

Optimization of Zebrafish Larvae Sectioning for Mass Spectrometry Imaging
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Considerations of Embedding
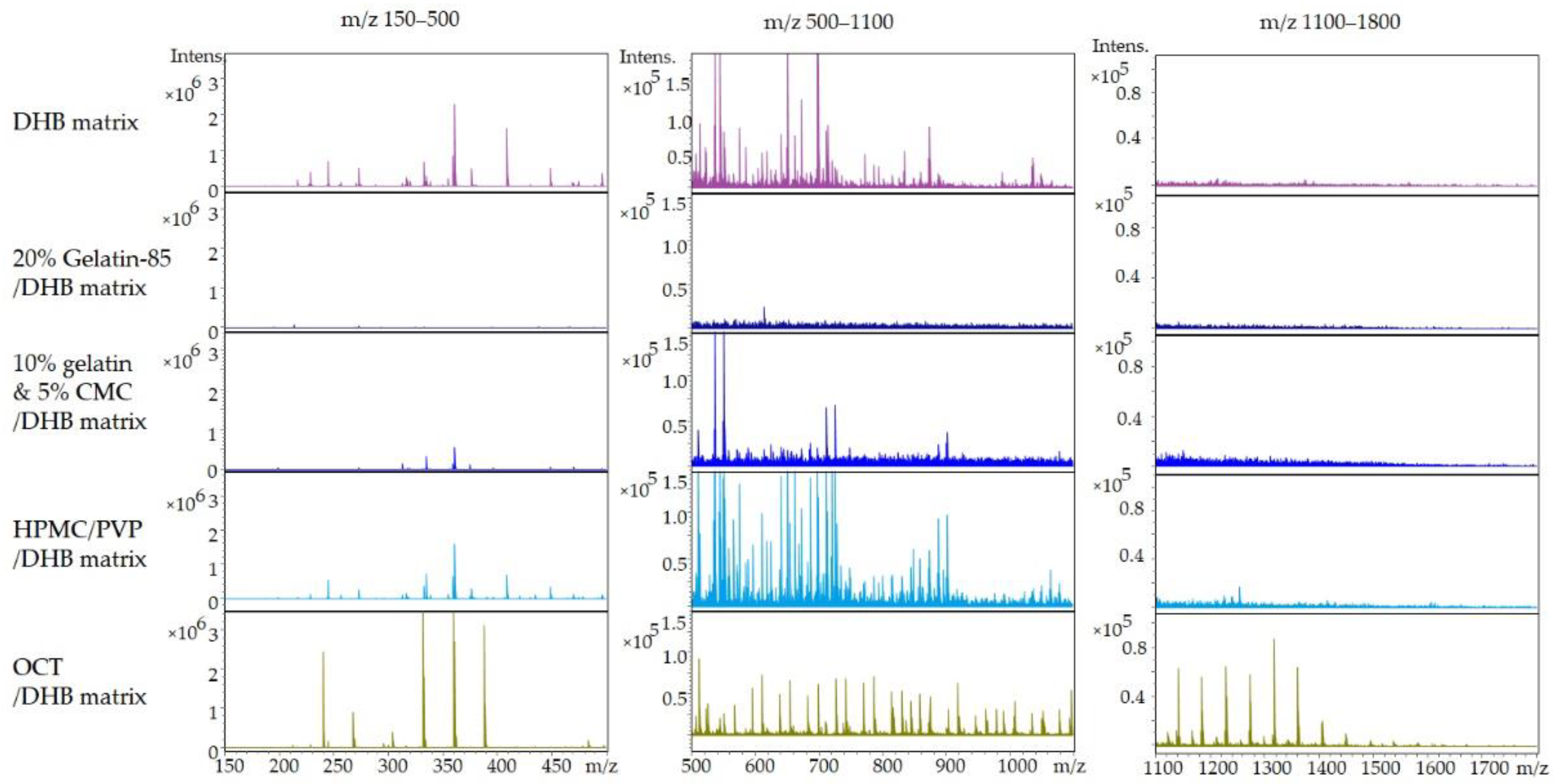
- Embedding medium has little interference and ion suppression towards detection.
