calculator/ (accessed on June 17 2021)).
4.7. Motility PI labeling Assays
xL3 and L4 were dispensed into triplicate wells of 96 well plates containing more than twenty larvae each, in 100 μL of medium containing treatments. Movement was scored using the inverted microscope, and 200× magnification. Plates were agitated manually to encourage movement of larvae and larval movement was scored if observed during an approximate 45 sec observation period. Mean motility for each treatment was determined for each day over the course of 4 days. The setup for PI labeling assays was identical, except that wells also contained PI and BB, and PI labeling was scored with the inverted fluorescent microscope using 200× magnification. A conservative PI scoring system was used in that more than 5 nuclei labeled with PI in a given larva were required to be scored positive. The mean number of PI positive larvae was determined for each treatment, each day, over a course of days.
4.8. RNA Extraction, RNA-seq and Differential Expression Analysis
For RNAseq analysis (see below), approximately 80,000
H. contortus xL3 or L4 were aliquoted into each of triplicate wells in a 48 well plate and 1 mL of media for each treatment. The treatments included leflunomide (500 μM), staurosporine (25 μM), CID1067700 (500 μM), or diluent (1% DMSO) followed by incubation for 2 h, then rinsing with three 1 mL volumes of PBS (7.4), withdrawing excess PBS and mixing larval pellets with 50 μL TRIzol (Invitrogen/Life Technologies, Carlsbad, CA, USA), and then freezing samples at −80 °C until processed for RNA. RNA was extracted from pellets of untreated and treated larval
H. contortus frozen in Trizol (described under
Section 2.5) for RNAseq analysis by homogenizing larval pellets as they thawed using a microfuge pestle, and then processing TRIzol extracts according to the manufacturer’s instructions. Ethanol pellets of isolated RNA were shipped to Washington University for further processing and sequencing. RNA-seq analysis was performed as previously described for
Ascaris suum [
4]. Briefly, cDNA libraries were prepared from RNA samples using the Clontech SMARTer universal low-input RNA kit to maximize yield and processed cDNA was sequenced on the Illumina NovaSeq S4 platform (paired-end 150bp reads). Trimmomatic v0.36 [
35] was used to trim adapters, and the STAR aligner [
36] (v2.7.3a; 2-pass mode, basic) was used to map RNA-seq reads to the
H. contortus genome assembly (PRJEB506.WBPS14 [
37]), which were then counted for every annotated protein-coding gene in the assembly using featureCounts [
38]. DESeq2 [
39] was used to perform differential expression comparisons, comparing each NIT the control samples collected for both xL3 and L4-stage larvae. Principal components analysis (PCA) was carried out using DESeq2 output (default settings). The raw RNA-seq read files (fastq) are accessible on the NCBI Sequence Read Archive (SRA [
40], BioProject PRJNA264197), and complete sample metadata, read counts, normalized expression values, and accession information are provided in
Supplementary Table S1.
H. contortus genes based on the PRJEB506.WBPS14 annotation and assembly [
37] were assigned functional annotations using both InterProScan v5.42 [
41] to identify gene ontology assignments as well as InterPro domains, and GhostKOALA v2.2 [
42] to assign KEGG annotations. Potentially secreted proteins were identified using both SignalP v5.0 [
43] for signal peptides and transmembrane domains, and SecretomeP v2.0 [
44] to identify proteins with non-classical secretion sequences. Gene set enrichment analysis (GSEA [
18]) was performed, based on DESeq2 differential expression results, to test for enrichment among non-human KEGG pathway annotations using WebGestalt v2019 [
45] (FDR-adjusted
p ≤ 0.05, minimum 5 genes differentially expressed). The input values for GSEA analysis were the ranks (largest value to smallest) of the genes when ranked based on (a) differential expression
p value from DESeq2 analysis, following log transformation, where upregulated genes were calculated using negative log, and downregulated genes were calculated using positive log, and (b) Log
2 fold change value when
p values were tied. The top 15 most significantly enriched KEGG pathways shared between both xL3 and L4 for each NIT were identified (according to FDR-adjusted
p values). “Normalized enrichment scores” shown in
Table 1,
Table 2,
Table 3 and
Table 4 were the primary statistic calculated by Gene Set Enrichment Analysis to evaluate results, and represent the degree to which a gene set is overrepresented at the top or bottom of a ranked list of genes, normalized for differences in gene set sizes (since smaller gene sets are more likely to all be close to the top of a list by random chance).
5. Conclusions
This study presented an interdisciplinary approach designed to intersect (i) methodology that comprehensively resolved, in whole unfixed parasites, NIT-induced pathology among most cells and organs in
H. contortus, and (ii) interrogation of transcriptional responses to NIT treatments, resulting in the identification of their putative molecular targets and pathways. The assays directly identified the constellation of cells and organ systems that undergo cell death with respect to each of multiple chemical toxins/toxicants. Susceptibility to inducible cell death adds rationale to investigate the cells and organs identified in anthelmintic research, and advances established here provided new experimental methods to do so. We also identified, for some NITs, cellular pathways and molecular targets that are potential antecedents to irreparable pathologic outcomes. Our original derivation of NITs involved a
de novo approach [
3] which, when building on methodology we have developed [
4] and integrated with results described here, establish a unique and useful experimental system for the purposes of discovery and detailed analysis of anthelmintic compounds and investigation of mechanisms of their toxicity. Ultimately, knowledge of mechanisms that actually mediate cell death coupled with pathways whose disruption initiate death processes hold much potential for application in anthelmintic research. The results shown here confirmed cross-species efficacy of the NITs, which were computationally predicted to have broad efficacy across nematodes. In addition to their previously demonstrated efficacy against
A. suum, an important parasite of humans, our results demonstrated efficacy against species of a great veterinary importance, the blood feeding parasitic nematode of small ruminants
H. contortus. In the future, the efficacy of these NITs may also be tested to other parasitic nematodes of economic and clinical importance, including as plant or aquatic parasitic nematodes, given that here, we demonstrated consistent efficacy spanning multiple nematode clades.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}