
Aescin Protects against Experimental Benign Prostatic Hyperplasia and Preserves Prostate Histomorphology in Rats via Suppression of Inflammatory Cytokines and COX-2

 , , , and
, , , and
Abstract
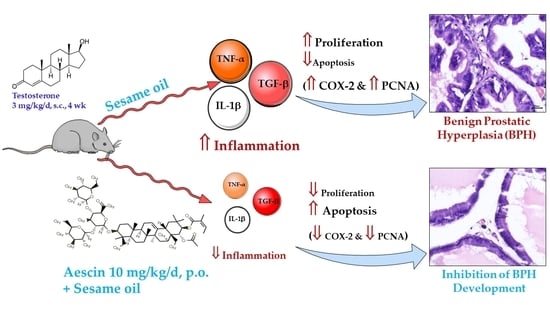
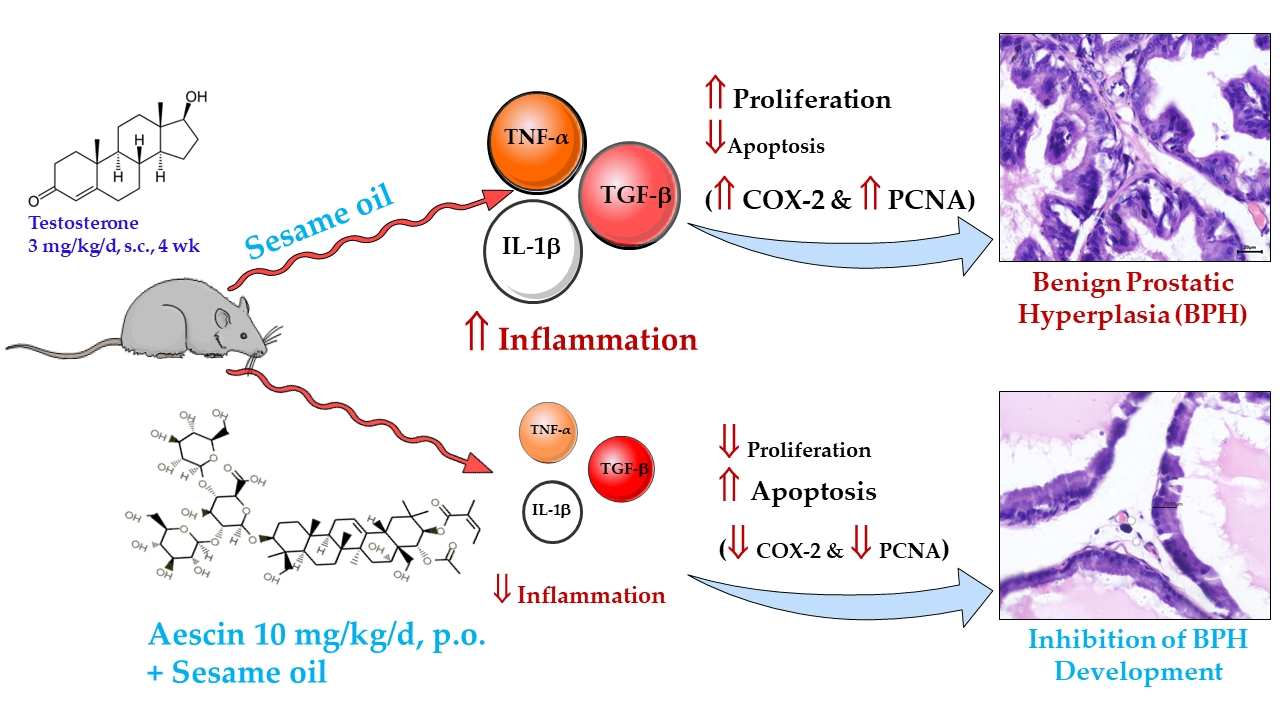
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