- Embedding medium brings minimal interference for the interpretation of the stained sections.
- Sections will need to be preserved well and have no defects and freezing artifacts.
2.1.1. Embedding Medium Selection Based on Minimal Mass Spectrum Background
2.1.2. Embedding Medium Selection Based on the Embedding Properties of the Medium
2.2. Embedding Process with the Zebrafish-Shaped Mold
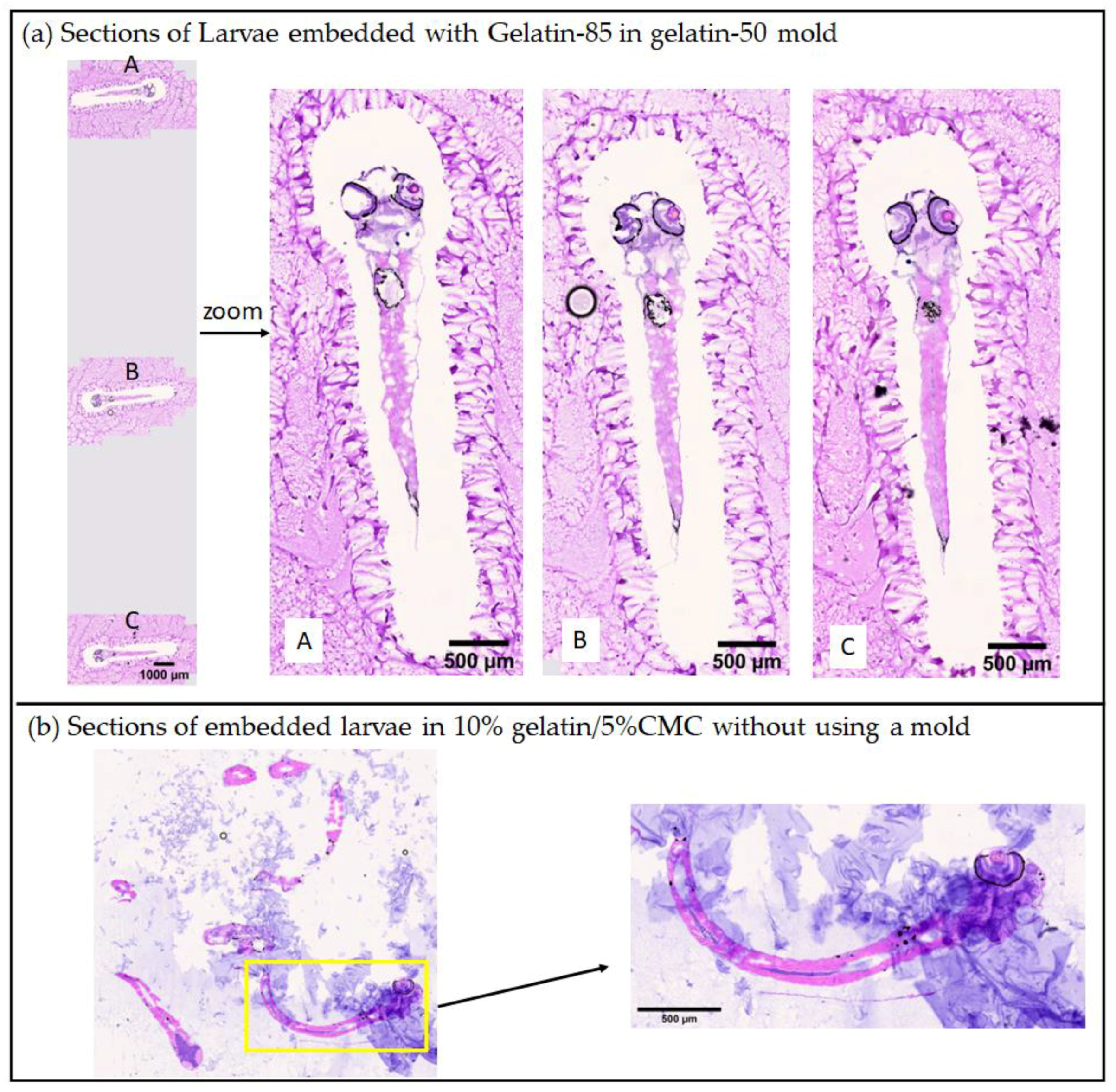
2.2.1. The 3D-Printed Zebrafish Larvae Mold for Controlled Embedding and Sectioning
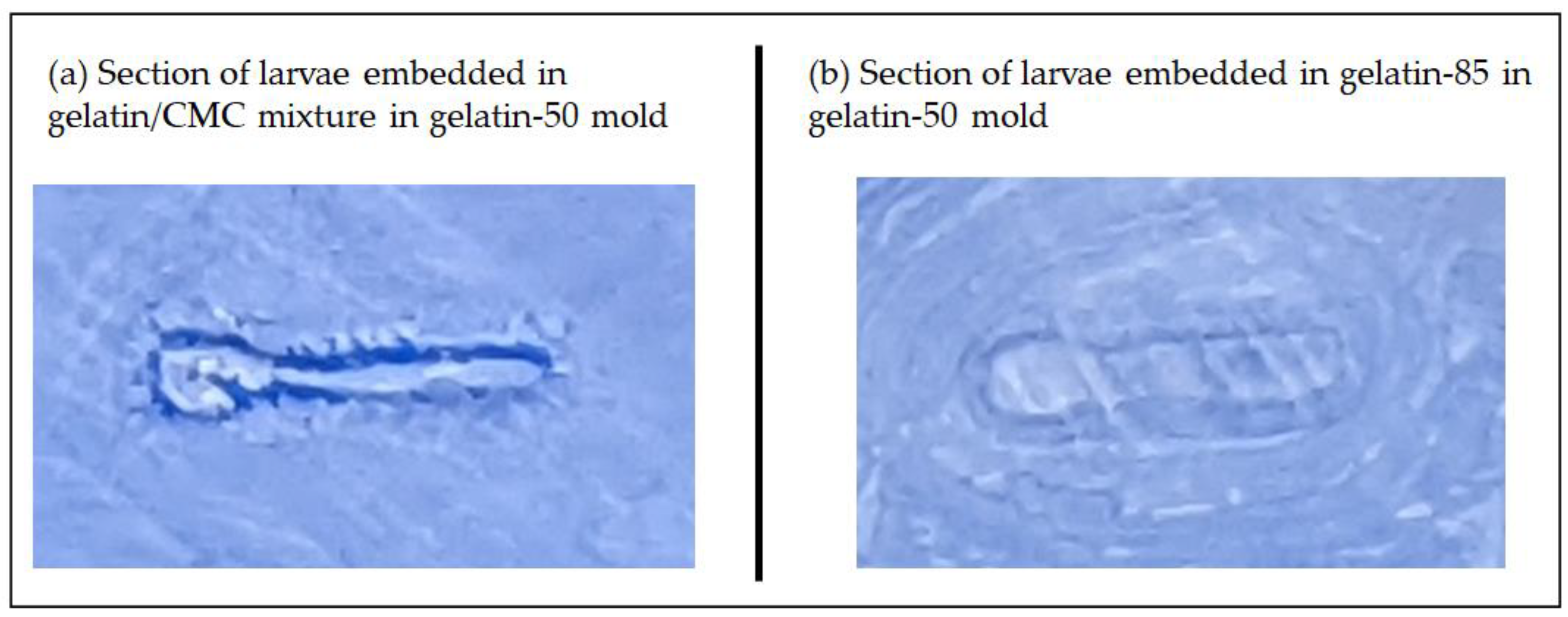
2.2.2. Advantages of Using Two Gelatin Solutions Prepared under 50 and 85 °C for Embedding
2.2.3. The PTFE Surface for Section Placement to Reduce Tissue Folding
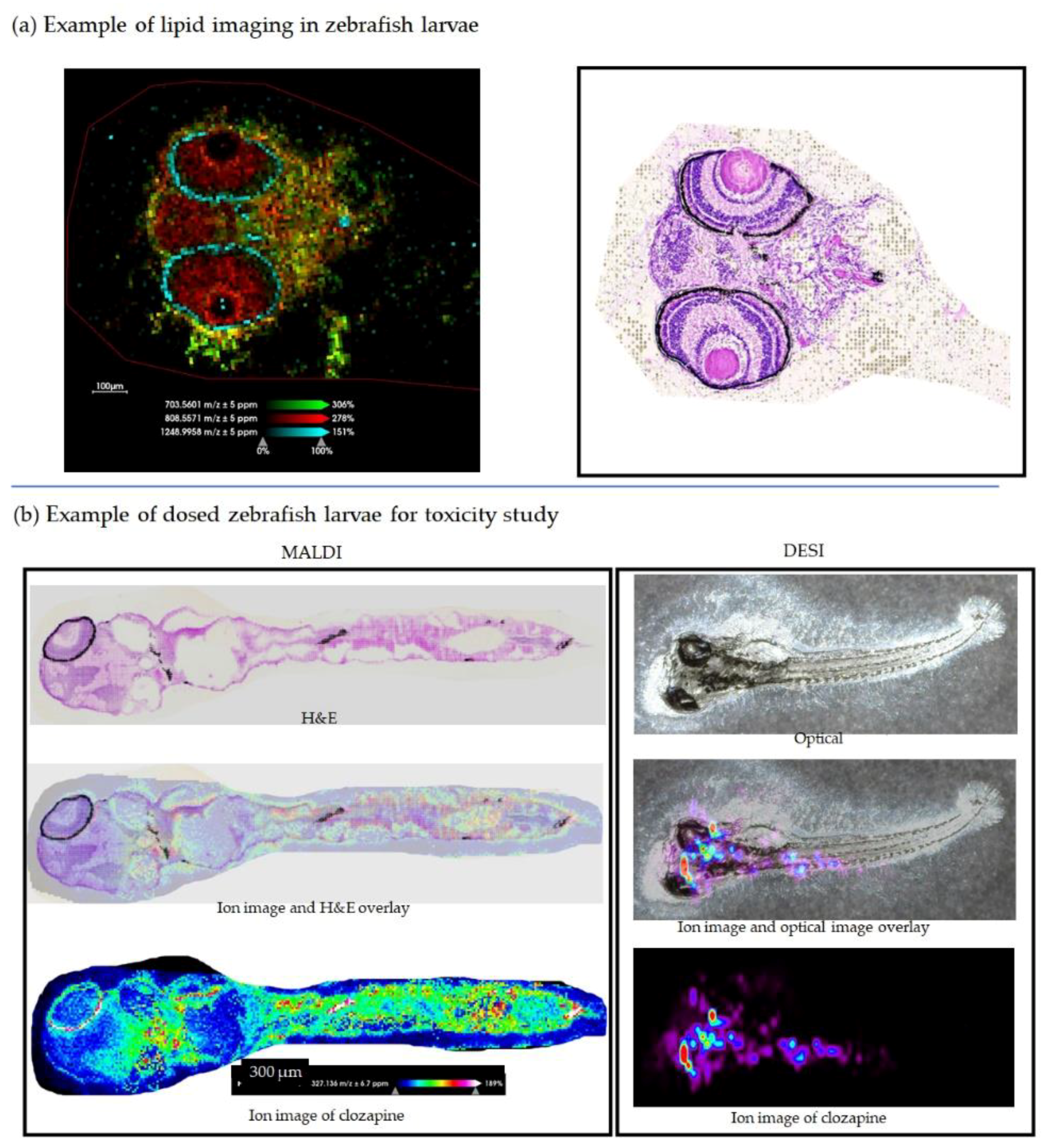
2.3. Mass Spectrometry Imaging of Zebrafish Larvae
3. Materials and Methods
3.1. Materials
3.2. Sample Preparation
3.3. Generating Molds
3.4. Animal Husbandry
3.5. Embedding Larval Zebrafish into Molds
3.6. Zebrafish Larvae Sectioning Assisted with PTFE Surface Placement
3.7. Matrix Coating
3.8. Mass Spectrometry Imaging
3.9. H&E Staining
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oberg, M.; Tornqvist, E.; Annas, A.; Granath, B.; Jalkesten, E.; Cotgreave, I. Strategic focus on 3R principles reveals major reductions in the use of animals in pharmaceutical toxicity testing. Toxicol. Lett. 2014, 229, S46. [Google Scholar] [CrossRef] [Green Version]
- Strahle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments-A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef]
