
Red Algal Sulfated Galactan Binds and Protects Neural Cells from HIV-1 gp120 and Tat

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
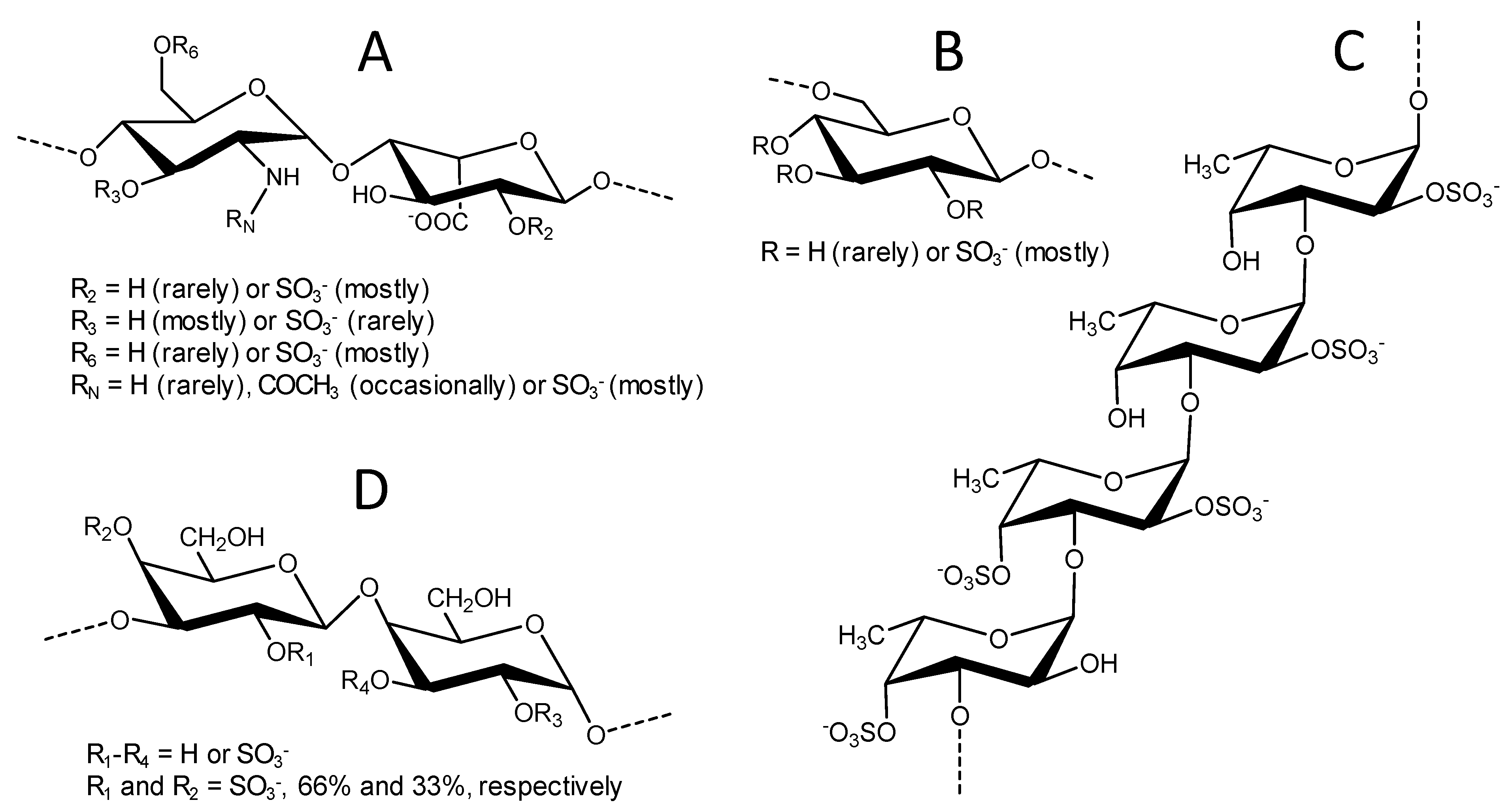
2.1. Structure and Selection of Sulfated Glycans
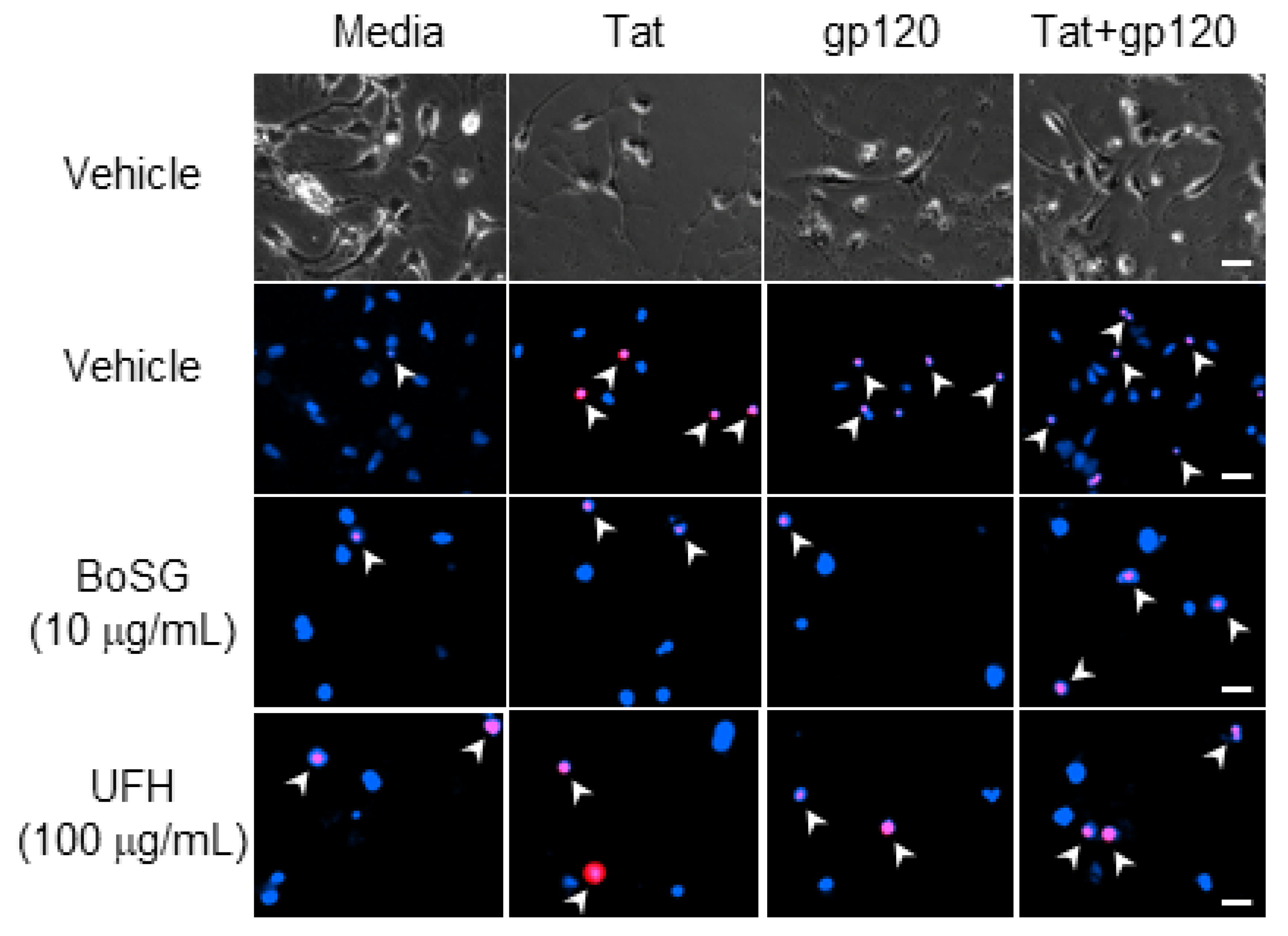
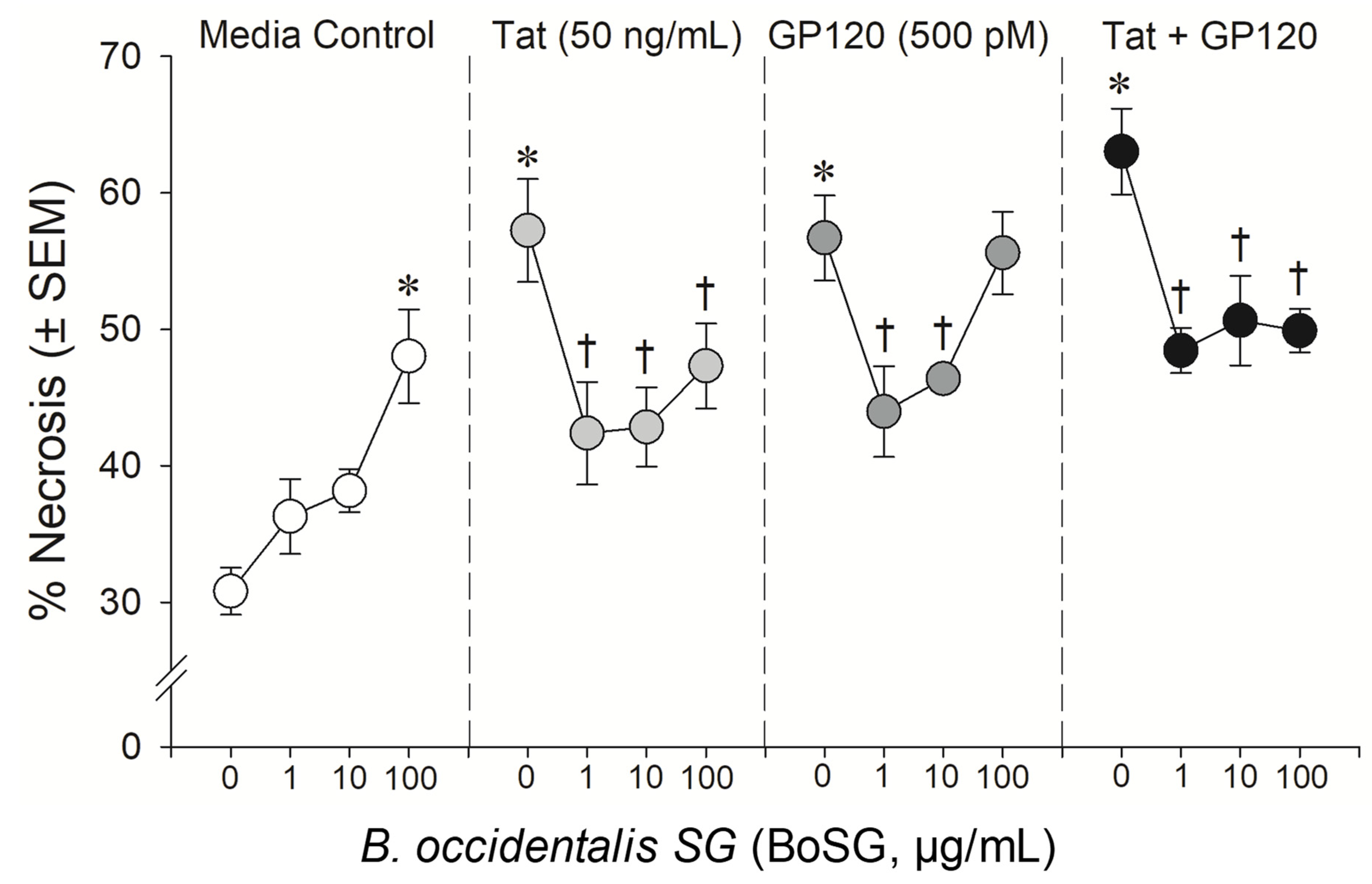
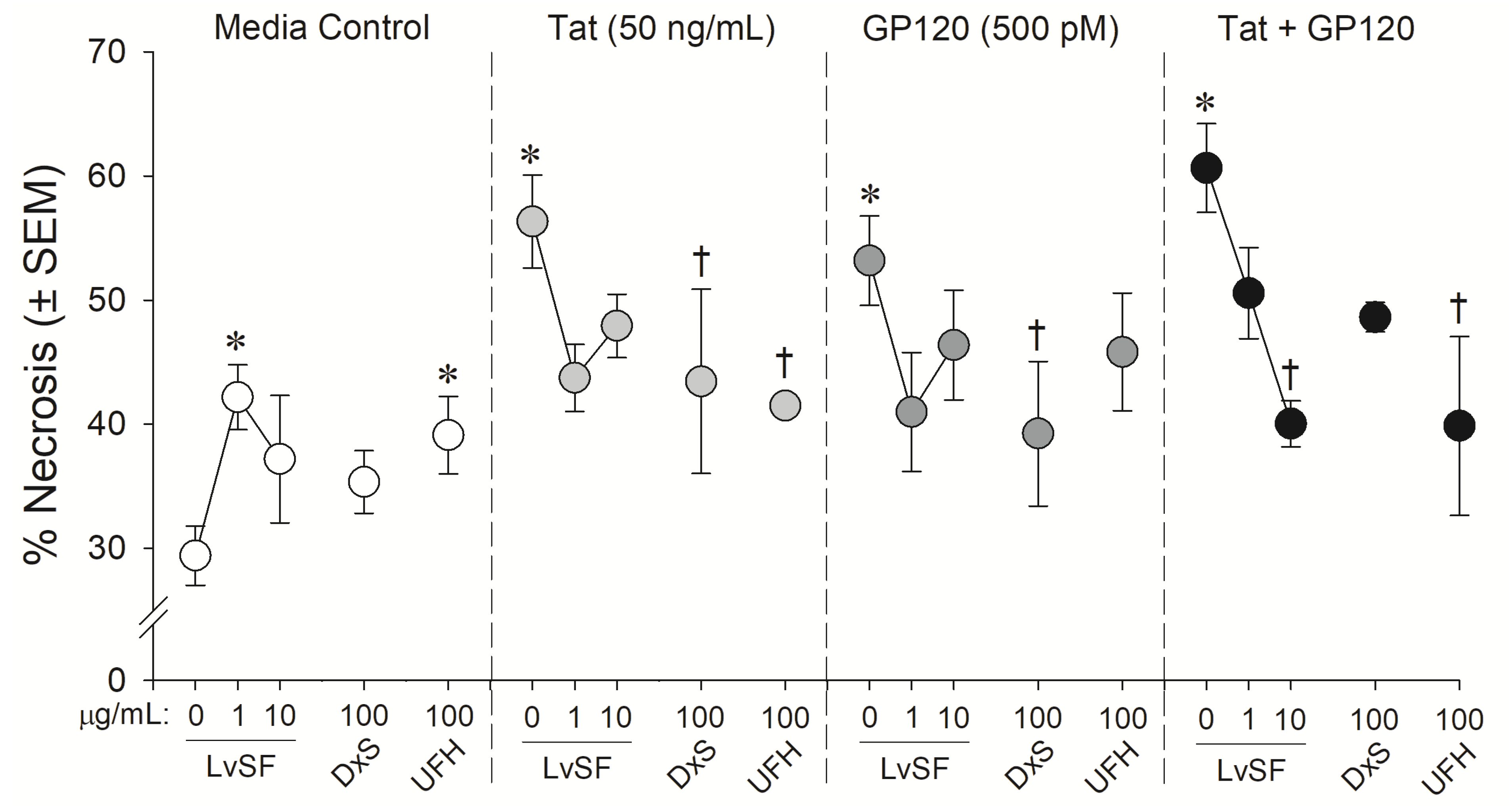
2.2. Neuroprotective Capacity of Marine and Control Sugars against HIV-1 Proteins
2.3. Binding Capacity of BoSG and UFH of HIV-1 Proteins
3. Discussion
4. Materials and Methods
4.1. Chemical, Reagents and Instruments
4.2. Sulfated Glycans
4.3. Live/Dead Neural Cell-Based Assay
4.3.1. Neural Cell Co-Culture
4.3.2. Live/Dead Assay
4.3.3. Statistical Analyses
4.4. SPR
4.4.1. Preparation of BoSG and UFH Biochip
4.4.2. Measurement of Interaction between Sulfated Glycans and Proteins Using BIAcore
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nickoloff-Bybel, E.A.; Calderon, T.M.; Gaskill, P.J.; Berman, J.W. HIV Neuropathogenesis in the Presence of a Disrupted Dopamine System. J. Neuroimmune Pharmacol. 2020, 15, 729–742. [Google Scholar] [CrossRef]
- Ajasin, D.; Eugenin, E.A. HIV-1 Tat: Role in Bystander Toxicity. Front. Cell Infect. Microbiol. 2020, 10, 61. [Google Scholar] [CrossRef]
- Thaney, V.E.; Kaul, M. Type I Interferons in NeuroHIV. Viral Immunol. 2019, 32, 7–14. [Google Scholar] [CrossRef]
- Raybuck, J.D.; Hargus, N.J.; Thayer, S.A. A GluN2B-Selective NMDAR Antagonist Reverses Synapse Loss and Cognitive Impairment Produced by the HIV-1 Protein Tat. J. Neurosci. 