
Neuropharmacological Activities of Ceiba aesculifolia (Kunth) Britten & Baker f (Malvaceae)

 , , ,
, , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of C. aesculifolia Extract
2.2. Acute Toxicity
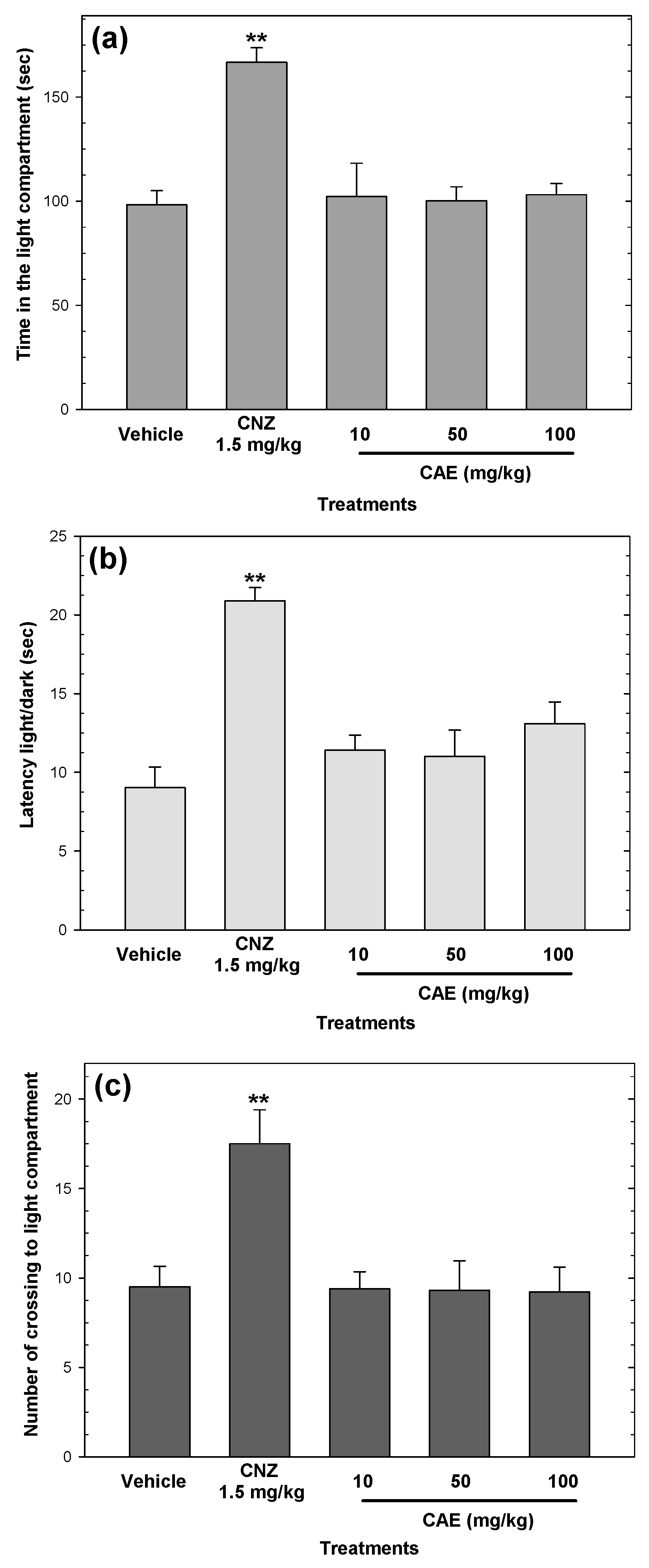
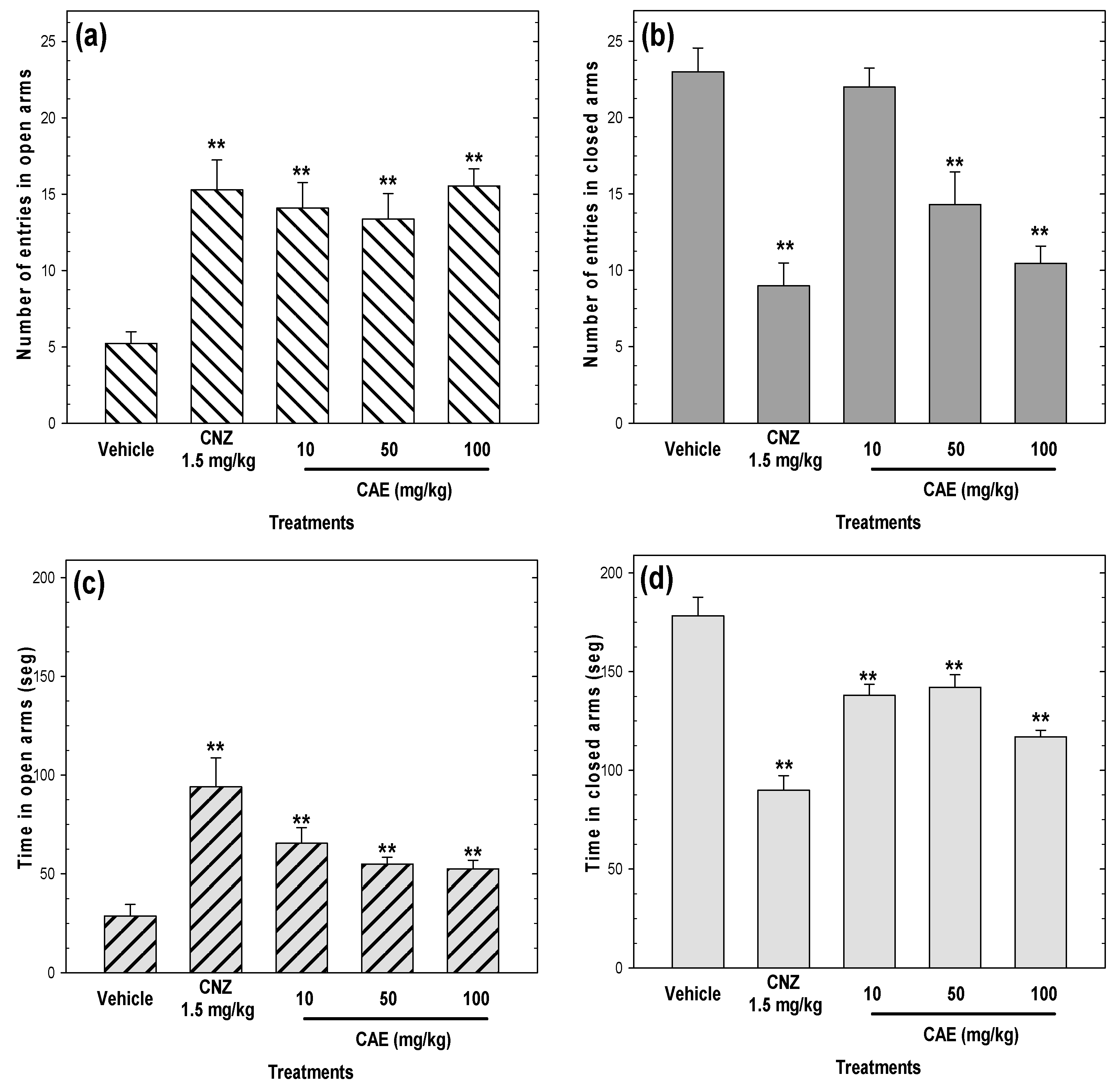
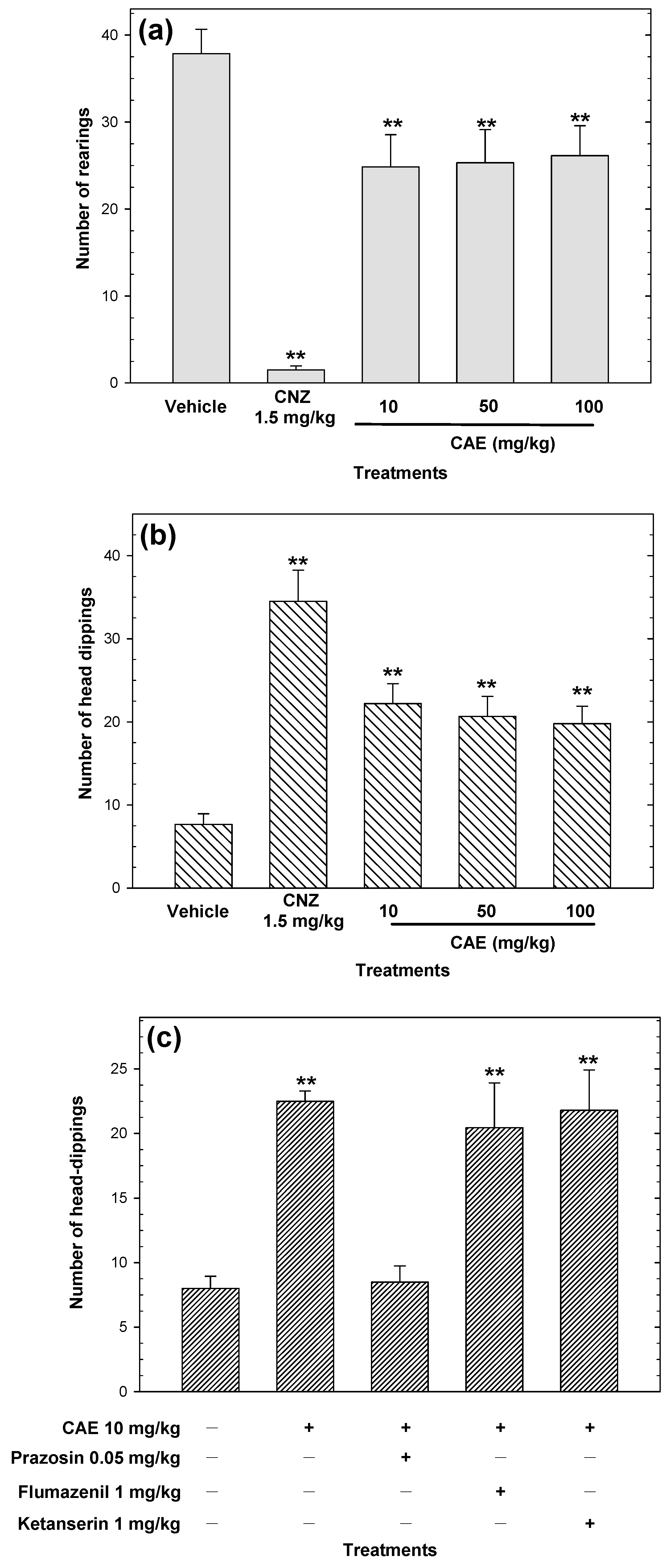
2.3. Anxiolytic-Like Activity
2.4. Antidepressant-like Activity
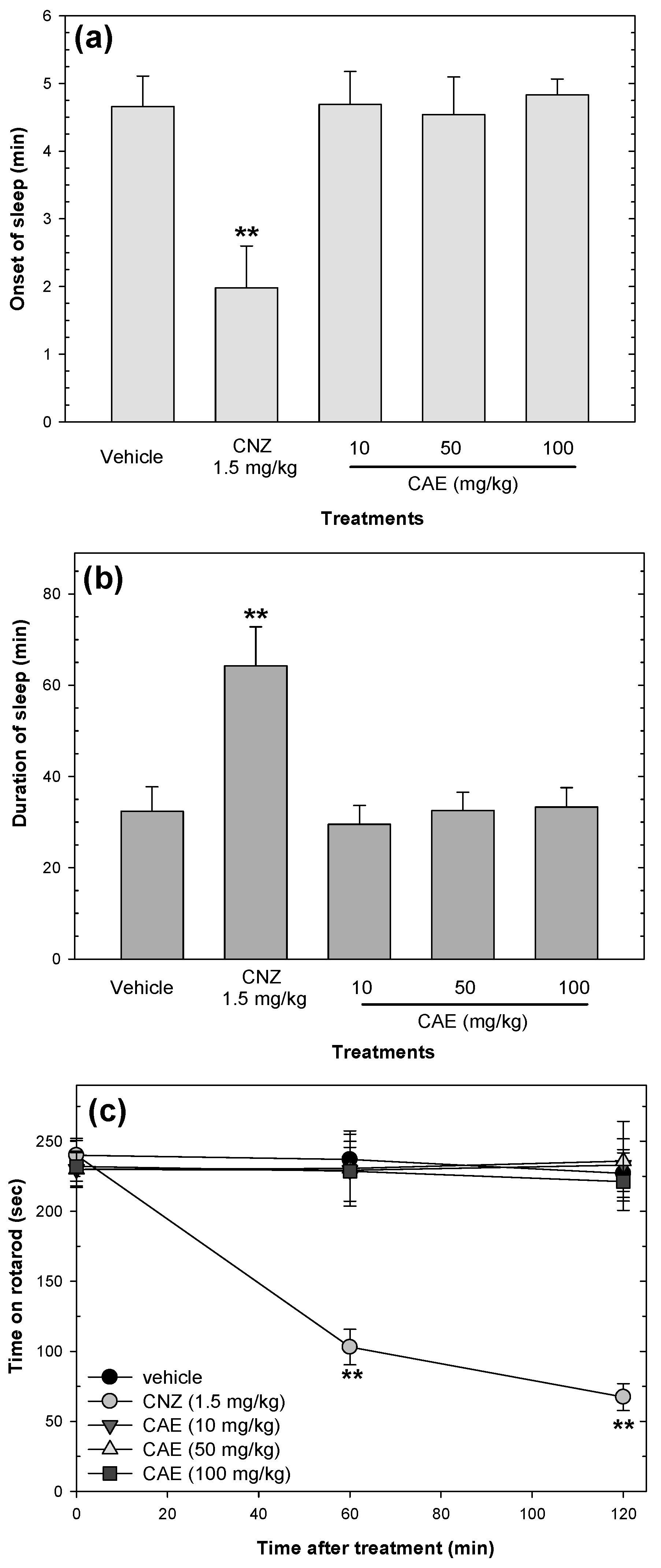
2.5. Effects on Sedation and Motor Coordination
2.6. Effects on Spatial Working Memory
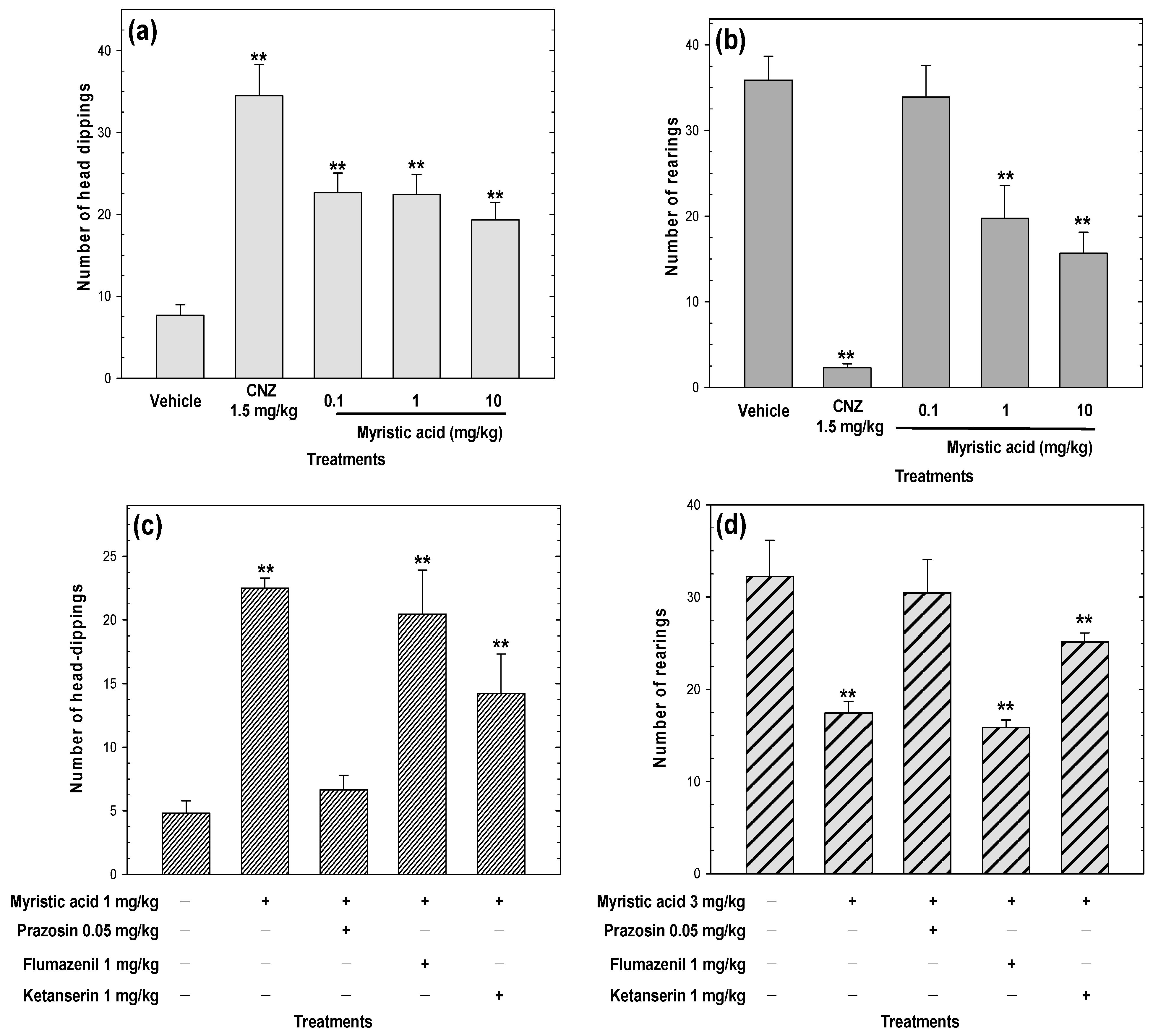
2.7. Myristic Acid Induces Anxiolytic-Like Effects
2.8. Myristic Acid Induces Sedative Effects
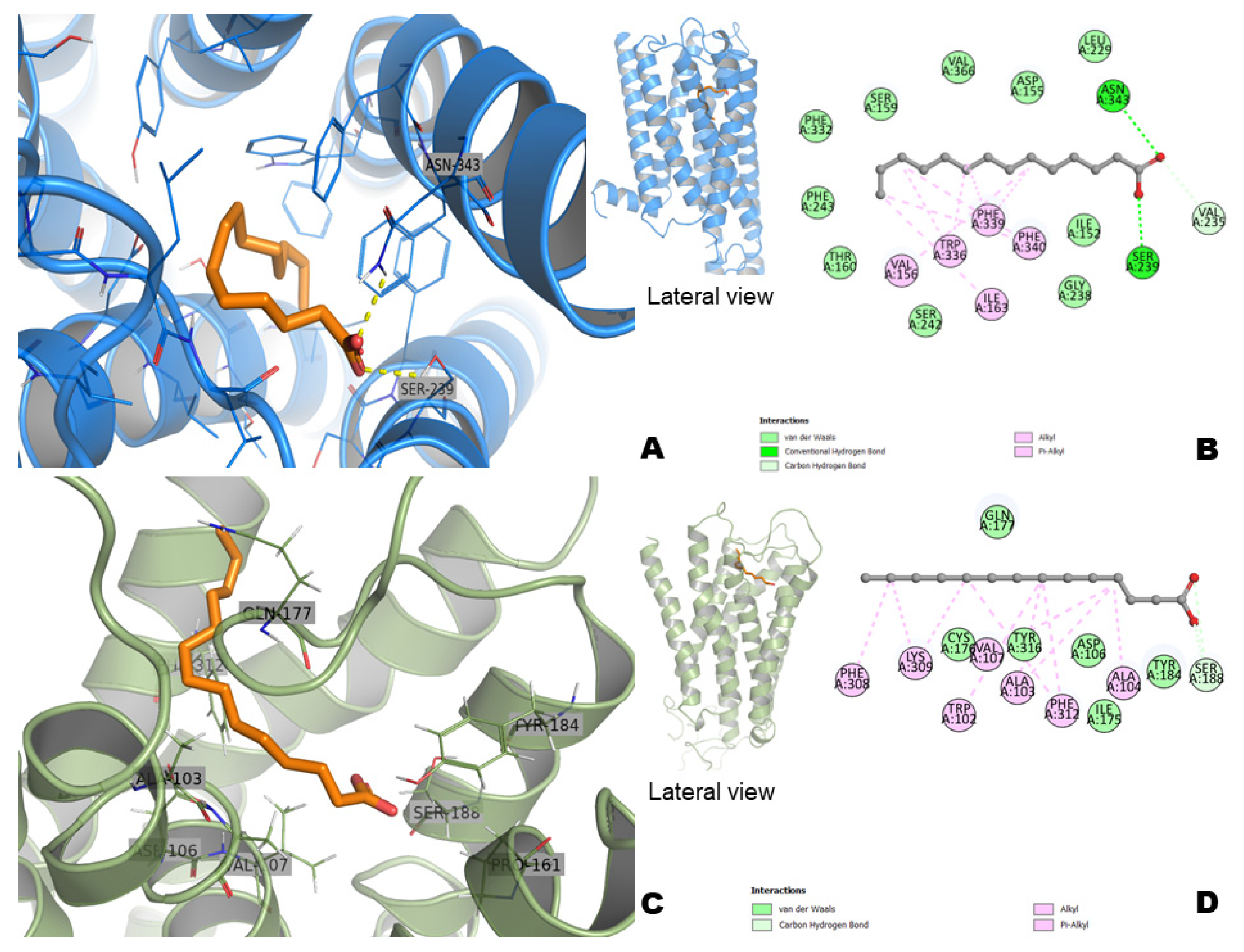
2.9. Docking Study with Myristic Acid
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Plant Material
4.3. Extraction
4.4. GC-MS Analysis
4.5. Animals
4.6. Pharmacological Treatment
4.7. Acute Toxicity
4.8. Light/Dark Test
4.9. Open Field Test
4.10. Elevated Plus Maze
4.11. Hole-Board Test
4.12. Exploratory Cylinder Test
4.13. Tail Suspension Test
4.14. Forced Swimming Test
4.15. Pentobarbital-Induced Sleep Test
4.16. Rotarod Test
4.17. Y-maze Alternation Task
4.18. Homology Modeling
4.19. Molecular Docking Studies
4.20. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Efferth, T.; Zacchino, S.; Georgiev, M.I.; Liu, L.; Wagner, H.; Panossian, A. Nobel Prize for artemisinin brings phytotherapy into the spotlight. Phytomedicine 2015, 22, A1–A3. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Maqueo, X.; Gamboa-deBuen, A. The biology of the genus ceiba, a potential source for sustainable production of natural fiber. Plants 2022, 11, 521. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Gómez, C.I.; Alonso-Castro, A.J. La ceiba: Un símbolo en la mitología Maya con propiedades farmacológicas (Ceiba: A symbol in Mayan mythology with pharmacological properties). Nat. Tecnol. 2021, 2, 13–17. [Google Scholar]
