
Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model

 , and
, and
Abstract
:1. Introduction
2. Results
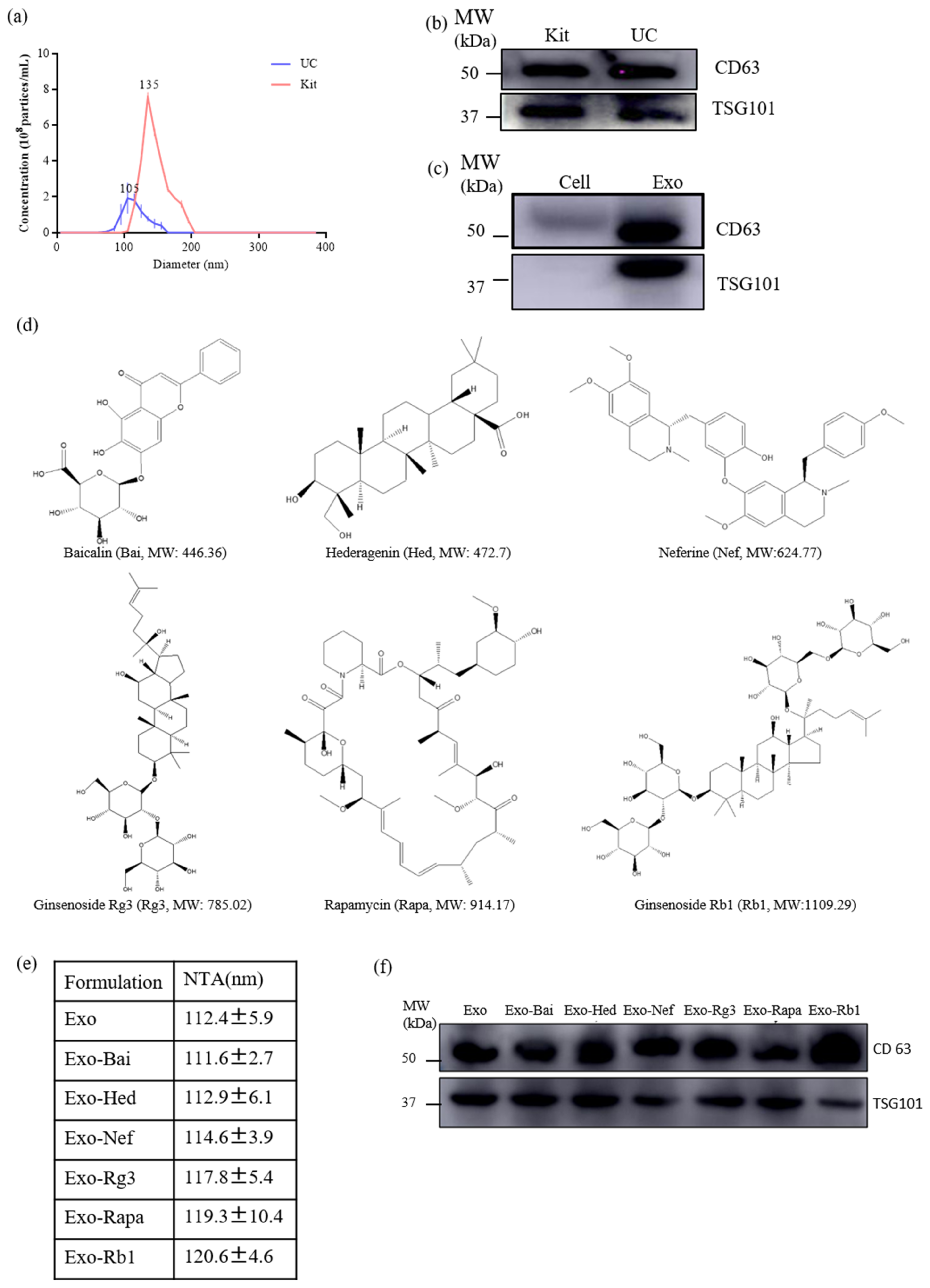
2.1. Characterization of Exosomes and Exosomal Encapsulated-Compounds
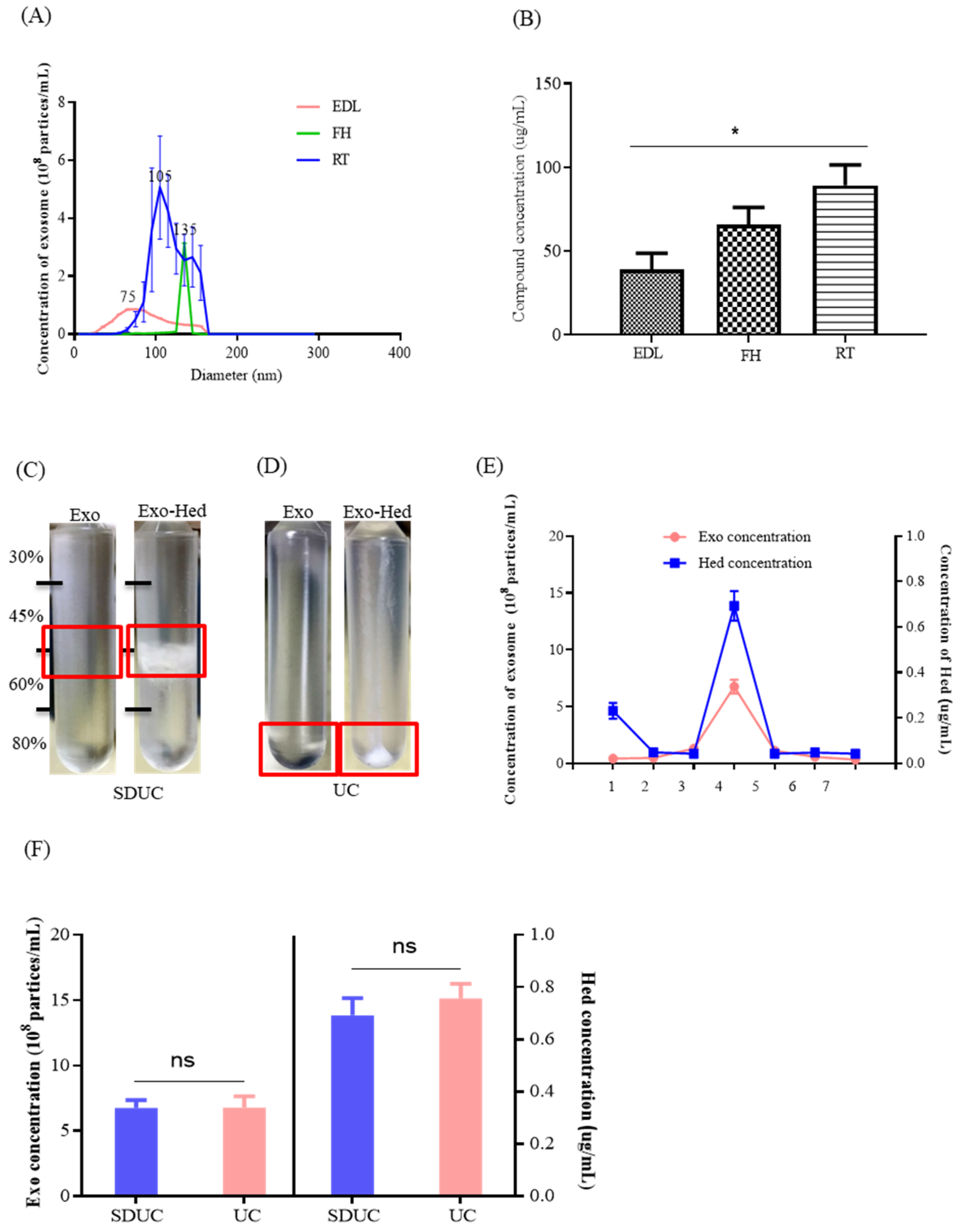
2.2. Optimization of the Compound Loading Methods for the Exosomes
2.3. Blood–Brain Barrier Permeability of Exosomal-Encapsulated Compounds with Increasing Molecular Weights
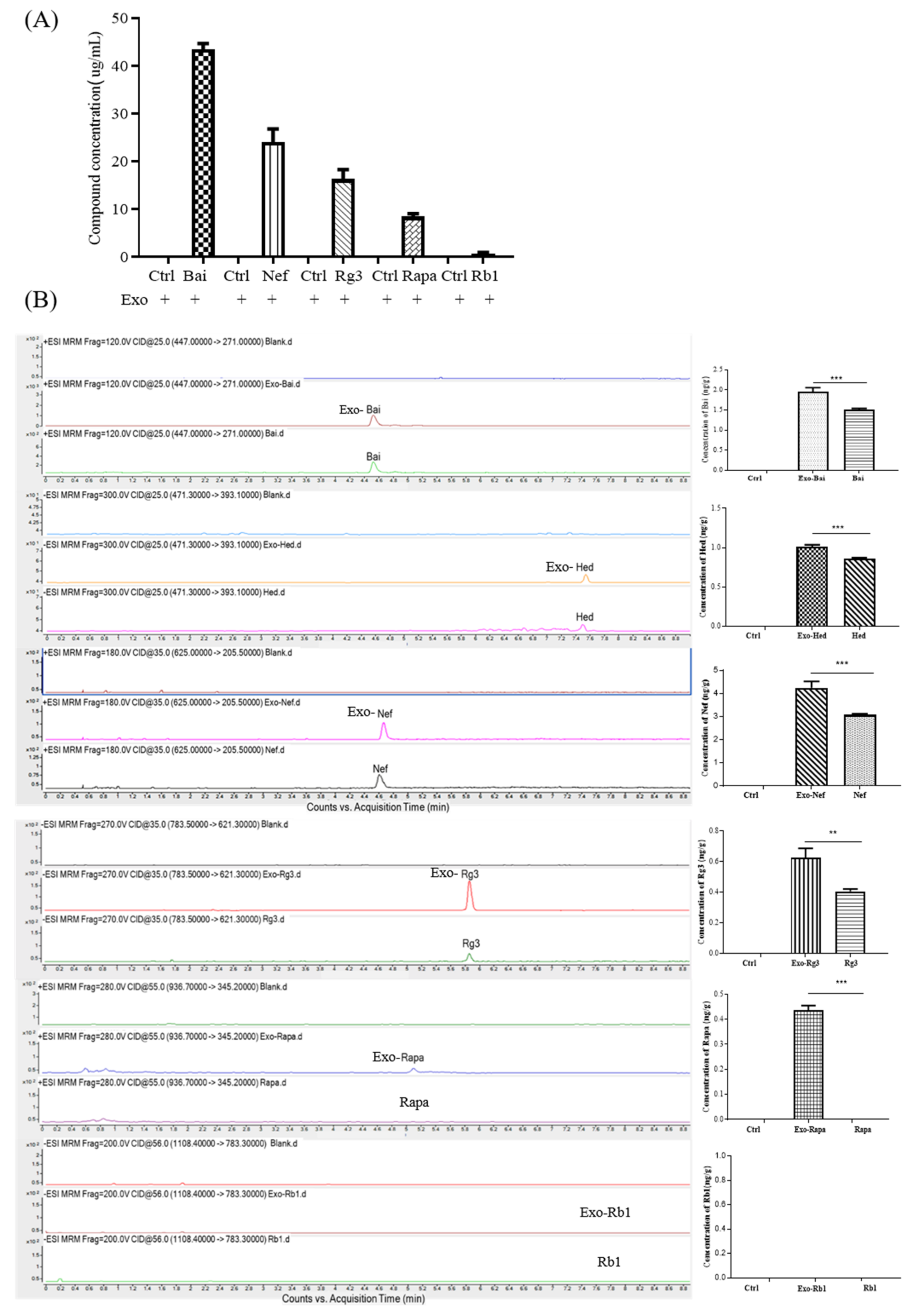
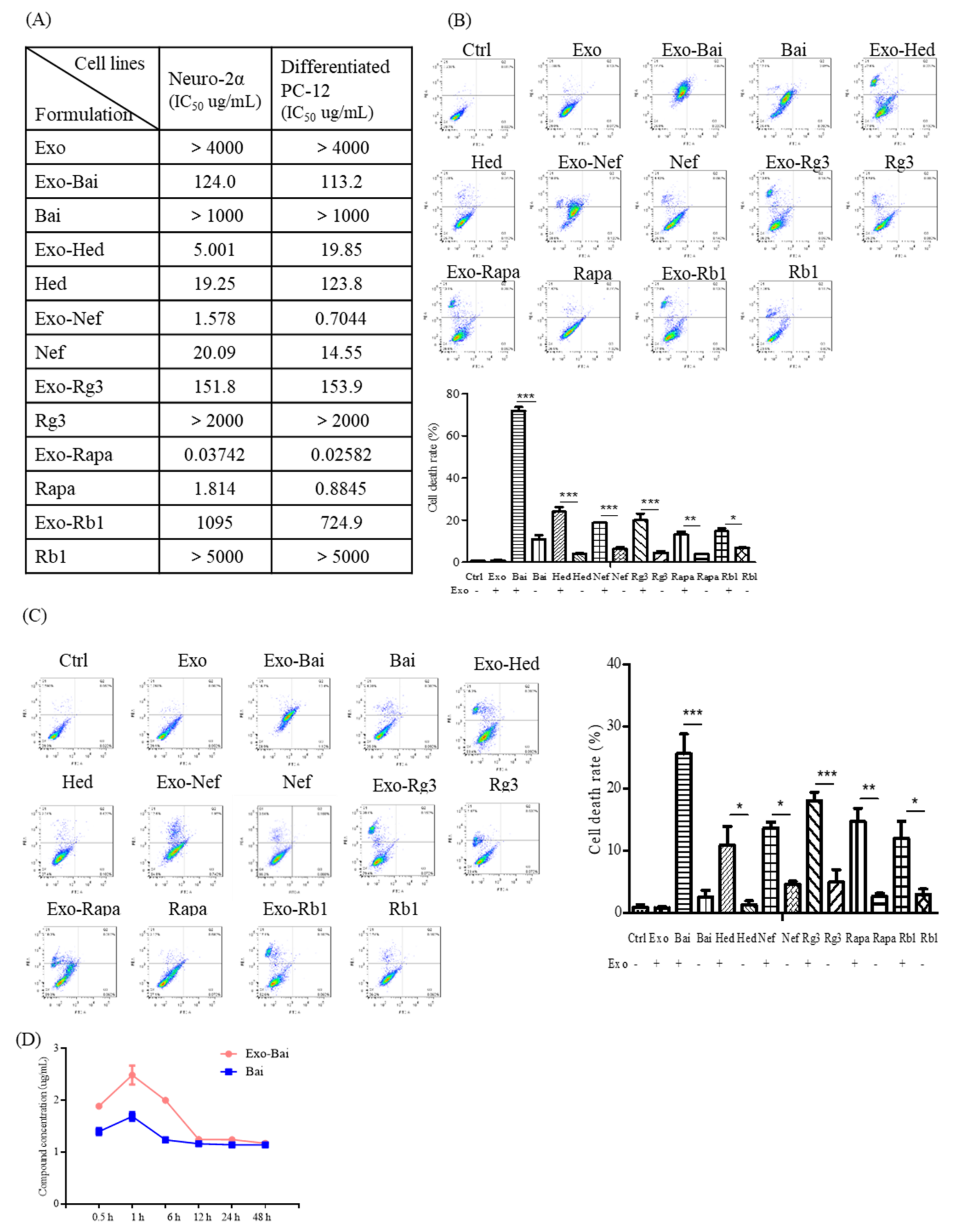
2.4. Bio-Availability and Cytotoxicity of Exosomal Compounds in the Cellular System
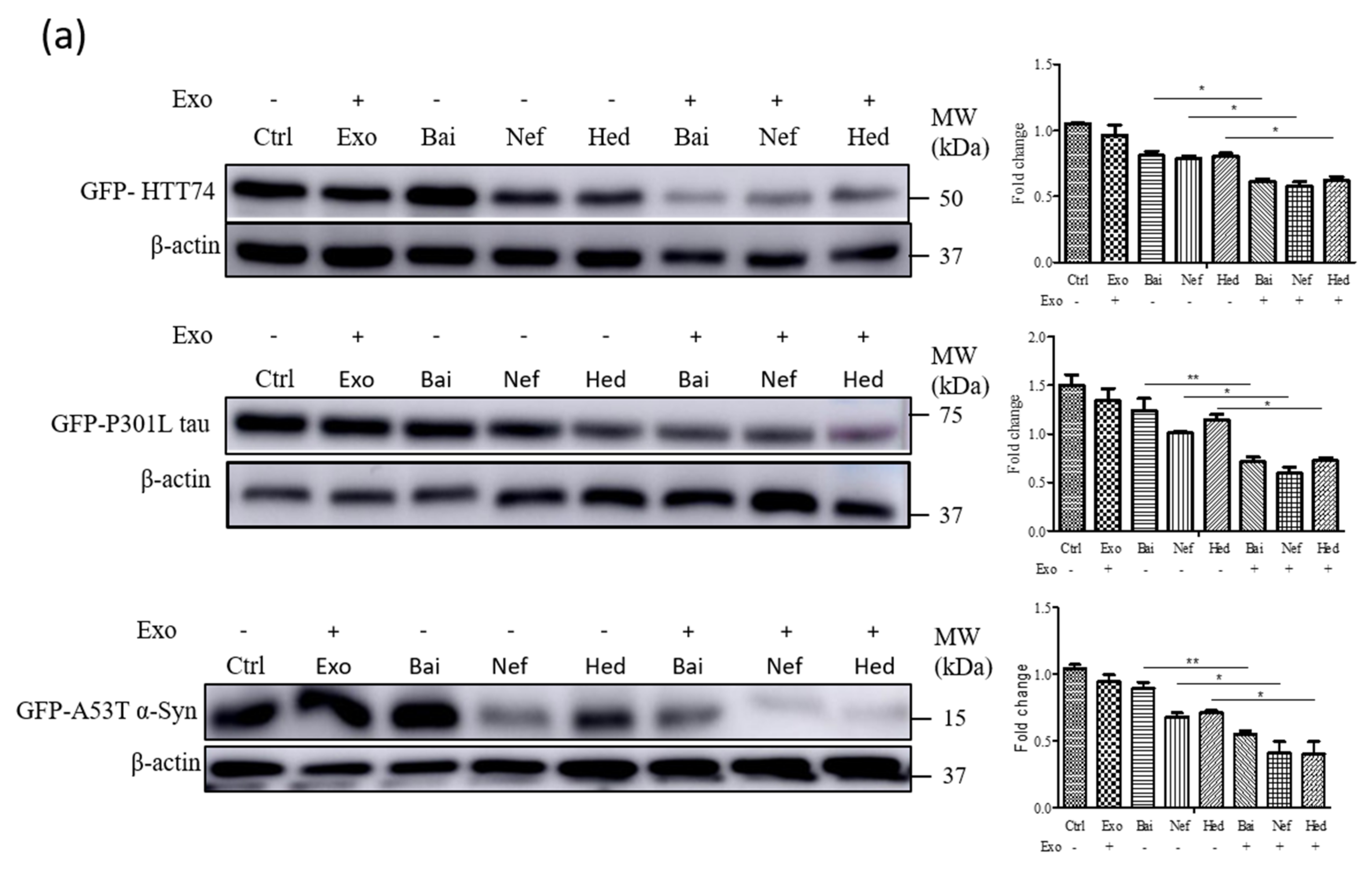
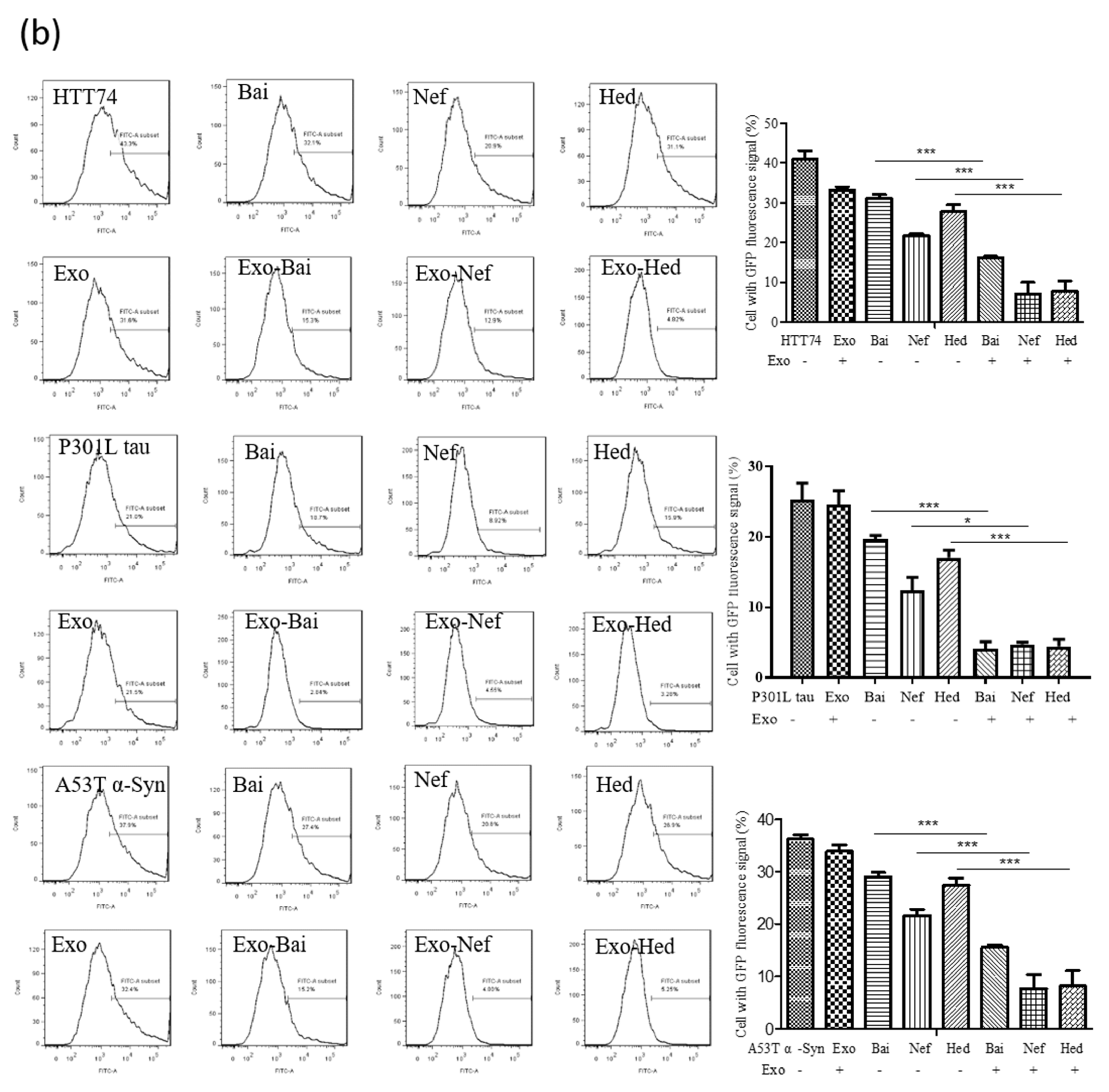
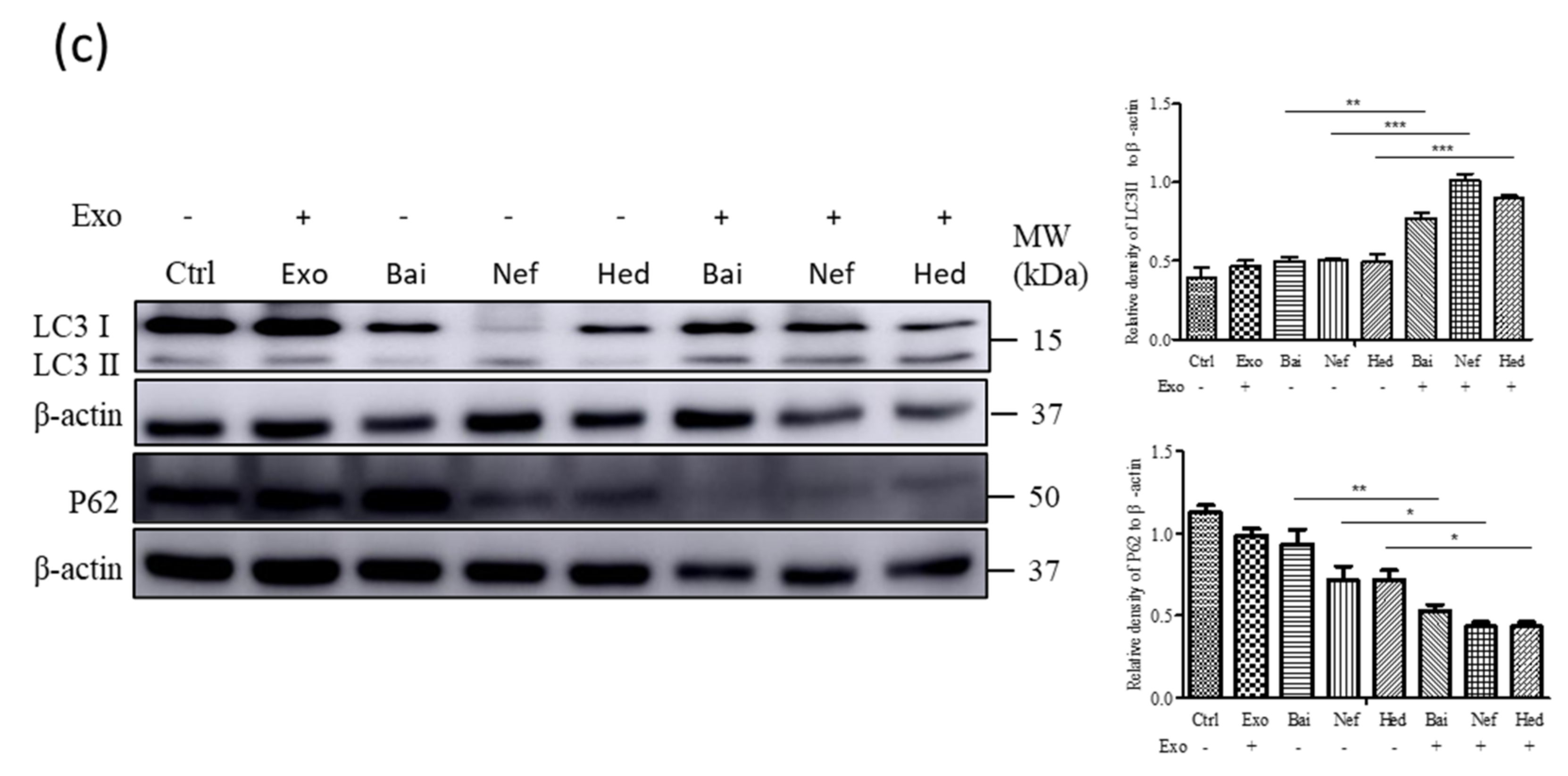
2.5. Exosomal Compounds Attenuate the Cellular Level of Neurodegenerative Disease Proteins
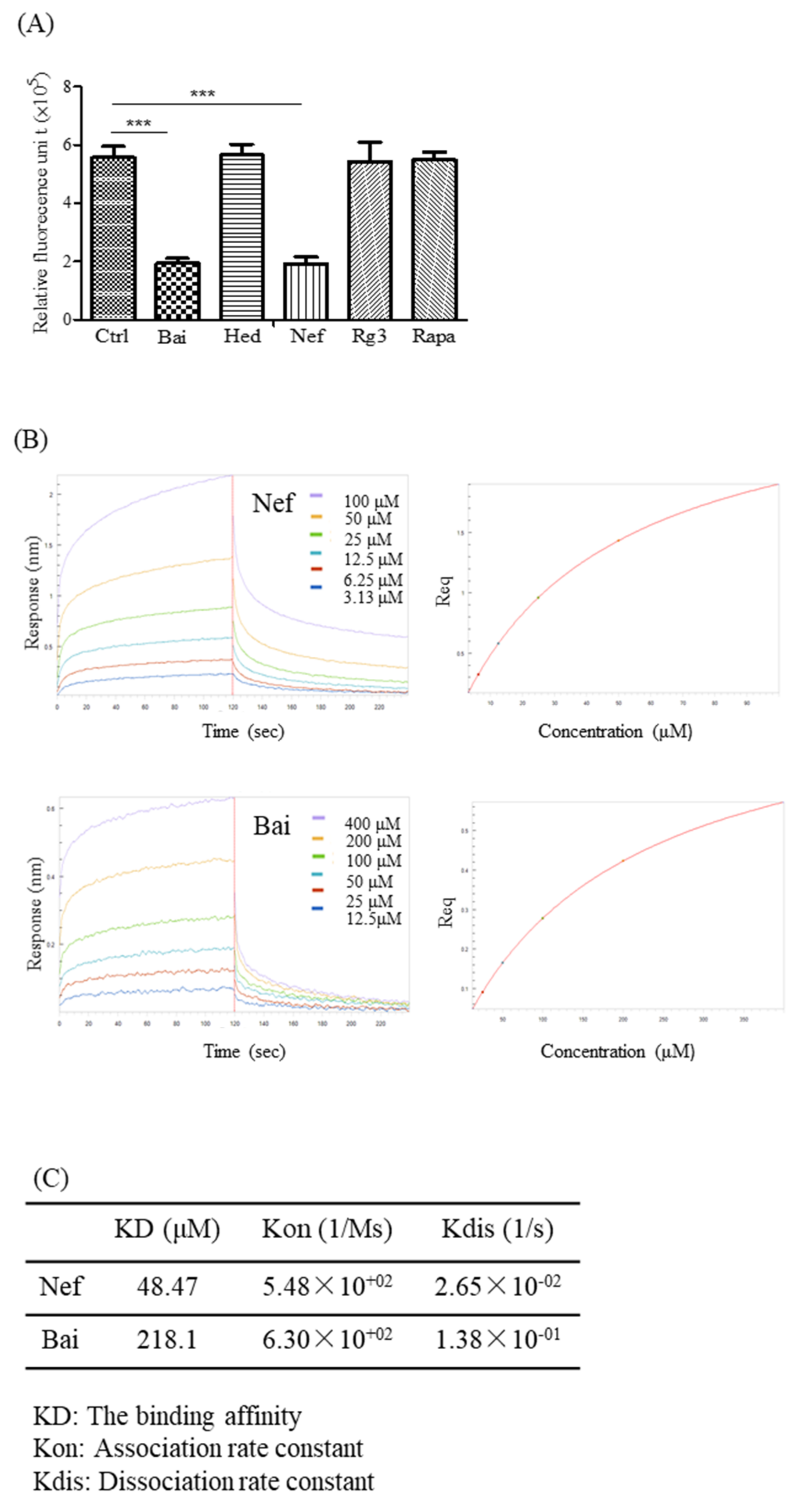
2.6. Validation of the Aβ-Binding Propensity of the Selected Compounds by ThT and BLI
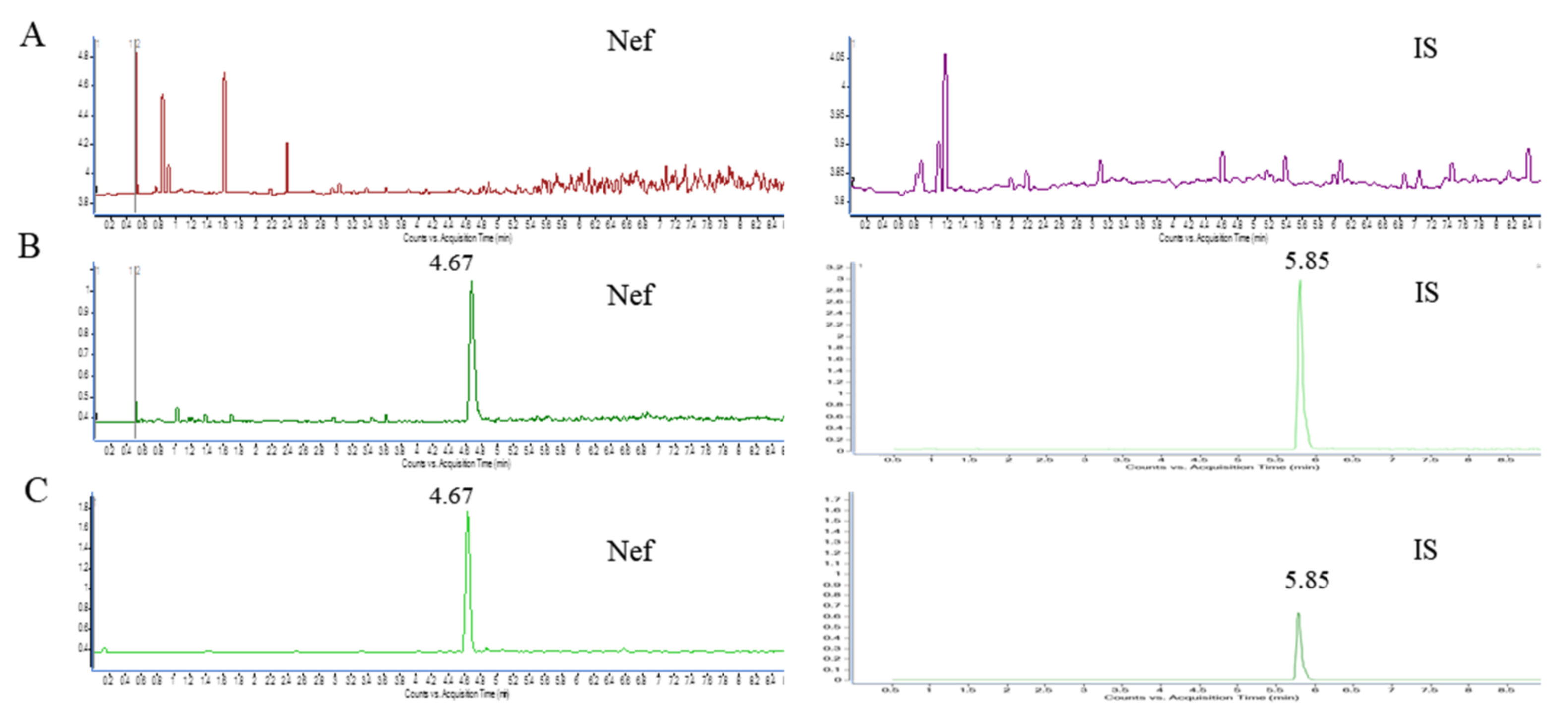
2.7. The Validation of the UHPLC-MS Analytical Method
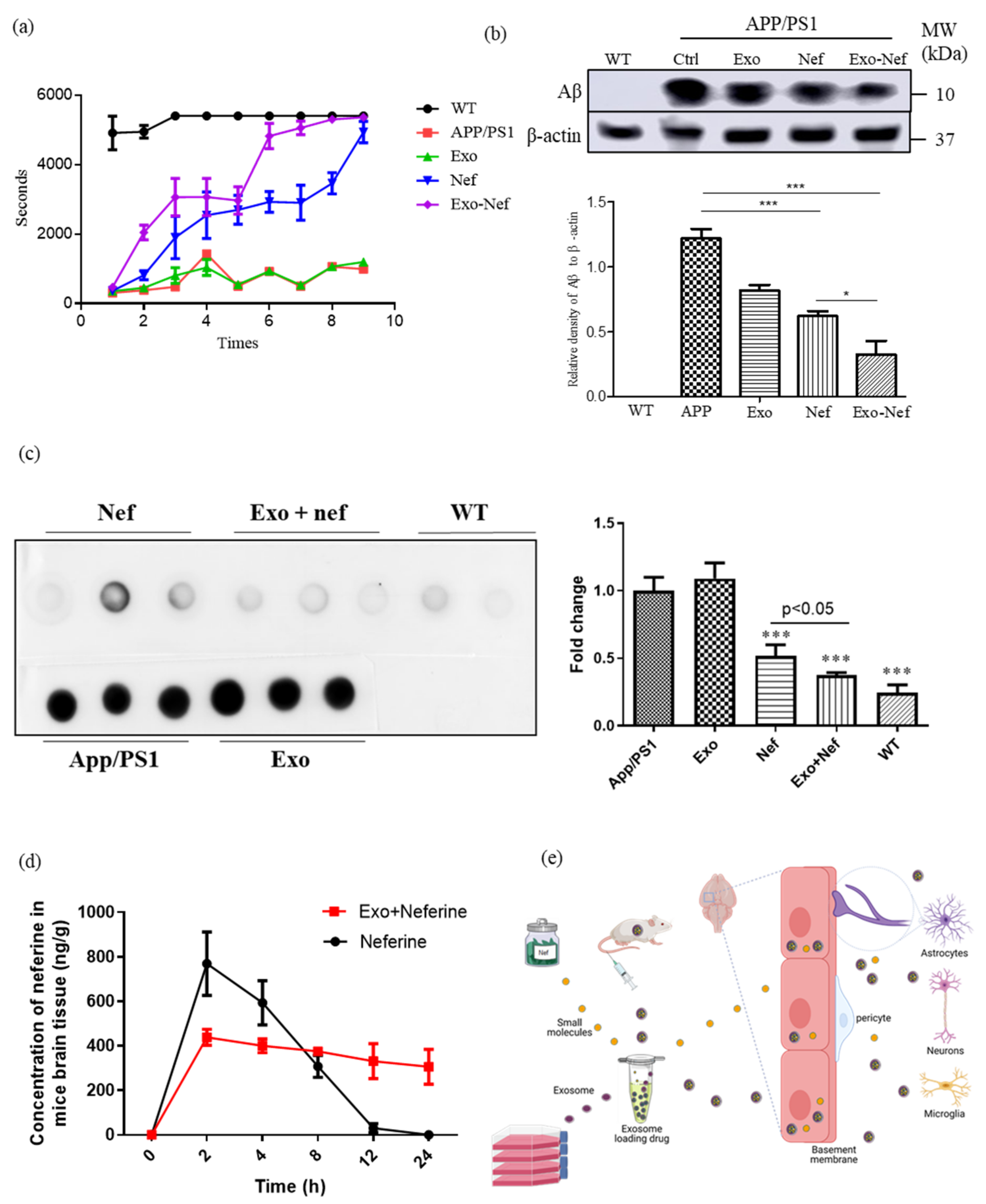
2.8. Exo-Nef Improves the Motor Deficiency of APP/PS1 Mice, and Reduces Aβ Deposition in Mouse Brains
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of the Exosomes
4.3. Preparation of the Exosome-Encapsulated Compounds
4.4. Nanoparticle Tracking Analysis (NTA)
4.5. Analysis of the Concentration of the Exosomal Compounds in the Brain Tissue of the Mice
4.6. Cell Viability Analysis
4.7. Flow Cytometry Analysis
4.8. Western Blot Analysis
4.9. Thioflavin-T (ThT) Fluorescence Assay
4.10. Dot Blot Analysis
4.11. Biolayer Interferometry Analysis
4.12. Animals
4.13. Rotarod Behavioral Test
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, B.-T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Witwer, K.W.; Théry, C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell. Vesicles 2019, 8, 1648167. [Google Scholar] [CrossRef]
- Plebanek, M.; Angeloni, N.; Vinokour, E.; Dalia, M.-M.; Henkin, A.; Martinez-Marin, D.; Filleur, S.; Bhowmick, R.; Henkin, J.; Miller, S.D.; et al. Pre-metastatic cancer exosomes induce immune surveillance by patrolling monocytes at the metastatic niche. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Budnik, V.; Ruiz-Cañada, C.; Wendler, V.B.C.R.-C.F. Extracellular vesicles round off communication in the nervous system. Nat. Rev. Neurosci. 2016, 17, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-X.; Sun, C.; Wang, L.; Guo, X.