
Aromatase-Inhibitor-Induced Musculoskeletal Inflammation Is Observed Independent of Oophorectomy in a Novel Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
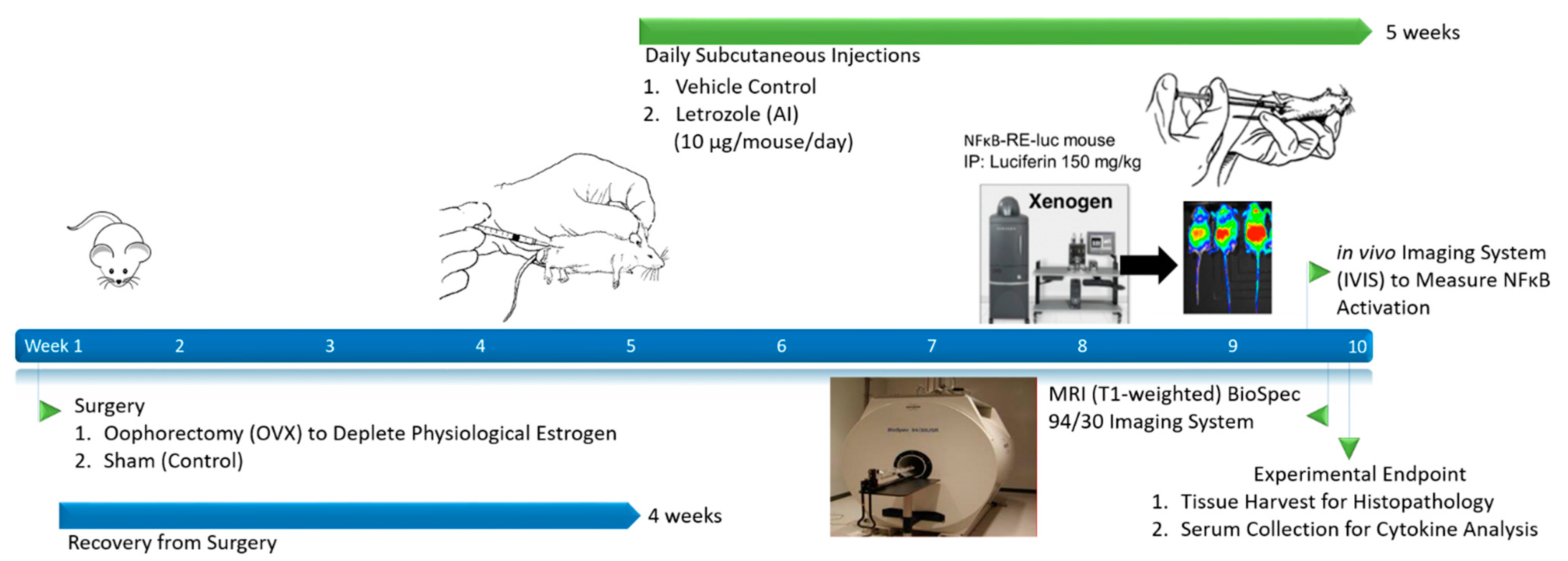
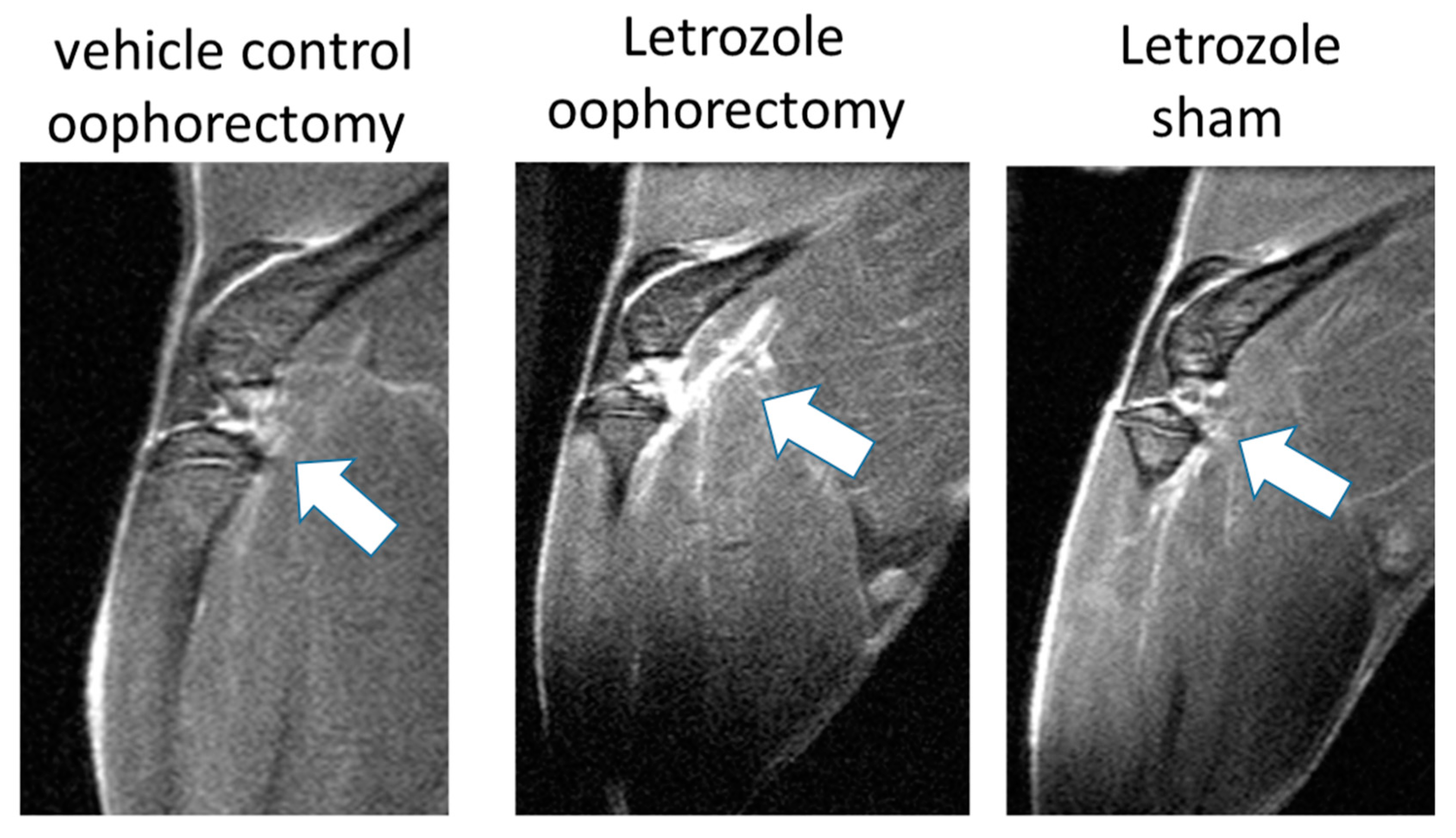
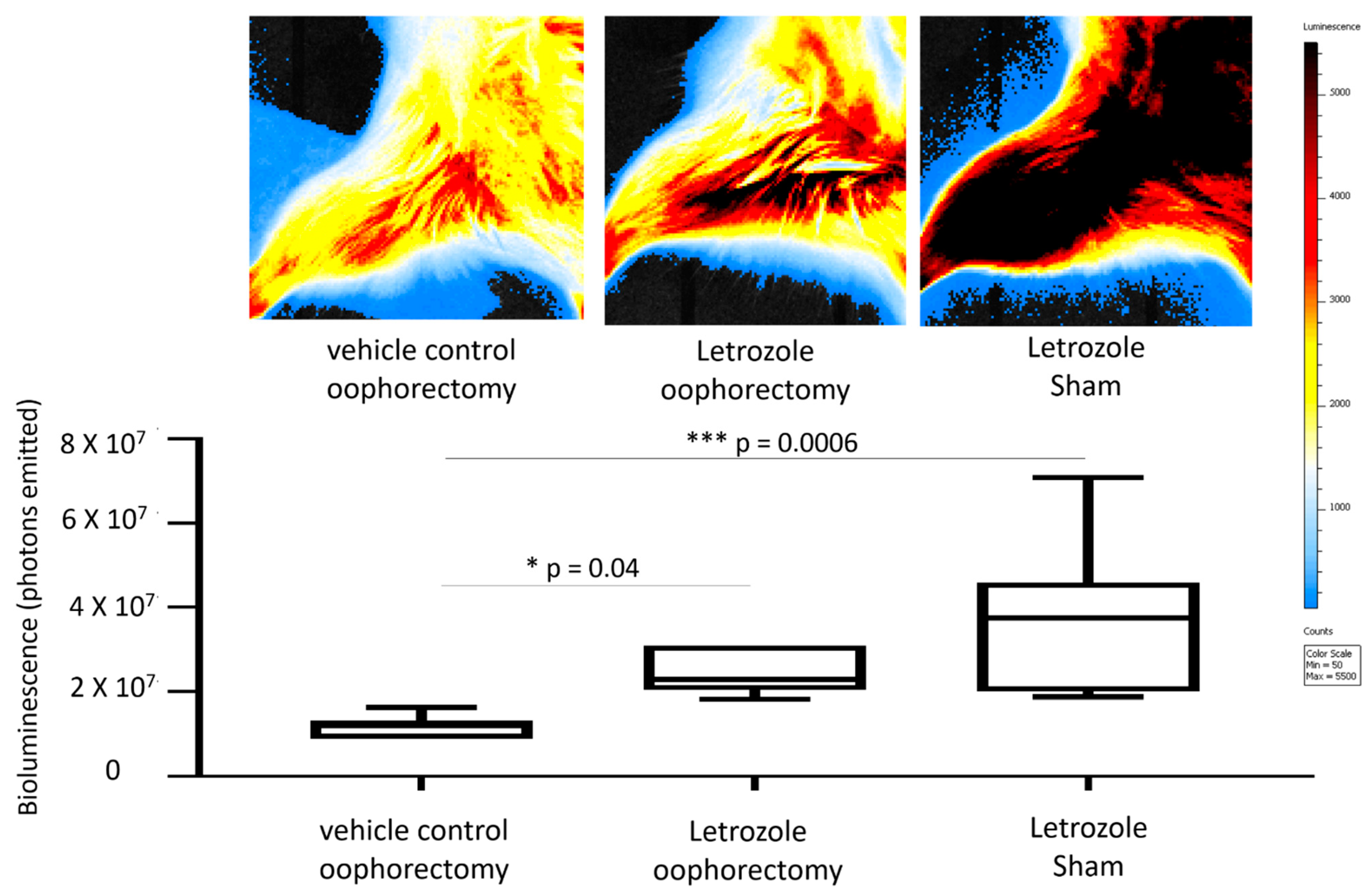
2.1. Letrozole-Induced Inflammation Detection via MRI and IVIS in Murine Hind Limbs
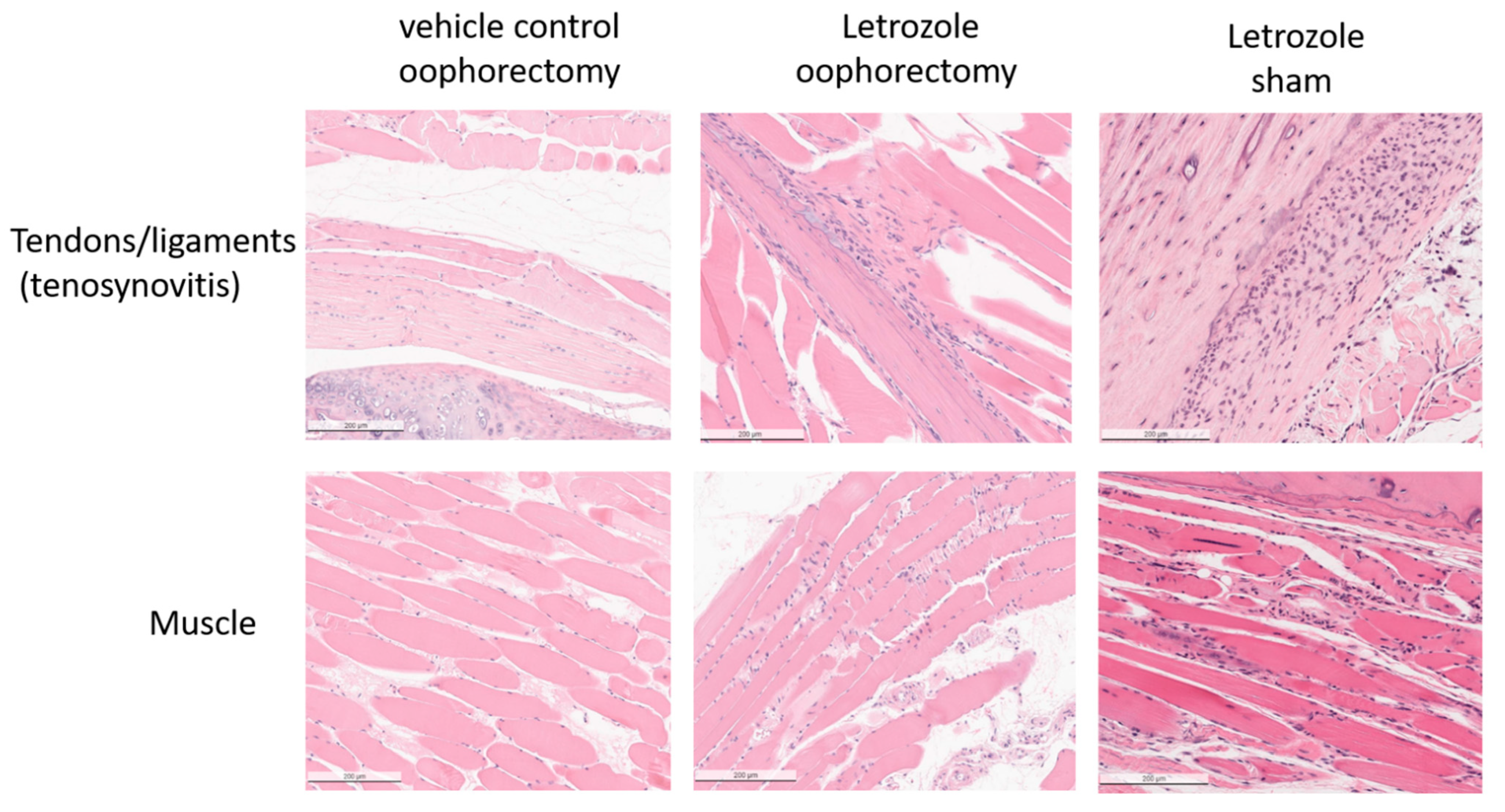
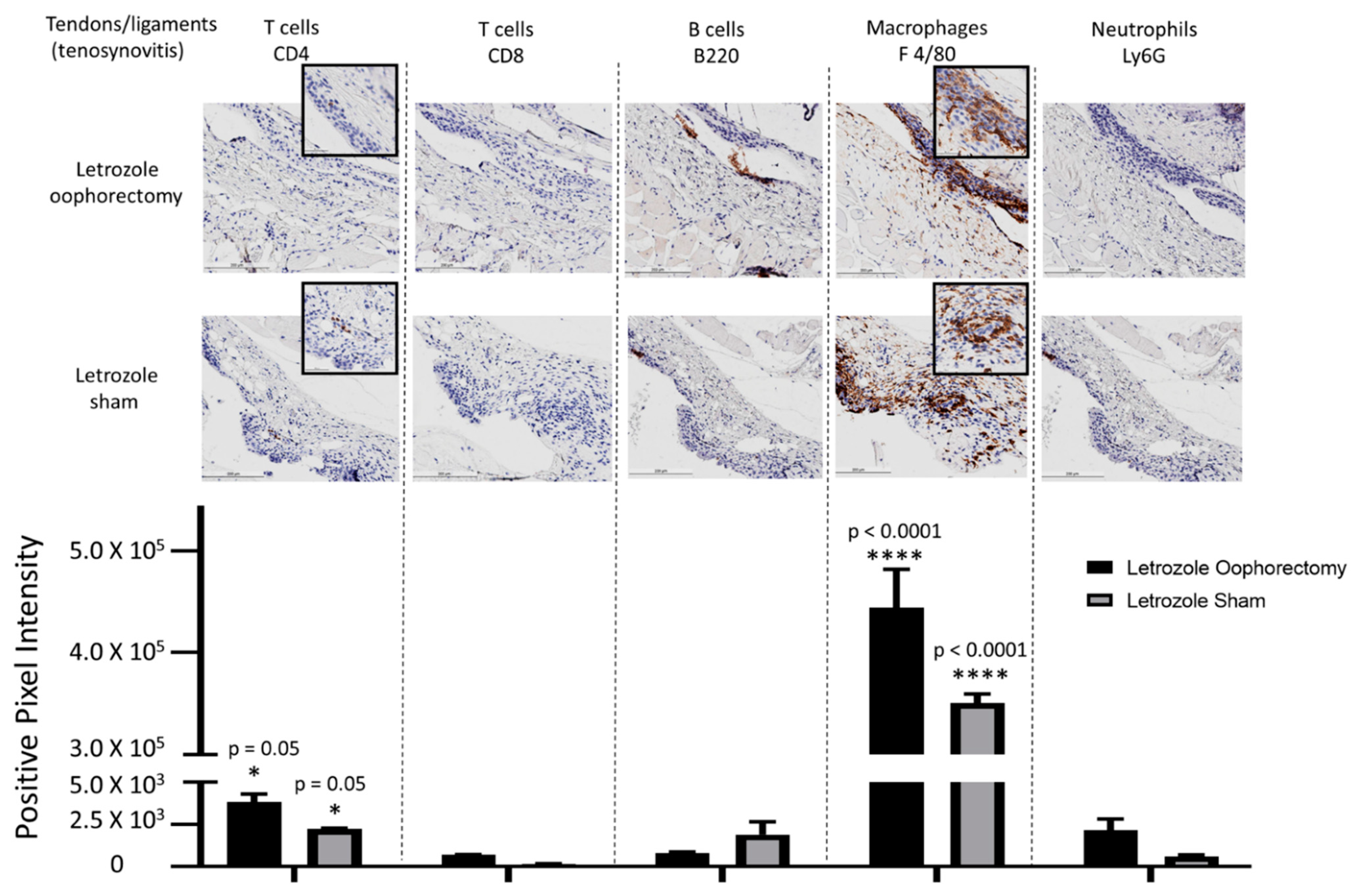
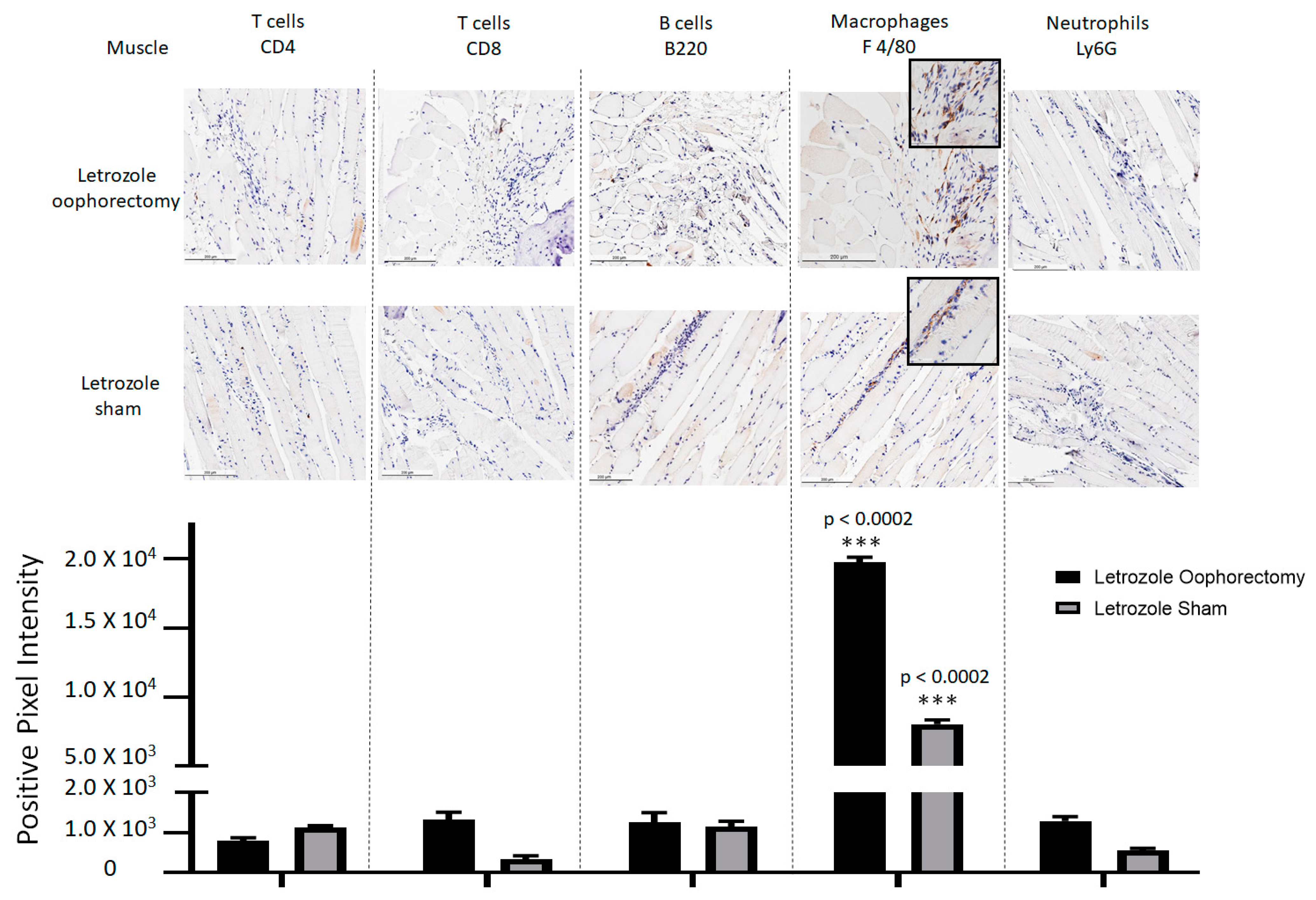
2.2. Histopathological Analysis Shows That Letrozole Increases Macrophage Infiltration
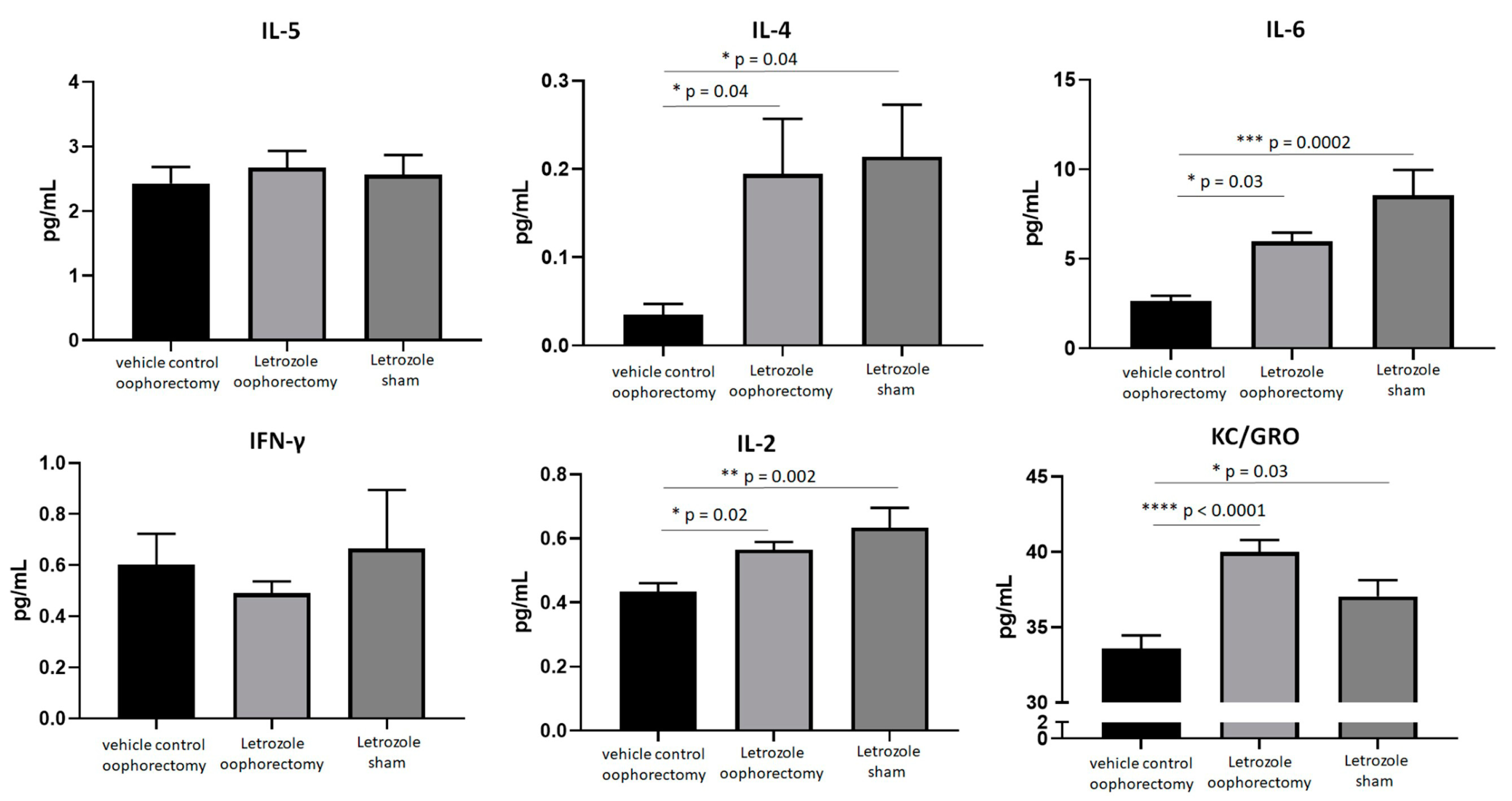
2.3. AIIA Mouse Model Shows Induction of Pro-Inflammatory Cytokines
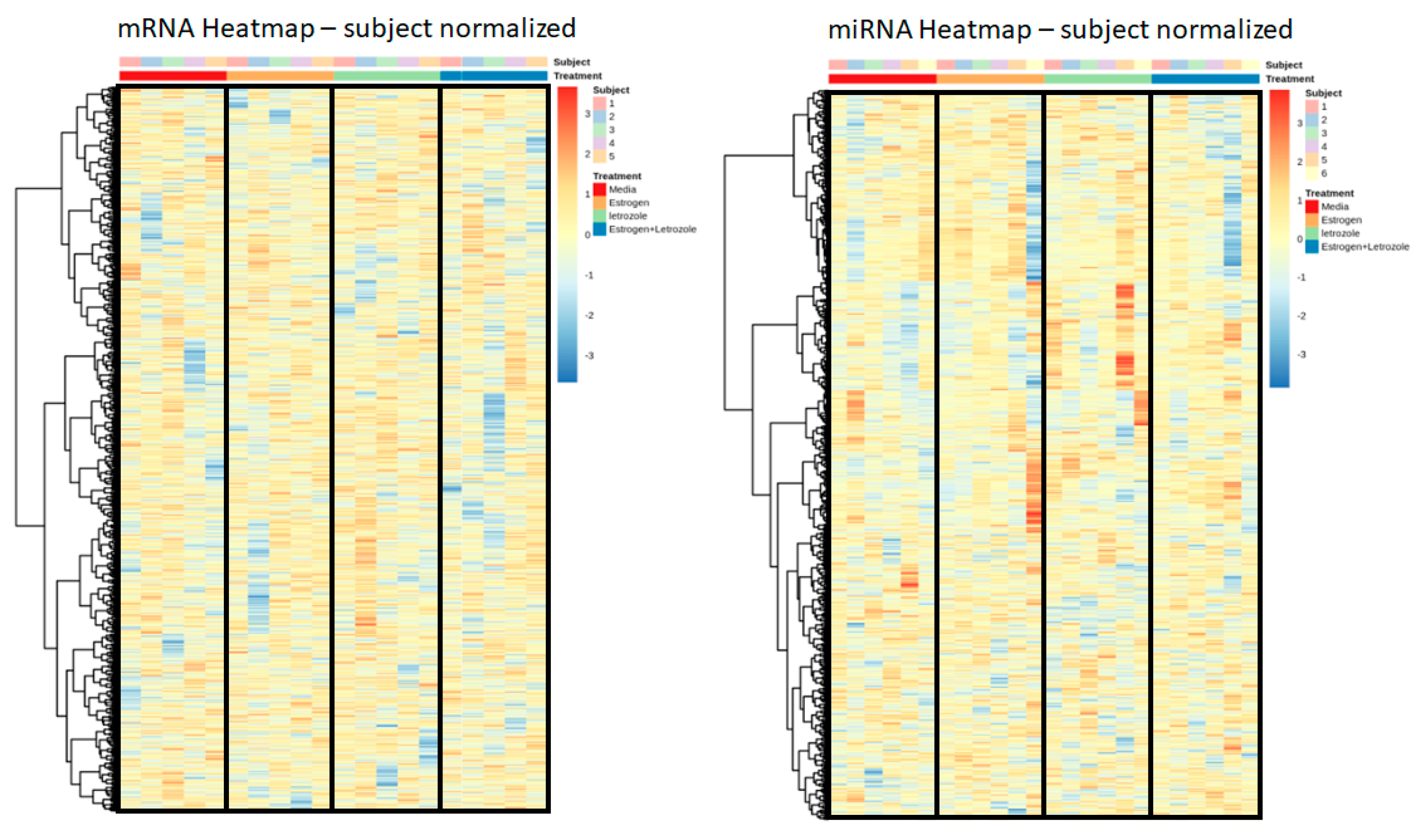
2.4. Letrozole Treatment of Human PBMCs In Vitro Does Not Produce an Observable Genomic Effect
3. Discussion
4. Materials and Methods
4.1. AIIA Induction and Sample Size Determination
4.2. Tissue Collection
4.3. Histopathology
4.4. Cytokine Measurement
4.5. Bioluminescent Imaging with IVIS
4.6. MRI
4.7. Letrozole Stimulation of Human PBMCs