- López-Coronado, G.A. Contribución al conocimiento de las plantas medicinales de los tianguis de la zona metropolitana de Guadalajara (Contribution to the knowledge of the medicinal plants of the tianguis of the metropolitan area of Guadalajara). Bachelor’s Thesis, University of Guadalajara, Guadalajara, México, 1994. [Google Scholar]
- Serralta-Peraza, L.E.; Rosado-May, F.J.; Méndez-Mena, J.I.; Cruz-Martínez, S. Flora con uso medicinal en Oxtankah, Quinatana Roo, México. In Contribuciones de la Ciencia al Manejo Costero Integrado de la Bahía de Chetumal y su Área de Influencia; Rosado-May, F.J., Romero-Mayo, R., Navarrete, A.J., Eds.; Universidad de Quintana Roo: Chetumal, México, 2002; pp. 45–56. [Google Scholar]
- Khalil, A.; Giribabu, N.; Yelumalai, S.; Shahzad, H.; Kilari, E.K.; Salleh, N. Myristic acid defends against testicular oxidative stress, inflammation, apoptosis: Restoration of spermatogenesis, steroidogenesis in diabetic rats. Life Sci. 2021, 278, 119605. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Serrano-Vega, R.; Pérez-Gutiérrez, S.; Isiordia-Espinoza, M.A.; Solorio-Alvarado, C.R. Myristic acid reduces skin inflammation and nociception. J. Food Biochem. 2022, 46, e14013. [Google Scholar] [CrossRef]
- Muñoz-Cazares, N.; Aguilar-Rodríguez, S.; García-Contreras, R.; Soto-Hernández, M.; Martínez-Vázquez, M.; Palma-Tenango, M.; Prado-Galbarro, F.J.; Castillo-Juárez, I. Phytochemical screening and anti-virulence properties of Ceiba pentandra and Ceiba aesculifolia (Malvaceae) bark