-L. New insight into isolation, identification techniques and medical applications of exosomes. J. Control. Release 2019, 308, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Jan, L.; Andrew, F.H.; Fred, H.; Edit, I.B.; Dolores Di, V.; Christopher, G.; Yong Song, G.; Igor, V.K.; Suresh, M.; Peter, Q.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the international society for extracellular vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar]
- Witwer, K.W.; Soekmadji, C.; Hill, A.F.; Wauben, M.H.; Buzás, E.I.; Di Vizio, D.; Falcon-Perez, J.M.; Gardiner, C.; Hochberg, F.; Kurochkin, I.V.; et al. Updating the MISEV minimal requirements for extracellular vesicle studies: Building bridges to reproducibility. J. Extracell. Vesicles 2017, 6, 1396823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taverna, S.; Giallombardo, M.; Gil-Bazo, I.; Carreca, A.P.; Castiglia, M.; Chacártegui, J.; Araújo, A.; Alessandro, R.; Pauwels, P.; Peeters, M.; et al. Exosomes isolation and characterization in serum is feasible in non-small cell lung cancer patients: Critical analysis of evidence and potential role in clinical practice. Oncotarget 2016, 7, 28748–28760. [Google Scholar] [CrossRef]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome Delivered Anticancer Drugs Across the Blood-Brain Barrier for Brain Cancer Therapy in Danio Rerio. Pharm. Res. 2015, 32, 2003–2014. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [Green Version]
- Vella, L.J.; Scicluna, B.J.; Cheng, L.; Bawden, E.G.; Masters, C.; Ang, C.-S.; Williamson, N.; McLean, C.; Barnham, K.J.; Hill, A.F. A rigorous method to enrich for exosomes from brain tissue. J. Extracell. Vesicles 2017, 6, 1348885. [Google Scholar] [CrossRef]
- Edgar, J.R. Q&a: What are exosomes, exactly? BMC Biol. 2016, 14, 7. [Google Scholar]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Sun, D.; Zhuang, X.; Xiang, X.; Liu, Y.; Zhang, S.; Liu, C.; Barnes, S.; Grizzle, W.; Miller, D.; Zhang, H.-G. A Novel Nanoparticle Drug Delivery System: The Anti-inflammatory Activity of Curcumin Is Enhanced When Encapsulated in Exosomes. Mol. Ther. 2010, 18, 1606–1614. [Google Scholar] [CrossRef]
- Saari, H.O.; Lázaro-Ibáñez, E.; Viitala, T.; Vuorimaa-Laukkanen, E.; Siljander, P.; Yliperttula, M. Microvesicle- and exosome-mediated drug delivery enhances the cytotoxicity of Paclitaxel in autologous prostate cancer cells. J. Control. Release 2015, 220, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Pardridge, W.M. Brain Drug Targeting: The Future of Brain Drug Development; Cambridge University Press: Cambridge, UK, 2011; pp. 1–347. [Google Scholar]