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burstein, H.J.; Prestrud, A.A.; Seidenfeld, J.; Anderson, H.; Buchholz, T.A.; Davidson, N.E.; Gelmon, K.E.; Giordano, S.H.; Hudis, C.A.; Malin, J.; et al. American Society of Clinical O. American Society of Clinical Oncology clinical practice guideline: Update on adjuvant endocrine therapy for women with hormone receptor-positive breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 3784–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crew, K.D.; Greenlee, H.; Capodice, J.; Raptis, G.; Brafman, L.; Fuentes, D.; Sierra, A.; Hershman, D.L. Prevalence of joint symptoms in postmenopausal women taking aromatase inhibitors for early-stage breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2007, 25, 3877–3883. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.L.; Giles, J.T.; Ang, D.; Mohan, M.; Dadabhoy, D.; Robarge, J.; Hayden, J.; Lemler, S.; Shahverdi, K.; Powers, P.; et al. Prospective characterization of musculoskeletal symptoms in early stage breast cancer patients treated with aromatase inhibitors. Breast Cancer Res. Treat. 2008, 111, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presant, C.A.; Bosserman, L.; Young, T.; Vakil, M.; Horns, R.; Upadhyaya, G.; Ebrahimi, B.; Yeon, C.; Howard, F. Aromatase inhibitor-associated arthralgia and/or bone pain: Frequency and characterization in non-clinical trial patients. Clin. Breast Cancer 2007, 7, 775–778. [Google Scholar] [CrossRef]