2017, 37, 7837–7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schier, C.J.; Marks, W.D.; Paris, J.J.; Barbour, A.J.; McLane, V.D.; Maragos, W.F.; McQuiston, A.R.; Knapp, P.E.; Hauser, K.F. Selective Vulnerability of Striatal D2 versus D1 Dopamine Receptor-Expressing Medium Spiny Neurons in HIV-1 Tat Transgenic Male Mice. J. Neurosci. 2017, 37, 5758–5769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speidell, A.; Asuni, G.P.; Wakulski, R.; Mocchetti, I. Up-regulation of the p75 neurotrophin receptor is an essential mechanism for HIV-gp120 mediated synaptic loss in the striatum. Brain Behav. Immun. 2020, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Langford, D.; Oh Kim, B.; Zou, W.; Fan, Y.; Rahimain, P.; Liu, Y.; He, J.J. Doxycycline-inducible and astrocyte-specific HIV-1 Tat transgenic mice (iTat) as an HIV/neuroAIDS model. J. Neurovirol. 2018, 24, 168–179. [Google Scholar] [CrossRef]
- Thaney, V.E.; Sanchez, A.B.; Fields, J.A.; Minassian, A.; Young, J.W.; Maung, R.; Kaul, M. Transgenic mice expressing HIV-1 envelope protein gp120 in the brain as an animal model in neuroAIDS research. J. Neurovirol. 2018, 24, 156–167. [Google Scholar] [CrossRef]
- Vivès, R.R.; Imberty, A.; Sattentau, Q.J.; Lortat-Jacob, H. Heparan Sulfate Targets the HIV-1 Envelope Glycoprotein gp120 Coreceptor Binding Site. J. Biol. Chem. 2005, 280, 21353–21357. [Google Scholar] [CrossRef] [Green Version]
- Connell, B.; Lortat-Jacob, H. Human Immunodeficiency Virus and Heparan Sulfate: From Attachment to Entry Inhibition. Front. Immunol. 2013, 4, 385. [Google Scholar] [CrossRef] [Green Version]