- Pardridge, W.M. Drug transport across the blood–brain barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.J.; O’Loughlin, A.J.; Lakhal, S. Exosomes and the blood–brain barrier: Implications for neurological diseases. Ther. Deliv. 2011, 2, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Pascucci, L.; Coccè, V.; Bonomi, A.; Ami, D.; Ceccarelli, P.; Ciusani, E.; Viganò, L.; Locatelli, A.; Sisto, F.; Doglia, S.M.; et al. Paclitaxel is incorporated by mesenchymal stromal cells and released in exosomes that inhibit in vitro tumor growth: A new approach for drug delivery. J. Control. Release 2014, 192, 262–270. [Google Scholar] [CrossRef]
- Fuhrmann, G.; Serio, A.; Mazo, M.M.; Nair, R.; Stevens, M.M. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J. Control. Release 2015, 205, 35–44. [Google Scholar] [CrossRef]
- Yu, L.; Wu, A.-G.; Wong, V.K.-W.; Qu, L.-Q.; Zhang, N.; Qin, D.-L.; Zeng, W.; Tang, B.; Wang, H.-M.; Wang, Q.; et al. The new application of uhplc-dad-tof/ms in identification of inhibitors on β-amyloid fibrillation from. Front. Pharmacol. 2019, 10, 194. [Google Scholar] [CrossRef]
- Wu, A.-G.; Zeng, W.; Wong, V.K.-W.; Zhu, Y.-Z.; Lo, A.C.; Liu, L.; Law, B.Y.-K. Hederagenin and α-hederin promote degradation of proteins in neurodegenerative diseases and improve motor deficits in MPTP-mice. Pharmacol. Res. 2016, 115, 25–44. [Google Scholar] [CrossRef]
- Wong, V.K.W.; Wu, A.G.; Wang, J.R.; Liu, L.; Law, B.Y.-K. Neferine Attenuates the Protein Level and Toxicity of Mutant Huntingtin in PC-12 Cells via Induction of Autophagy. Molecules 2015, 20, 3496–3514. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wang, T.; Li, C.; Wang, Z.; Zhao, Y.; He, J.; Fu, L.; Han, B. Ginsenoside Rg3 exerts a neuroprotective effect in rotenone-induced Parkinson’s disease mice via its anti-oxidative properties. Eur. J. Pharmacol. 2021, 909, 174413. [Google Scholar] [CrossRef]
- Song, Y.; Xue, H.; Liu, T.-T.; Liu, J.-M.; Chen, D. Rapamycin Plays a Neuroprotective Effect after Spinal Cord Injury via Anti-Inflammatory Effects. J. Biochem. Mol. Toxicol. 2014, 29, 29–34. [Google Scholar] [CrossRef]