extracts and fractions. Bot. Sci. 2018, 96, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aziz, A.W.A.; Elwan, N.M.; Shaaban, R.S.; Osman, N.S.; Mohamed, M.A. High-performance liquid chromatography-fingerprint analyses, in vitro cytotoxicity, antimicrobial and antioxidant activities of the extracts of Ceiba speciosa growing in Egypt. Egypt. J. Chem. 2021, 64, 1831–1843. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Orabi, M.A.A.; Abdelhamid, R.A.; Abdelkader, M.S.A.; Darwish, F.M.M. Chemical and cytotoxic investigation of non-polar extract from Ceiba pentandra (L.) Gaertn.: A study supported by computer based screening. J. Appl. Pharm. Sci. 2018, 8, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Rosselli, S.; Tundis, R.; Bruno, M.; Leporini, M.; Falco, T.; Gagliano Candela, R.; Badalamenti, N.; Loizzo, M.R. Ceiba speciosa (A. St.-Hil.) seeds oil: Fatty acids profiling by GC-MS and NMR and bioactivity. Molecules 2020, 25, 1037. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Castro, A.J.; Domínguez, F.; Ruiz-Padilla, A.J.; Campos-Xolalpa, N.; Zapata-Morales, J.R.; Carranza-Alvarez, C.; Maldonado-Miranda, J.J. Medicinal plants from North and Central America and the Caribbean considered toxic for humans: The other side of the coin. Evid. Based Complement. Alternat. Med. 2017, 2017, 9439868. [Google Scholar] [CrossRef] [Green Version]
- Kullesskaya, N.; Voikar, V. Assessment of mouse anxiety-like behavior in the light-dark box and open-field arena: Role of equipment and procedure. Physiol. Behav. 2014, 133, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.K.; Sharma, A.C. Elevated plus-maze: A novel psychobehavioral tool to measure anxiety in rodents. Methods Find. Exp. Clin. Pharmacol. 1991, 13, 573–577. [Google Scholar] [PubMed]