- Crublet, E.; Andrieu, J.-P.; Vivès, R.R.; Lortat-Jacob, H. The HIV-1 Envelope Glycoprotein gp120 Features Four Heparan Sulfate Binding Domains, Including the Co-receptor Binding Site. J. Biol. Chem. 2008, 283, 15193–15200. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H.; Bezerra, F.F.; Soares, P.A.G. Sulfated Glycans in HIV Infection and Therapy. Curr. Pharm. Des. 2017, 23, 3405–3414. [Google Scholar] [CrossRef] [PubMed]
- Argyris, E.G.; Kulkosky, J.; Meyer, M.E.; Xu, Y.; Mukhtar, M.; Pomerantz, R.J.; Williams, K.J. The perlecan heparan sulfate proteoglycan mediates cellular uptake of HIV-1 Tat through a pathway responsible for biological activity. Virology 2004, 330, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.C.; Samaniego, F.; Nair, B.C.; Buonaguro, L.; Ensoli, B. HIV-1 Tat protein exits from cells via a leaderless secretory pathway and binds to extracellular matrix-associated heparan sulfate proteoglycans through its basic region. AIDS 1997, 11, 1421–1431. [Google Scholar] [CrossRef]
- Richard, J.P.; Melikov, K.; Brooks, H.; Prevot, P.; Lebleu, B.; Chernomordik, L.V. Cellular Uptake of Unconjugated TAT Peptide Involves Clathrin-dependent Endocytosis and Heparan Sulfate Receptors. J. Biol. Chem. 2005, 280, 15300–15306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, M.; Rusnati, M.; Presta, M.; Giacca, M. Internalization of HIV-1 Tat Requires Cell Surface Heparan Sulfate Proteoglycans. J. Biol. Chem. 2001, 276, 3254–3261. [Google Scholar] [CrossRef] [Green Version]
- Urbinati, C.; Nicoli, S.; Giacca, M.; David, G.; Fiorentini, S.; Caruso, A.; Alfano, M.; Cassetta, L.; Presta, M.; Rusnati, M. HIV-1 Tat and heparan sulfate proteoglycan interaction: A novel mechanism of lymphocyte adhesion and migration across the endothelium. Blood 2009, 114, 3335–3342. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.; Seelig, J. Interaction of the protein transduction domain of HIV-1 TAT with heparan sulfate: Binding mechanism and thermodynamic parameters. Biophys. J. 2004, 86, 254–263. [Google Scholar] [CrossRef] [Green Version]