- Simao, M.L.; Chahud, F.; Bicas, H.E. Effects of intraorbital injection of 6.0% carboxymethylcellulose in rabbits: Histological and ocular mechanics evaluation. Arq. Bras. Oftalmol. 2009, 72, 799–804. [Google Scholar]
- Fallowfield, L.J.; Bliss, J.; Porter, L.S.; Price, M.H.; Snowdon, C.F.; Jones, S.E.; Coombes, R.C.; Hall, E. Quality of life in the intergroup exemestane study: A randomized trial of exemestane versus continued tamoxifen after 2 to 3 years of tamoxifen in postmenopausal women with primary breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 910–917. [Google Scholar] [CrossRef]
- Din, O.S.; Dodwell, D.; Wakefield, R.J.; Coleman, R.E. Aromatase inhibitor-induced arthralgia in early breast cancer: What do we know and how can we find out more? Breast Cancer Res. Treat. 2010, 120, 525–538. [Google Scholar] [CrossRef]
- Dowsett, M.; Lonning, P.E. Anastrozole—A new generation in aromatase inhibition: Clinical pharmacology. Oncology 1997, 54 (Suppl. S2), 11–14. [Google Scholar] [CrossRef]
- Nielsen, R.H.; Christiansen, C.; Stolina, M.; Karsdal, M.A. Oestrogen exhibits type II collagen protective effects and attenuates collagen-induced arthritis in rats. Clin. Exp. Immunol. 2008, 152, 21–27. [Google Scholar] [CrossRef]