- Alonso-Castro, A.J.; Alba-Betancourt, C.; Yáñez-Barrientos, E.; Luna-Rocha, C.; Páramo-Castillo, A.S.; Aragón-Martínez, O.H.; Zapata-Morales, J.R.; Cruz-Jiménez, G.; Gasca-Martínez, D.; González-Ibarra, A.A.; et al. Diuretic activity and neuropharmacological effects of an ethanol extract from Senna septemtrionalis (Viv.) H.S. Irwin & Barneby (Fabaceae). J. Ethnopharmacol. 2019, 239, 111923. [Google Scholar] [CrossRef] [PubMed]
- File, S.E.; Wardill, A.G. Validity of head-dipping as a measure of exploration in a modified hole-board. Psychopharmacologia 1975, 44, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Markou, A.; Lucki, I. Assessing antidepressant activity in rodents: Recent developments and future needs. Trends Pharmacol. Sci. 2002, 23, 238–245. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Shinomiya, K.; Shigemoto, Y.; Omichi, J.; Utsu, Y.; Mio, M.; Kamei, C. Effects of three hypnotics on the sleep–wakefulness cycle in sleep-disturbed rats. Psychopharmacol. 2004, 173, 203–209. [Google Scholar] [CrossRef]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depres. Anxiety. 2000, 12 (Suppl. 1), 2–19. [Google Scholar] [CrossRef]
- Stone, E.A.; Quartermain, D.; Lin, Y.; Lehmann, M.L. Central alpha1-adrenergic system in behavioral activity and depression. Biochem. Pharmacol. 2007, 73, 1063–1075. [Google Scholar] [CrossRef]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Contreras, C.M.; Rodríguez-Landa, J.F.; García-Ríos, R.I.; Cueto-Escobedo, J.; Guillen-Ruiz, G.; Bernal-Morales, B. Myristic acid produces anxiolytic-like effects in Wistar rats in the elevated plus maze. BioMed Res. Int. 2014, 2014, 