- De Francesco, M.A.; Baronio, M.; Poiesi, C. HIV-1 p17 matrix protein interacts with heparan sulfate side chain of CD44v3, syndecan-2, and syndecan-4 proteoglycans expressed on human activated CD4+ T cells affecting tumor necrosis factor alpha and interleukin 2 production. J. Biol. Chem. 2011, 286, 19541–19548. [Google Scholar] [CrossRef] [Green Version]
- Poiesi, C.; De Francesco, M.A.; Baronio, M.; Manca, N. HIV-1 p17 binds heparan sulfate proteoglycans to activated CD4+ T cells. Virus Res. 2008, 132, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Francesca, C.; Stefania, M.; Simona, F.; Arnaldo, C.; Cinzia, G. HIV-1 Matrix Protein p17 and its Receptors. Curr. Drug Targets 2016, 17, 23–32. [Google Scholar]
- Cladera, J.; Martin, I.; O’Shea, P. The fusion domain of HIV gp41 interacts specifically with heparan sulfate on the T-lymphocyte cell surface. EMBO J. 2001, 20, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Amornrut, C.; Toida, T.; Imanari, T.; Woo, E.-R.; Park, H.; Linhardt, R.; Wu, S.J.; Kim, Y.S. A new sulfated β-galactan from clams with anti-HIV activity. Carbohydr. Res. 1999, 321, 121–127. [Google Scholar] [CrossRef]
- Huang, N.; Wu, M.-Y.; Zheng, C.-B.; Zhu, L.; Zhao, J.-H.; Zheng, Y.-T. The depolymerized fucosylated chondroitin sulfate from sea cucumber potently inhibits HIV replication via interfering with virus entry. Carbohydr. Res. 2013, 380, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Mulloy, B. Current structural biology of the heparin