- Pfeilschifter, J.; Köditz, R.; Pfohl, M.; Schatz, H. Changes in proinflammatory cytokine activity after menopause. Endocr. Rev. 2002, 23, 90–119. [Google Scholar] [CrossRef]
- Dawson-Basoa, M.; Gintzler, A.R. Involvement of spinal cord delta opiate receptors in the antinociception of gestation and its hormonal simulation. Brain Res. 1997, 757, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Young, N.A.; Wu, L.-C.; Burd, C.; Friedman, A.K.; Kaffenberger, B.H.; Rajaram, M.; Schlesinger, L.S.; James, H.; Shupnik, M.A.; Jarjour, W.N. Estrogen modulation of endosome-associated toll-like receptor 8: An IFNalpha-independent mechanism of sex-bias in systemic lupus erythematosus. Clin. Immunol. 2014, 151, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burstein, H.J.; Lacchetti, C.; Anderson, H.; Buchholz, T.; Davidson, N.E.; Gelmon, K.E.; Giordano, S.H.; Hudis, C.A.; Solky, A.J.; Stearns, V.; et al. Adjuvant Endocrine Therapy for Women With Hormone Receptor-Positive Breast Cancer: American Society of Clinical Oncology Clinical Practice Guideline Update on Ovarian Suppression. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 1689–1701. [Google Scholar] [CrossRef] [PubMed]