- Qu, S.; Meng, X.; Liu, Y.; Zhang, X.; Zhang, Y. Ginsenoside Rb1 prevents MPTP-induced changes in hippocampal memory via regulation of the α-synuclein/PSD-95 pathway. Aging 2019, 11, 1934–1964. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Liu, Y.; Zhang, C. Pharmacokinetics and Bioavailability Enhancement of Baicalin: A Review. Eur. J. Drug Metab. Pharmacokinet. 2018, 44, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-J.; Cheng, J.-J.; Liao, C.-C.; Cheng, H.-L.; Huang, H.-T.; Kuo, L.-M.Y.; Kuo, Y.-H. Triterpene Acids from Euscaphis japonica and Assessment of Their Cytotoxic and Anti-NO Activities. Planta Med. 2012, 78, 1584–1590. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.-H.; Guan, J.-H.; Huang, Y.-L.; Pan, Y.-W.; Yang, W.; Lan, H.; Huang, S.; Hu, J.; Zhao, G. Experimental Study of Antiatherosclerosis Effects with Hederagenin in Rats. Evidence-Based Complement. Altern. Med. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Mariappan, R.; Muthusamy, S.; Velmurugan, B.K. Pharmacological benefits of neferine—A comprehensive review. Life Sci. 2018, 199, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, T.; Li, W.; Li, J.; Wang, C.; Zhang, K. Insights into the antitumor mechanism of ginsenosides Rg3. Mol. Biol. Rep. 2021, 48, 2639–2652. [Google Scholar] [CrossRef]
- Maiese, K. The Mechanistic Target of Rapamycin (mTOR): Novel Considerations as an Antiviral Treatment. Curr. Neurovascular Res. 2020, 17, 332–337. [Google Scholar] [CrossRef]
- Yoo, Y.J.; Kim, H.; Park, S.R.; Yoon, Y.J. An overview of rapamycin: From discovery to future perspectives. J. Ind. Microbiol. Biotechnol. 2016, 44, 537–553. [Google Scholar] [CrossRef]
- Zheng, Q.; Bao, X.-Y.; Zhu, P.-C.; Tong, Q.; Zheng, G.-Q.; Wang, Y. Ginsenoside Rb1 for Myocardial Ischemia/Reperfusion Injury: Preclinical Evidence and Possible Mechanisms. Oxidative Med. Cell. Longev. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Kong, H.; Sun, Y.; Feng, H.; Zhang, Y.; Su, X.; Qu, H.; Wang, Q. Assessment of baicalin in mouse blood by monoclonal antibody-based icELISA. Biomed. Chromatogr. 2014, 28, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jing, F.-B.; Zhang, Z. Development and validation of a quantification method for oleanolic acid and hederagenin in rat plasma: Application to the pharmacokinetic study. Biomed. Chromatogr. 2016, 31, e3801. [Google Scholar] [CrossRef]
- Zhang, H.X.; Xing, Y. Study on the distribution of neferine in the brain tissue of mice. China Pharm. 2010, 1, 3. [Google Scholar]
- Xie, H.-T.; Wang, G.-J.; Sun, J.-G.; Tucker, I.; Zhao, X.-C.; Xie, Y.-Y.; Li, H.; Jiang, X.-L.; Wang, R.; Xu, M.-J. High performance liquid chromatographic–mass spectrometric determination of ginsenoside Rg3 and its metabolites in rat plasma using solid-phase extraction for pharmacokinetic studies. J. Chromatogr. B 2005, 818, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Trepanier, D.J.; Gallant, H.; Legatt, D.F.; Yatscoff, R.W. Rapamycin: Distribution, pharmacokinetics and therapeutic range investigations: An update. Clin. Biochem. 1998, 31, 345–351. [Google Scholar] [CrossRef]