interactome. Curr. Opin. Struct. Biol. 2015, 34, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Tazi, L.M.; Jayawickreme, S. Determination of residual dextran sulfate in protein products by SEC–HPLC. J. Chromatogr. B 2016, 1011, 89–93. [Google Scholar] [CrossRef]
- Mulloy, B.; Ribeiro, A.C.; Alves, A.P.; Vieira, R.P.; Mourão, P.A. Sulfated fucans from echinoderms have a regular tetrasaccharide repeating unit defined by specific patterns of sulfation at the 0–2 and 0-4 positions. J. Biol. Chem. 1994, 269, 22113–22123. [Google Scholar] [CrossRef]
- Farias, W.R.L.; Valente, A.-P.; Pereira, M.S.; Mourão, P.A.S. Structure and Anticoagulant Activity of Sulfated Galactans: Isolation of a unique sulfated galactan from the red algaebotryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H. Fucanomics and galactanomics: Current status in drug discovery, mechanisms of action and role of the well-defined structures. Biochim. Biophys. Acta 2012, 1820, 1971–1979. [Google Scholar] [CrossRef]
- Quinderé, A.-L.G.; Santos, G.R.C.; Oliveira, S.-N.; Glauser, B.F.; Fontes, B.P.; Queiroz, I.N.L.; Benevides, N.M.B.; Pomin, V.H.; Mourão, P.A.S. Is the antithrombotic effect of sulfated galactans independent of serpin? J. Thromb. Haemost. 2014, 12, 43–53. [Google Scholar] [CrossRef]
- Baroletti, S.A.; Goldhaber, S.Z. Heparin-Induced Thrombocytopenia. Circulation 2006, 114, e355–e356. [Google Scholar] [CrossRef] [Green Version]
- Flexner, C.; Barditch-Crovo, P.A.; Kornhauser, D.M.; Farzadegan, H.; Nerhood, L.J.; Chaisson, R.E.; Bell, K.M.; Lorentsen, K.J.; Hendrix, C.W.; Petty, B.G.; et al. Pharmacokinetics, toxicity, and activity of intravenous dextran sulfate in human immunodeficiency virus infection. Antimicrob. Agents Chemother. 