- Burstein, H.J.; Griggs, J.J.; Prestrud, A.A.; Temin, S. American society of clinical oncology clinical practice guideline update on adjuvant endocrine therapy for women with hormone receptor-positive breast cancer. J. Oncol. Pract. 2010, 6, 243–246. [Google Scholar] [CrossRef]
- Young, N.A.; Bruss, M.S.; Gardner, M.; Willis, W.L.; Mo, X.; Valiente, G.R.; Cao, Y.; Liu, Z.; Jarjour, W.N.; Wu, L.-C. Oral Administration of Nano-Emulsion Curcumin in Mice Suppresses Inflammatory-Induced NFkappaB Signaling and Macrophage Migration. PLoS ONE 2014, 9, e111559. [Google Scholar] [CrossRef]
- Davignon, J.-L.; Hayder, M.; Baron, M.; Boyer, J.-F.; Constantin, A.; Apparailly, F.; Poupot, R.; Cantagrel, A. Targeting monocytes/macrophages in the treatment of rheumatoid arthritis. Rheumatology 2013, 52, 590–598. [Google Scholar] [CrossRef] [Green Version]
- Cutolo, M.; Campitiello, R.; Gotelli, E.; Soldano, S. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis. Front. Immunol. 2022, 13, 867260. [Google Scholar] [CrossRef]
- Liao, W.; Lin, J.-X.; Leonard, W.J. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 2013, 38, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Choi, P.; Reiser, H. IL-4: Role in disease and regulation of production. Clin. Exp. Immunol. 