- Morikane, D.; Zang, L.; Nishimura, N. Evaluation of the Percutaneous Absorption of Drug Molecules in Zebrafish. Molecules 2020, 25, 3974. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.V.; Candiracci, M. Zebrafish as screening model for detecting toxicity and drugs efficacy. J. Unexplored Med. Data 2018, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caprioli, R.M.; Farmer, T.B.; Gile, J. Molecular imaging of biological samples: Localization of peptides and proteins using MALDI-TOF MS. Anal. Chem. 1997, 69, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Sullivan-Brown, J.; Bisher, M.E.; Burdine, R.D. Embedding, serial sectioning and staining of zebrafish embryos using JB-4 resin. Nat. Protoc. 2011, 6, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.A.; Daniels, G.J.; Fournie, J.W.; Hemmer, M.J. Optimization of whole-body zebrafish sectioning methods for mass spectrometry imaging. J. Biomol. Tech. 2013, 24, 119–127. [Google Scholar] [CrossRef]
- Truchado-Garcia, M.; Harland, R.M.; Abrams, M.J. 3D-printable tools for developmental biology: Improving embryo injection and screening techniques through 3D-printing technology. bioRxiv 2018, 376657. [Google Scholar]
- Asslan, M.; Lauzon, N.; Beus, M.; Maysinger, D.; Rousseau, S. Mass spectrometry imaging in zebrafish larvae for assessing drug safety and metabolism. Anal. Bioanal. Chem. 2021, 413, 5135–5146. [Google Scholar] [CrossRef]
- Truong, J.X.M.; Spotbeen, X.; White, J.; Swinnen, J.V.; Butler, L.M.; Snel, M.F.; Trim, P.J. Removal of optimal cutting temperature (OCT) compound from embedded tissue for MALDI imaging of lipids. Anal. Bioanal. Chem. 2021, 413, 2695–2708. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.A.; Reyzer, M.L.; Caprioli, R.M. Direct tissue analysis using matrix-assisted laser desorption/ionization mass spectrometry: Practical aspects of sample preparation. J. Mass Spectrom. 