1991, 35, 2544–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hileman, R.E.; Fromm, J.R.; Weiler, J.M.; Linhardt, R.J. Glycosaminoglycan-protein interactions: Definition of consensus sites in glycosaminoglycan binding proteins. BioEssays 1998, 20, 156–167. [Google Scholar] [CrossRef]
- Pugliese, A.; Gennero, L.; Vidotto, V.; Beltramo, T.; Petrini, S.; Torre, D. A review of cardiovascular complications accompanying AIDS. Cell Biochem. Funct. 2004, 22, 137–141. [Google Scholar] [CrossRef]
- Fonseca, R.J.; Oliveira, S.-N.; Melo, F.R.; Pereira, M.G.; Benevides, N.M.B.; Mourão, P.A. Slight differences in sulfation of algal galactans account for differences in their anticoagulant and venous antithrombotic activities. Thromb. Haemost. 2008, 99, 539–545. [Google Scholar] [CrossRef]
- Ma, Q.; Dudas, B.; Hejna, M.; Cornelli, U.; Lee, J.M.; Lorens, S.; Mervis, R.; Hanin, I.; Fareed, J. The blood-barrier accessibility of a heparin-derived oligosaccharide C3. Thromb. Res. 2002, 105, 447–453. [Google Scholar] [CrossRef]
- Melo, F.R.; Pereira, M.S.; Foguel, D.; Mourão, P.A.S. Antithrombin-mediated anticoagulant activity of sulfated polysaccharides: Different mechanisms of heparin and sulfated galactans. J. Biol. Chem. 2004, 279, 20824–20835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farndale, R.W.; Buttle, D.J.; Barrett, A.J. Improved quantitation and discrimination of sulphated glycosaminoglycans by use of dimethylmethylene blue. Biochim. Biophys. Acta (BBA) Gen. Subj. 1986, 883, 173–177. [Google Scholar] [CrossRef]