1998, 113, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, H.; Moskaug, J.O.; Fromm, S.H.; Blomhoff, R. In vivo imaging of NF-kappa B activity. J. Immunol. 2002, 168, 1441–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, D.J.; Jansen, E.D.; Powers, A.C.; Wang, T.G. A novel method of monitoring response to islet transplantation: Bioluminescent imaging of an NF-kB transgenic mouse model. Transplantation 2006, 81, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Sadikot, R.T.; Zeng, H.; Joo, M.; Everhart, M.B.; Sherrill, T.P.; Li, B.; Cheng, D.-S.; Yull, F.E.; Christman, J.W.; Blackwell, T.S. Targeted immunomodulation of the NF-kappaB pathway in airway epithelium impacts host defense against Pseudomonas aeruginosa. J. Immunol. 2006, 176, 4923–4930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemble, S.; Croft, A.P. Critical Role of Synovial Tissue-Resident Macrophage and Fibroblast Subsets in the Persistence of Joint Inflammation. Front. Immunol. 2021, 12, 715894. [Google Scholar] [CrossRef] [PubMed]
- Chow, Y.Y.; Chin, K.-Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [Green Version]
- Pfeiler, G.; Königsberg, R.; Hadji, P.; Fitzal, F.; Maroske, M.; Dressel-Ban, G.; Zellinger, J.; Exner, R.; Seifert, M.; Singer, C.; et al. Impact of body mass index on estradiol depletion by aromatase inhibitors in postmenopausal women with early breast cancer. Br. J. Cancer 2013, 109, 1522–1527. [Google Scholar] [CrossRef] [Green Version]
- Niravath, P. Aromatase inhibitor-induced arthralgia: A review. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2013, 24, 1443–1449. [Google Scholar] [CrossRef]
- Cirillo, D.; Wallace, R.B.; Wu, L.; Yood, R.A. Effect of hormone therapy on risk of hip and knee joint replacement in the Women’s Health Initiative. Arthritis Rheum. 2006, 54, 3194–3204. [Google Scholar] [CrossRef]
- Young, N.A.; Sharma, R.; Friedman, A.K.; Kaffenberger, B.H.; Bolon, B.; Jarjour, W.N. Aberrant muscle antigen exposure in mice is sufficient to cause myositis in a treg cell-deficient milieu. Arthritis Rheum. 2013, 65, 3259–3270. [Google Scholar] [CrossRef] [Green Version]
- Young, N.A.; Wu, L.-C.; Bruss, M.; Kaffenberger, B.H.; Hampton, J.; Bolon, B.; Jarjour, W.N. A chimeric human-mouse model of Sjogren’s syndrome. Clin. Immunol. 2014, 156, 1–8. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, N.A.; Hampton, J.; Sharma, J.; Jablonski, K.; DeVries, C.; Bratasz, A.; Wu, L.