- Xu, Q.F.; Fang, X.L.; Chen, D.F. Pharmacokinetics and bioavailability of ginsenoside Rb1 and Rg1 from Panax notoginseng in rats. J. Ethnopharmacol. 2003, 84, 187–192. [Google Scholar] [CrossRef]
- Li, K. Pharmacokinetics of Diosgenin and Ginsenoside rg3 in Animals. Ph.D. Thesis, Shenyang Pharmaceutical University, Shenyang, China, 2005. [Google Scholar]
- Lu, J.H.; Ardah, M.T.; Durairajan, S.S.K.; Liu, L.F.; Xie, L.X.; Fong, W.F.D.; Hasan, M.Y.; Huang, J.D.; El-Agnaf, O.M.A.; Li, M. Baicalein inhibits formation of alpha-synuclein oligomers within living cells and prevents a beta peptide fibrillation and oligomerisation. Chembiochem 2011, 12, 615–624. [Google Scholar] [CrossRef]
- Zhu, X.; Yao, P.; Liu, J.; Guo, X.; Jiang, C.; Tang, Y. Baicalein attenuates impairment of hepatic lysosomal acidification induced by high fat diet via maintaining V-ATPase assembly. Food Chem. Toxicol. 2020, 136, 110990. [Google Scholar] [CrossRef] [PubMed]
- Mathew, B.; Chennakesavalu, M.; Sharma, M.; Torres, L.A.; Stelman, C.R.; Tran, S.; Patel, R.; Burg, N.; Salkovski, M.; Kadzielawa, K.; et al. Autophagy and post-ischemic conditioning in retinal ischemia. Autophagy 2020, 17, 1479–1499. [Google Scholar] [CrossRef]
- Nisbet, R.M.; Götz, J. Amyloid-β and Tau in Alzheimer’s Disease: Novel Pathomechanisms and Non-Pharmacological Treatment Strategies. J. Alzheimer’s Dis. 2018, 64, S517–S527. [Google Scholar] [CrossRef]
- Xue, C.; Lin, T.Y.; Chang, D.; Guo, Z. Thioflavin T as an amyloid dye: Fibril quantification, optimal concentration and effect on aggregation. R. Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [Green Version]
- Batrakova, E.V.; Kim, M. Using exosomes, naturally-equipped nanocarriers, for drug delivery. J. Control. Release 2015, 219, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, C.K.; Jena, B.C.; Banerjee, I.; Das, S.; Parekh, A.; Bhutia, S.K.; Mandal, M. Exosome as a Novel Shuttle for Delivery of Therapeutics across Biological Barriers. Mol. Pharm. 2018, 16, 24–40. [Google Scholar] [CrossRef]
- Sidhaye, J.; Knoblich, J.A. Brain organoids: An ensemble of bioassays to investigate human neurodevelopment and disease. Cell Death Differ. 2020, 28, 52–67. [Google Scholar] [CrossRef]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnology 2020, 18, 1–15. [Google Scholar] [CrossRef]