- Zoepfl, M.; Dwivedi, R.; Taylor, M.C.; Pomin, V.H.; McVoy, M.A. Antiviral activities of four marine sulfated glycans againts adenovirus and human cytomegalovirus. Antivir. Res. 2021, 190, 105077. [Google Scholar] [CrossRef]
- Kim, S.; Hahn, Y.K.; Podhaizer, E.M.; McLane, V.D.; Zou, S.; Hauser, K.F.; Knapp, P.E. A central role for glial CCR5 in directing the neuropathological interactions of HIV-1 Tat and opiates. J. Neuroinflamm. 2018, 15, 285. [Google Scholar] [CrossRef] [Green Version]
- Paris, J.J.; Zou, S.; Hahn, Y.K.; Knapp, P.E.; Hauser, K.F. 5α-reduced progestogens ameliorate mood-related behavioral pathology, neurotoxicity, and microgliosis associated with exposure to HIV-1 Tat. Brain Behav. Immun. 2016, 55, 202–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris, J.J.; Liere, P.; Kim, S.; Mahdi, F.; Buchanan, M.E.; Nass, S.R.; Qrareya, A.N.; Salahuddin, M.F.; Pianos, A.; Fernandez, N.; et al. Pregnane steroidogenesis is altered by HIV-1 Tat and morphine: Physiological allopregnanolone is protective against neurotoxic and psychomotor effects. Neurobiol. Stress 2020, 12, 100211. [Google Scholar] [CrossRef]
- Salahuddin, M.F.; Qrareya, A.N.; Mahdi, F.; Jackson, D.; Foster, M.; Vujanovic, T.; Box, J.G.; Paris, J.J. Combined HIV-1 Tat and oxycodone activate the hypothalamic-pituitary-adrenal and -gonadal axes and promote psychomotor, affective, and cognitive dysfunction in female mice. Horm. Behav. 2020, 119, 104649. [Google Scholar] [CrossRef]
- Castro, M.O.; Pomin, V.H.; Santos, L.L.; Vilela-Silva, A.-C.E.S.; Hirohashi, N.; Pol-Fachin, L.; Verli, H.; Mourão, P.A.S. A Unique 2-Sulfated β-Galactan from the Egg Jelly of the Sea Urchin Glyptocidaris crenularis: Conformation flexibility versus induction of the sperm acrosome reaction. J. Biol. Chem. 2009, 284, 18790–18800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farias, E.H.C.; Pomin, V.H.; Valente, A.