-C.; Lustberg, M.; Reinbolt, R.; Jarjour, W.N. Aromatase-Inhibitor-Induced Musculoskeletal Inflammation Is Observed Independent of Oophorectomy in a Novel Mouse Model. Pharmaceuticals 2022, 15, 1578. https://doi.org/10.3390/ph15121578
Young NA, Hampton J, Sharma J, Jablonski K, DeVries C, Bratasz A, Wu L-C, Lustberg M, Reinbolt R, Jarjour WN. Aromatase-Inhibitor-Induced Musculoskeletal Inflammation Is Observed Independent of Oophorectomy in a Novel Mouse Model. Pharmaceuticals. 2022; 15(12):1578. https://doi.org/10.3390/ph15121578
Chicago/Turabian StyleYoung, Nicholas A., Jeffrey Hampton, Juhi Sharma, Kyle Jablonski, Courtney DeVries, Anna Bratasz, Lai-Chu Wu, Maryam Lustberg, Raquel Reinbolt, and Wael N. Jarjour. 2022. "Aromatase-Inhibitor-Induced Musculoskeletal Inflammation Is Observed Independent of Oophorectomy in a Novel Mouse Model" Pharmaceuticals 15, no. 12: 1578. https://doi.org/10.3390/ph15121578
APA StyleYoung, N. A., Hampton, J., Sharma, J., Jablonski, K., DeVries, C., Bratasz, A., Wu, L. -C., Lustberg, M., Reinbolt, R., & Jarjour, W. N. (2022). Aromatase-Inhibitor-Induced Musculoskeletal Inflammation Is Observed Independent of Oophorectomy in a Novel Mouse Model. Pharmaceuticals, 15(12), 1578. https://doi.org/10.3390/ph15121578