- Usman, W.M.; Pham, T.C.; Kwok, Y.Y.; Vu, L.T.; Ma, V.; Peng, B.; Chan, Y.S.; Wei, L.; Chin, S.M.; Azad, A.; et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat. Commun. 2018, 9, 2359. [Google Scholar] [CrossRef]
- Sun, W.; Xing, C.; Zhao, L.; Zhao, P.; Yang, G.; Yuan, L. Ultrasound Assisted Exosomal Delivery of Tissue Responsive mRNA for Enhanced Efficacy and Minimized Off-Target Effects. Mol. Ther. Nucleic Acids 2020, 20, 558–567. [Google Scholar] [CrossRef]
- Xu, M.; Feng, T.; Liu, B.; Qiu, F.; Xu, Y.; Zhao, Y.; Zheng, Y. Engineered exosomes: Desirable target-tracking characteristics for cerebrovascular and neurodegenerative disease therapies. Theranostics 2021, 11, 8926–8944. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Usuki, S.; Sakai, S.; Hanamatsu, H.; Mioka, T.; Kimura, N.; Okada, M.; Tahara, H.; Furukawa, J.-I.; et al. A potential function for neuronal exosomes: Sequestering intracerebral amyloid-β peptide. FEBS Lett. 2014, 589, 84–88. [Google Scholar] [CrossRef]
- Bahrini, I.; Song, J.-H.; Diez, D.; Hanayama, R. Neuronal exosomes facilitate synaptic pruning by up-regulating complement factors in microglia. Sci. Rep. 2015, 5, 7989. [Google Scholar] [CrossRef] [Green Version]
- Abdulrahman, B.; Abdelaziz, D.H.; Schatzl, H.M. Autophagy regulates exosomal release of prions in neuronal cells. J. Biol. Chem. 2018, 293, 8956–8968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Xu, Y.; Deng, W.; Zhang, L.; Zhu, H.; Yu, P.; Qu, Y.; Zhao, W.; Han, Y.; Qin, C. Brain Derived Exosomes Are a Double-Edged Sword in Alzheimer’s Disease. Front. Mol. Neurosci. 2020, 13, 79. [Google Scholar] [CrossRef]
- Yeo, R.W.Y.; Lai, R.C.; Zhang, B.; Tan, S.S.; Yin, Y.; Teh, B.J.; Lim, S.K. Mesenchymal stem cell: An efficient mass producer of exosomes for drug delivery. Adv. Drug Deliv. Rev. 2013, 65, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, P.; Pizzuti, V.; Zia, S.; Sargenti, A.; Gazzola, D.; Roda, B.; Bonsi, L.; Alviano, F. Microfluidic Tools for Enhanced Characterization of Therapeutic Stem Cells and Prediction of Their Potential Antimicrobial Secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef]