492141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisstaub, N.V.; Zhou, M.; Lira, A.; Lambe, E.; González-Maeso, J.; Hornung, J.P.; Sibille, E.; Underwood, M.; Itohara, S.; Dauer, W.T.; et al. Cortical 5-HT2A receptor signaling modulates anxiety-like behaviors in mice. Science 2006, 313, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanover, K.E.; Davis, R.E. Role of 5-HT2A receptor antagonists in the treatment of insomnia. Nat. Sci. Sleep 2010, 2, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, D.M.; Doze, V.A. Cardiac and neuroprotection regulated by α(1)-adrenergic receptor subtypes. J. Recept. Signal. Transduct. Res. 2011, 31, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Lara, B.; Enciso-Donis, I.; Wrobel, K.; Wrobel, K. Determination of six priority phthalates and di(ethylhexyl) adipate in maize tortilla by gas chromatography-tandem mass spectrometry in multiple reaction monitoring mode. J. Mex. Chem. Soc. 2018, 62, 270–281. [Google Scholar] [CrossRef] [Green Version]
- OECD. OECD Guideline for Testing of Chemical 423: Acute Oral Toxicity—Acute Toxic Class Method; Organization of Economic Co-Operation and Development: Paris, France, 2001. [Google Scholar]
- Tang, A.H.; Smith, M.W.; Carter, D.B.; Im, W.B.; VonVoigtlander, P.F. U-90042, a sedative/hypnotic compound that interacts differentially with the GABAA receptor subtypes. J. Pharmacol. Exp. Ther. 1995, 275, 761–767. [Google Scholar]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound | RT | ERI | TRI | %RSD |
|---|---|---|---|---|---|
| 1 | Trimethylsilyl 3,4-bis[(trimethylsilyl)-oxy]butanoate | 7.97 | 1422 | 1457 | 2.40 |
| 2 | 2,2,9,9-Tetramethyl-5,6-bis[(trimethylsilyl)oxy]-3,8-dioxa-2,9-disiladecane | 8.673 | 1488 | 1478 | 0.68 |