2003, 38, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Dannhorn, A.; Kazanc, E.; Ling, S.; Nikula, C.; Karali, E.; Serra, M.P.; Vorng, J.L.; Inglese, P.; Maglennon, G.; Hamm, G.; et al. Universal Sample Preparation Unlocking Multimodal Molecular Tissue Imaging. Anal. Chem. 2020, 92, 11080–11088. [Google Scholar] [CrossRef] [PubMed]
- Copper, J.E.; Budgeon, L.R.; Foutz, C.A.; van Rossum, D.B.; Vanselow, D.J.; Hubley, M.J.; Clark, D.P.; Mandrell, D.T.; Cheng, K.C. Comparative analysis of fixation and embedding techniques for optimized histological preparation of zebrafish. Comp. Biochem. Phys. C 2018, 208, 38–46. [Google Scholar] [CrossRef]
- Sha, X.M.; Hu, Z.Z.; Ye, Y.H.; Xu, H.; Tu, Z.C. Effect of extraction temperature on the gelling properties and identification of porcine gelatin. Food Hydrocolloid 2019, 92, 163–172. [Google Scholar] [CrossRef]
- Baydin, T.; Aarstad, O.A.; Dille, M.J.; Hattrem, M.N.; Draget, K.I. Long-term storage stability of type A and type B gelatin gels: The effect of Bloom strength and co-solutes. Food Hydrocolloid 2022, 127, 107535. [Google Scholar] [CrossRef]
- Duenas, M.E.; Carlucci, L.; Lee, Y.J. Matrix Recrystallization for MALDI-MS Imaging of Maize Lipids at High-Spatial Resolution. J. Am. Soc. Mass. Spectr. 2016, 27, 1575–1578. [Google Scholar] [CrossRef]
- Yang, J.H.; Caprioli, R.M. Matrix Sublimation/Recrystallization for Imaging Proteins by Mass Spectrometry at High Spatial Resolution. Anal. Chem. 2011, 83, 5728–5734. [Google Scholar] [CrossRef] [Green Version]