- Yaghoubi, Y.; Movassaghpour, A.; Zamani, M.; Talebi, M.; Mehdizadeh, A.; Yousefi, M. Human umbilical cord mesenchymal stem cells derived-exosomes in diseases treatment. Life Sci. 2019, 233, 116733. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Aoki, N.; Tominaga, N.; Takeshita, F.; Sonoda, H.; Yoshioka, Y.; Ochiya, T. Disruption of Circulating Extracellular Vesicles as a Novel Therapeutic Strategy against Cancer Metastasis. Mol. Ther. 2017, 25, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Betzer, O.; Perets, N.; Angel, A.; Motiei, M.; Sadan, T.; Yadid, G.; Offen, D.; Popovtzer, R. In Vivo Neuroimaging of Exosomes Using Gold Nanoparticles. ACS Nano 2017, 11, 10883–10893. [Google Scholar] [CrossRef] [PubMed]
- FDA, U. Guidance for Industry: Bioanalytical Method Validation; US Department of Health and Human Services, Food and Drug Administration: Center for Drug Evaluation and Research: Rockville, MD, USA, 2001. [Google Scholar]
- Zhang, T.-T.; Wang, Y.-L.; Jin, B.; Li, T.; Ma, C. Plasma pharmacokinetics of isorhapontigenin, a novel derivative of stilbenes, in mice by lc-ms/ms method. J. Asian Nat. Prod. Res. 2019, 21, 895–904. [Google Scholar] [CrossRef]
- Wong, V.K.; Chiu, P.; Chung, S.S.; Chow, L.; Zhao, Y.-Z.; Yang, B.B.; Ko, B.C.B. Pseudolaric Acid B, a Novel Microtubule-Destabilizing Agent That Circumvents Multidrug Resistance Phenotype and Exhibits Antitumor Activity In vivo. Clin. Cancer Res. 2005, 11, 6002–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, M.; Rubinstein, M.; Phillips, T.J.; Lessov, C.N.; Burkhart-Kasch, S.; Zhang, G.; Bunzow, J.R.; Fang, Y.; Gerhardt, G.A.; Grandy, D.K.; et al. Locomotor Activity in D2 Dopamine Receptor-Deficient Mice Is Determined by Gene Dosage, Genetic Background, and Developmental Adaptations. J. Neurosci. 1998, 18, 3470–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iancu, R.; Mohapel, P.; Brundin, P.; Paul, G. Behavioral characterization of a unilateral 6-OHDA-lesion model of Parkinson’s disease in mice. Behav. Brain Res. 2005, 162, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rozas, G.; García, J.L. Drug-free evaluation of rat models of parkinsonism and nigral grafts using a new automated rotarod test. Brain Res. 1997, 749, 188–199. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Natural Source | Pharmacological Activity | References |
|---|---|---|---|
| Baicalin | Scutellaria baicalensis Georgi (root) | anti-tumor, anti-microbial, and anti-oxidative effects | [29] |
| Hederagenin | Hedera nepalensisvar K.Koch var.sinensis (whole plant) | anti-tumor, anti-apoptosis, and anti-inflammatory effects | [30,31] |
| Neferine | Nelumbo nuci fera Gaertn (seed) | anti-cancer, anti-diabetic, anti-aging, anti-microbial, anti-thrombotic, anti-arrhythmic and anti-inflammatory effects | [32] |