| 3 | meso-Erythritol, tetrakis(trimethylsilyl) ether | 8.758 | 1496 | 1500 | 0.27 |
| 4 | Tetronic acid, tetrakis-O-(trimethylsilyl)- | 9.126 | 1531 | 1518 | 0.86 |
| 5 | Pentane-1,2,3,5-tetraol tetraTMS dec | 9.223 | 1540 | 1526 | 0.92 |
| 6 | 1,2,3,4-tetrakis-o-(trimethylsilyl)pentopyranose | 9.847 | 1600 | 1602 | 0.12 |
| 7 | 6-Deoxy-2,3,4,5-tetrakis-O-(trimethylsilyl)hexose | 9.969 | 1612 | 1632 | 1.23 |
| 8 | Trimethylsilyl p-(trimethylsilyloxy)benzoate | 10.083 | 1624 | 1621 | 0.19 |
| 9 | arabinonic acid, 2,3,5-tris-o-(trimethylsilyl)-, γ-lactone | 10.098 | 1626 | 1647 | 1.28 |
| 10 | D-Arabinopyranose, tetrakis(trimethylsilyl) ether (isomer 2) | 10.154 | 1632 | 1636 | 0.24 |
| 11 | L-(-)-Arabitol, pentakis(trimethylsilyl) ether | 10.842 | 1704 | 1746 | 2.41 |
| 12 | 1,2,3,4,5-Pentakis-O-(trimethylsilyl)pentitol | 11.27 | 1751 | 1746 | 0.29 |
| 13 | (2-(3,4-Bis[(trimethylsilyl)-oxy]phenyl)ethoxy)(trimethyl)silane | 11.327 | 1757 | 1773 | 0.90 |
| 14 | Vanillic acid, trimethylsiloxytrimethylsilyl ester | 11.352 | 1760 | 1750 | 0.57 |
| 15 | L-(-)-Arabitol, pentakis(trimethylsilyl) ether | 11.51 | 1751 | 1746 | 0.29 |
| 16 | D-(-)-Fructofuranose, pentakis(trimethylsilyl) ether (isomer 1) | 11.613 | 1788 | 1792 | 0.22 |
| 17 | D-Psicofuranose, pentakis(trimethylsilyl) ether (isomer 1) | 11.617 | 1788 | 1799 | 0.61 |
| 18 | D-(-)-Fructofuranose, pentakis(trimethylsilyl) ether (isomer 2) | 11.68 | 1796 | 1800 | 0.22 |
| 19 | D-(-)-Fructopyranose, pentakis(trimethylsilyl) ether (isomer 1) | 11.741 | 1802 | 1802 | 0.00 |
| 20 | Ethyl 4-(2-Hydroxymethylphenyl)butanoate | 11.811 | 1810 | 1814 | 0.22 |
| 21 | Benzoic acid, 3,4-bis[(trimethylsilyl)oxy]-, trimethylsilyl ester | 11.831 | 1812 | 1826 | 0.77 |