-P.; Nader, H.B.; Rocha, H.A.O.; Mourão, P.A.S. A preponderantly 4-sulfated, 3-linked galactan from the green alga Codium isthmocladum. Glycobiology 2008, 18, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interaction | ka (1/MS) | kd (1/S) | KD (M) |
|---|---|---|---|
| UFH + Tat | 48 ± 3.4 | 2.7 × 10−4 ± 2.0 × 10−5 | 5.7 × 10−6 |
| UFH + gp120 | 2.2 × 103 ± 64 | 1.9 × 10−3 ± 9.2 × 10−5 | 8.7 × 10−7 |
| BoSG + Tat | 1.4 × 104 ± 237 | 1.0 × 10−3 ± 1.1 × 10−4 | 7.5 × 10−8 |
| BoSG + gp120 | 3.1 × 103 ± 30 | 9.8 × 10−4 ± 5.9 × 10−5 | 3.2 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pomin, V.H.; Mahdi, F.; Jin, W.; Zhang, F.; Linhardt, R.J.; Paris, J.J. Red Algal Sulfated Galactan Binds and Protects Neural Cells from HIV-1 gp120 and Tat. Pharmaceuticals 2021, 14, 714. https://doi.org/10.3390/ph14080714
Pomin VH, Mahdi F, Jin W, Zhang F, Linhardt RJ, Paris JJ. Red Algal Sulfated Galactan Binds and Protects Neural Cells from HIV-1 gp120 and Tat. Pharmaceuticals. 2021; 14(8):714. https://doi.org/10.3390/ph14080714
Chicago/Turabian StylePomin, Vitor H., Fakhri Mahdi, Weihua Jin, Fuming Zhang, Robert J. Linhardt, and Jason J. Paris. 2021. "Red Algal Sulfated Galactan Binds and Protects Neural Cells from HIV-1 gp120 and Tat" Pharmaceuticals 14, no. 8: 714. https://doi.org/10.3390/ph14080714
APA StylePomin, V. H., Mahdi, F., Jin, W., Zhang, F., Linhardt, R. J., & Paris, J. J. (2021). Red Algal Sulfated Galactan Binds and Protects Neural Cells from HIV-1 gp120 and Tat. Pharmaceuticals, 14(8), 714. https://doi.org/10.3390/ph14080714