| Ginsenoside Rg3 | Panax ginseng C. A. Meyer (root) | anti-tumor effect | [33] |
| Rapamycin | Streptomyces hygroscopicus | anti-viral, anti-fungal, anti-tumor and neuroprotective effects | [34,35] |
| Ginsenoside Rb1 | Panax ginseng C. A. Meyer (root) | anti-oxidant, anti-inflammatory, anti-apoptosis and an anti-arrhythmic effects | [36] |
| Compounds | Dose(μg/kg) | Sacrificed Time * | References |
|---|---|---|---|
| Bai/Exo-Bai | 100 | 4 h | [37] |
| Hed/Exo-Hed | 100 | 50 min | [38] |
| Nef/Exo-Nef | 100 | 20 min | [39] |
| Rg3/Exo-Rg3 | 100 | 40 min | [43] |
| Rapa/Exo-Rapa | 100 | 3 h | [41] |
| Rb1/Exo-Rb1 | 100 | 1.5 h | [42] |
| Compound | Regression Equation | Correlation Coefficient | Detection Limit | Quantitation Limit |
|---|---|---|---|---|
| Y = aX + b | (ng/mL) | (ng/mL) | ||
| Nef | Y = 379.92X-764.77 | 0.9979 | 2.5 | 100 |
| Compound | Concentration(ng/mL) | Intra-Day | Inter-Day | ||
|---|---|---|---|---|---|
| Accuracy (%) | RSD (%) | Accuracy (%) | RSD (%) | ||
| Low | 2.5 | 14.3 | 2.4 | 5.7 | 14.2 |
| Medium | 25.0 | 7.3 | 1.2 | 6.7 | 8.4 |
| High | 100.0 | 5.3 | 8.5 | 2.0 | 14.2 |
| Subclass | IS | Spike (ng/mL) | Matrix Effect (Mean ± SD%) | RSD (%) | Extraction Recovery (Mean ± SD%) | RSD (%) | |
|---|---|---|---|---|---|---|---|
| Nef | Resv | Low | 2.5 | 97.2 ± 10.3 | 10.6 | 104.0 ± 2.1 | 8.1 |
| Medium | 25.0 | 100.9 ± 7.3 | 7.3 | 85.6 ± 3.5 | 1.2 | ||
| High | 100.0 | 100.6 ± 14.3 | 14.2 | 95.6 ± 5.7 | 2.5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, B.; Zeng, W.; Song, L.L.; Wang, H.M.; Qu, L.Q.; Lo, H.H.; Yu, L.; Wu, A.G.; Wong, V.K.W.; Law, B.Y.K. Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model. Pharmaceuticals 2022, 15, 83. https://doi.org/10.3390/ph15010083
Tang B, Zeng W, Song LL, Wang HM, Qu LQ, Lo HH, Yu L, Wu AG, Wong VKW, Law BYK. Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model. Pharmaceuticals. 2022; 15(1):83. https://doi.org/10.3390/ph15010083
Chicago/Turabian StyleTang, Bin, Wu Zeng, Lin Lin Song, Hui Miao Wang, Li Qun Qu, Hang Hong Lo, Lu Yu, An Guo Wu, Vincent Kam Wai Wong, and Betty Yuen Kwan Law. 2022. "Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model" Pharmaceuticals 15, no. 1: 83. https://doi.org/10.3390/ph15010083
APA StyleTang, B., Zeng, W., Song, L. L., Wang, H. M., Qu, L. Q., Lo, H. H., Yu, L., Wu, A. G., Wong, V. K. W., & Law, B. Y. K. (2022). Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model. Pharmaceuticals, 15(1), 83. https://doi.org/10.3390/ph15010083