| 22 | Tetradecanoic acid, trimethylsilyl ester | 12.12 | 1846 | 1842 | 0.22 |
| 23 | D-Psicose, pentakis(trimethylsilyl) ether | 12.351 | 1873 | 1884 | 0.58 |
| 24 | D-(+)-Galactopyranose, pentakis(trimethylsilyl) ether (isomer 2) | 12.401 | 1879 | 1889 | 0.53 |
| 25 | 3-(2-Ethyl-6-methylphenyl)-2-hydroxy-2,4-dimethyloxazolidine | 12.491 | 1890 | 1874 | 0.85 |
| 26 | Trimethylsilyl 3,5-dimethoxy-4-(trimethylsilyloxy)benzoate | 12.514 | 1892 | 1884 | 0.42 |
| 27 | D-Sorbitol, hexakis(trimethylsilyl) ether | 12.701 | 1914 | 1919 | 0.26 |
| 28 | Methyl 2,3,4,6-tetrakis-O-(trimethylsilyl)hexopyranoside | 12.751 | 1920 | 1928 | 0.41 |
| 29 | Hexadecanoic acid, trimethylsilyl ester | 13.763 | 2026 | 2039 | 0.64 |
| 30 | Myo-Inositol, 1,2,3,4,5,6-hexakis-O-(trimethylsilyl)- | 14.027 | 2047 | 2053 | 0.29 |
| 31 | Trimethylsilyl heptadecanoate | 14.527 | 2086 | 2087 | 0.05 |
| 32 | Linoleic acid trimethylsilyl esterI | 15.038 | 2218 | 2201 | 0.77 |
| 33 | cis-13-Octadecenoic acid, trimethylsilyl ester | 14.088 | 2238 | 2232 | 0.27 |
| 34 | Stearoxytrimethylsilane | 15.268 | 2297 | 2277 | 0.88 |
| 35 | 2H-Pyrazino[1’,2’:1,5]pyrrolo[2,3-b]indole-1,4(3H,5aH)-dione, 11a-ethoxy-6,10b,11,11a-tetrahydro-10b-hydroxy-2,6-dimethyl-3-methylene (5aα,10bα,11aβ) | 16.305 | 2654 | 2726 | 2.64 |
| 36 | (E/Z)-5-(N.N-Diethylcarbamoylmethylene)-1-phenyl-3-phenylamino-2,5-dihydropyrrol-2-one | 17.493 | 3062 | 3120 | 1.86 |
| 37 | Sucrose, octakis(trimethylsilyl) ether | 17.813 | 3172 | 3252 | 2.46 |
| 38 | 6H-Isoquino[2,1-b][2,7]naphthyridine, methanone derivative | 17.95 | 3219 | 3266 | 1.44 |
| 39 | D-(+)-Trehalose, octakis(trimethylsilyl) ether | 18.492 | 3560 | 3556 | 0.11 |