- Bouschen, W.; Schulz, O.; Eikel, D.; Spengler, B. Matrix vapor deposition/recrystallization and dedicated spray preparation for high-resolution scanning microprobe matrix-assisted laser desorption/ionization imaging mass spectrometry (SMALDI-MS) of tissue and single cells. Rapid Commun. Mass. Spectrometry 2010, 24, 355–364. [Google Scholar] [CrossRef]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Crilly, J. The history of clozapine and its emergence in the US market: A review and analysis. Hist Psychiatry 2007, 18, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Gundlach, M.; Di Paolo, C.; Chen, Q.Q.; Majewski, K.; Haigis, A.C.; Werner, I.; Hollert, H. Clozapine modulation of zebrafish swimming behavior and gene expression as a case study to investigate effects of atypical drugs on aquatic organisms. Sci. Total. Environ. 2022, 815, 152621. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, A.R.; DeLaney, K.; Johnson, J.; Li, L.J. Mass Spectrometry Imaging: A Review of Emerging Advancements and Future Insights. Anal. Chem. 2018, 90, 240–265. [Google Scholar] [CrossRef]
- Angel, P.M.; Spraggins, J.M.; Baldwin, H.S.; Caprioli, R. Enhanced Sensitivity for High Spatial Resolution Lipid Analysis by Negative Ion Mode Matrix Assisted Laser Desorption Ionization Imaging Mass Spectrometry. Anal. Chem. 2012, 84, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Quiason, C.M.; Shahidi-Latham, S.K. Imaging MALDI MS of Dosed Brain Tissues Utilizing an Alternative Analyte Pre-extraction Approach. J. Am. Soc. Mass Spectr 2015, 26, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Huizing, L.R.S.; Ellis, S.R.; Beulen, B.W.A.M.M.; Barre, F.P.Y.; Kwant, P.B.; Vreeken, R.J.; Heeren, R.M.A. Development and evaluation of matrix application techniques for high throughput mass spectrometry imaging of tissues in the clinic. Clin. Mass. Spectrom. 2019, 12, 7–15. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Worlein, J.M.; Baker, K.; Bloomsmith, M.; Coleman, K.; Koban, T.L. The Eighth Edition of the Guide for the Care and Use of Laboratory Animals (2011); Implications for Behavioral Management. Am. J. Primatol. 2011, 73, 98. [Google Scholar]
- Prophet, E.B.; Mills, B.; Arrington, J.B.; Sobbin, L.H. Laboratory Methods in Histotechnology; American Registry of Pathology: Washington, DC, USA, 1992. [Google Scholar]
- Bankhead, P.; Loughrey, M.B.; Fernandez, J.A.; Dombrowski, Y.; Mcart, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Rendino, L.; Cassar, S.; Buck, W.; Sawicki, J.; Talaty, N.; Wagner, D. Optimization of Zebrafish Larvae Sectioning for Mass Spectrometry Imaging. Pharmaceuticals 2022, 15, 1230. https://doi.org/10.3390/ph15101230
Yang J, Rendino L, Cassar S, Buck W, Sawicki J, Talaty N, Wagner D. Optimization of Zebrafish Larvae Sectioning for Mass Spectrometry Imaging. Pharmaceuticals. 2022; 15(10):1230. https://doi.org/10.3390/ph15101230
Chicago/Turabian StyleYang, Junhai, Lauren Rendino, Steven Cassar, Wayne Buck, James Sawicki, Nari Talaty, and David Wagner. 2022. "Optimization of Zebrafish Larvae Sectioning for Mass Spectrometry Imaging" Pharmaceuticals 15, no. 10: 1230. https://doi.org/10.3390/ph15101230
APA StyleYang, J., Rendino, L., Cassar, S., Buck, W., Sawicki, J., Talaty, N., & Wagner, D. (2022). Optimization of Zebrafish Larvae Sectioning for Mass Spectrometry Imaging. Pharmaceuticals, 15(10), 1230. https://doi.org/10.3390/ph15101230