| 40 | Docosanoic acid, 13-[[2,3,4,6-tetrakis-O-(trimethylsilyl)-D-glucopyranosyl]oxy] | 20.432 | 4072 | 4108 | 0.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustos-Gómez, C.I.; Gasca-Martínez, D.; Yáñez-Barrientos, E.; Hidalgo-Figueroa, S.; Gonzalez-Rivera, M.L.; Barragan-Galvez, J.C.; Zapata-Morales, J.R.; Isiordia-Espinoza, M.; Corrales-Escobosa, A.R.; Alonso-Castro, A.J. Neuropharmacological Activities of Ceiba aesculifolia (Kunth) Britten & Baker f (Malvaceae). Pharmaceuticals 2022, 15, 1580. https://doi.org/10.3390/ph15121580
Bustos-Gómez CI, Gasca-Martínez D, Yáñez-Barrientos E, Hidalgo-Figueroa S, Gonzalez-Rivera ML, Barragan-Galvez JC, Zapata-Morales JR, Isiordia-Espinoza M, Corrales-Escobosa AR, Alonso-Castro AJ. Neuropharmacological Activities of Ceiba aesculifolia (Kunth) Britten & Baker f (Malvaceae). Pharmaceuticals. 2022; 15(12):1580. https://doi.org/10.3390/ph15121580
Chicago/Turabian StyleBustos-Gómez, Chrystyan Iván, Deisy Gasca-Martínez, Eunice Yáñez-Barrientos, Sergio Hidalgo-Figueroa, Maria L. Gonzalez-Rivera, Juan Carlos Barragan-Galvez, Juan Ramón Zapata-Morales, Mario Isiordia-Espinoza, Alma Rosa Corrales-Escobosa, and Angel Josabad Alonso-Castro. 2022. "Neuropharmacological Activities of Ceiba aesculifolia (Kunth) Britten & Baker f (Malvaceae)" Pharmaceuticals 15, no. 12: 1580. https://doi.org/10.3390/ph15121580
APA StyleBustos-Gómez, C. I., Gasca-Martínez, D., Yáñez-Barrientos, E., Hidalgo-Figueroa, S., Gonzalez-Rivera, M. L., Barragan-Galvez, J. C., Zapata-Morales, J. R., Isiordia-Espinoza, M., Corrales-Escobosa, A. R., & Alonso-Castro, A. J. (2022). Neuropharmacological Activities of Ceiba aesculifolia (Kunth) Britten & Baker f (Malvaceae). Pharmaceuticals, 15(12), 1580. https://doi.org/10.3390/ph15121580