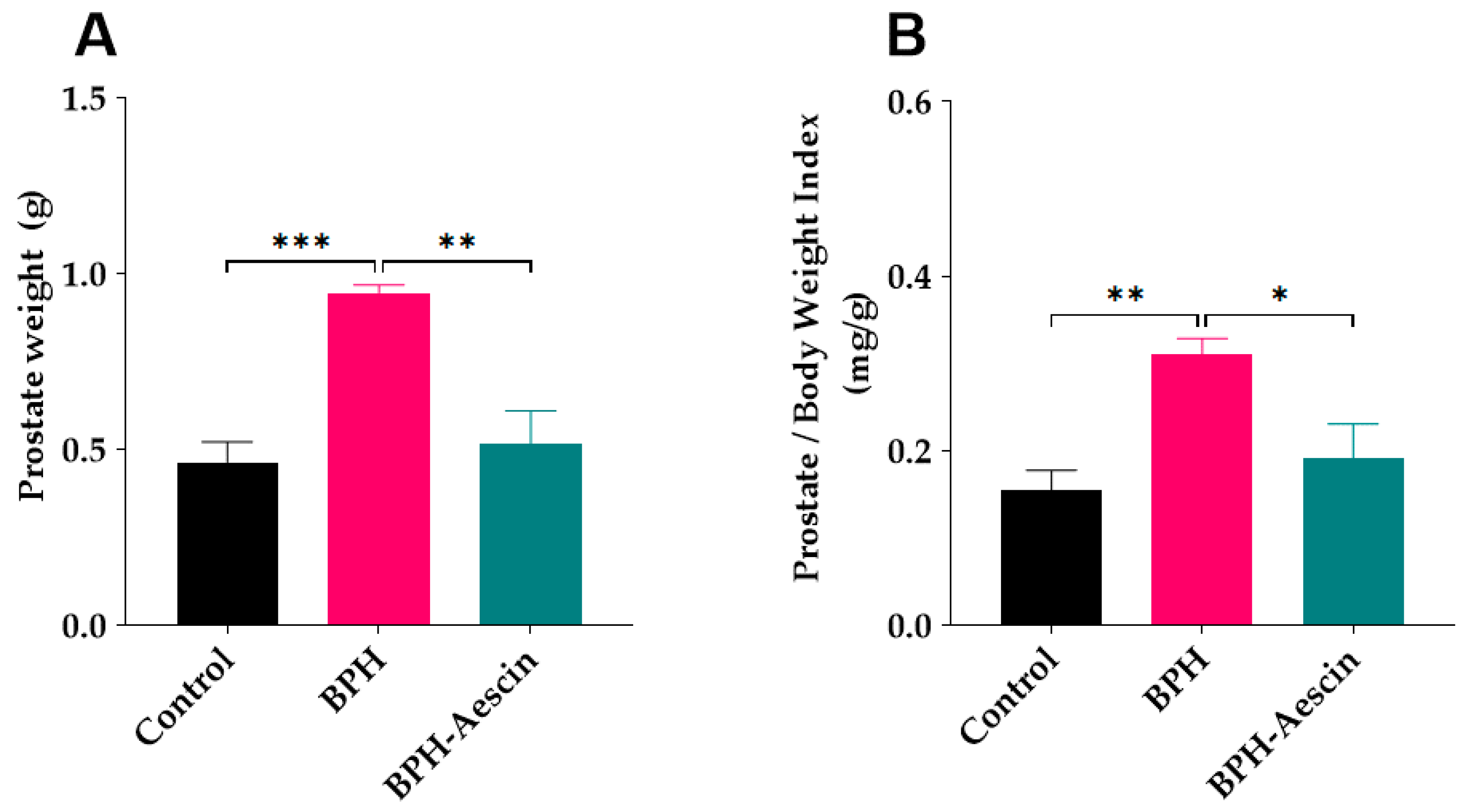
2.1. Effect of Aescin Treatment on Prostate Weight and Prostate Weight Index
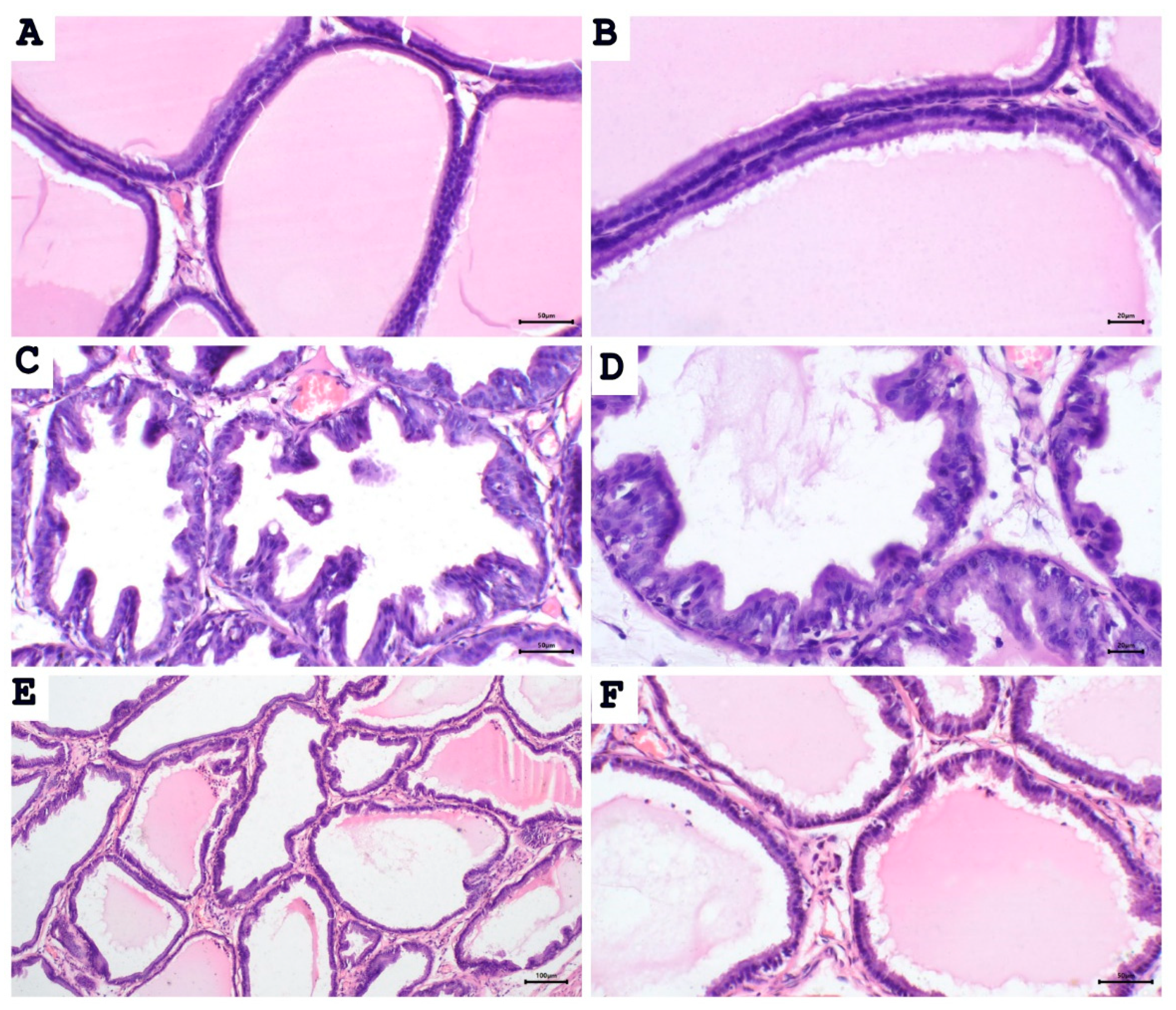
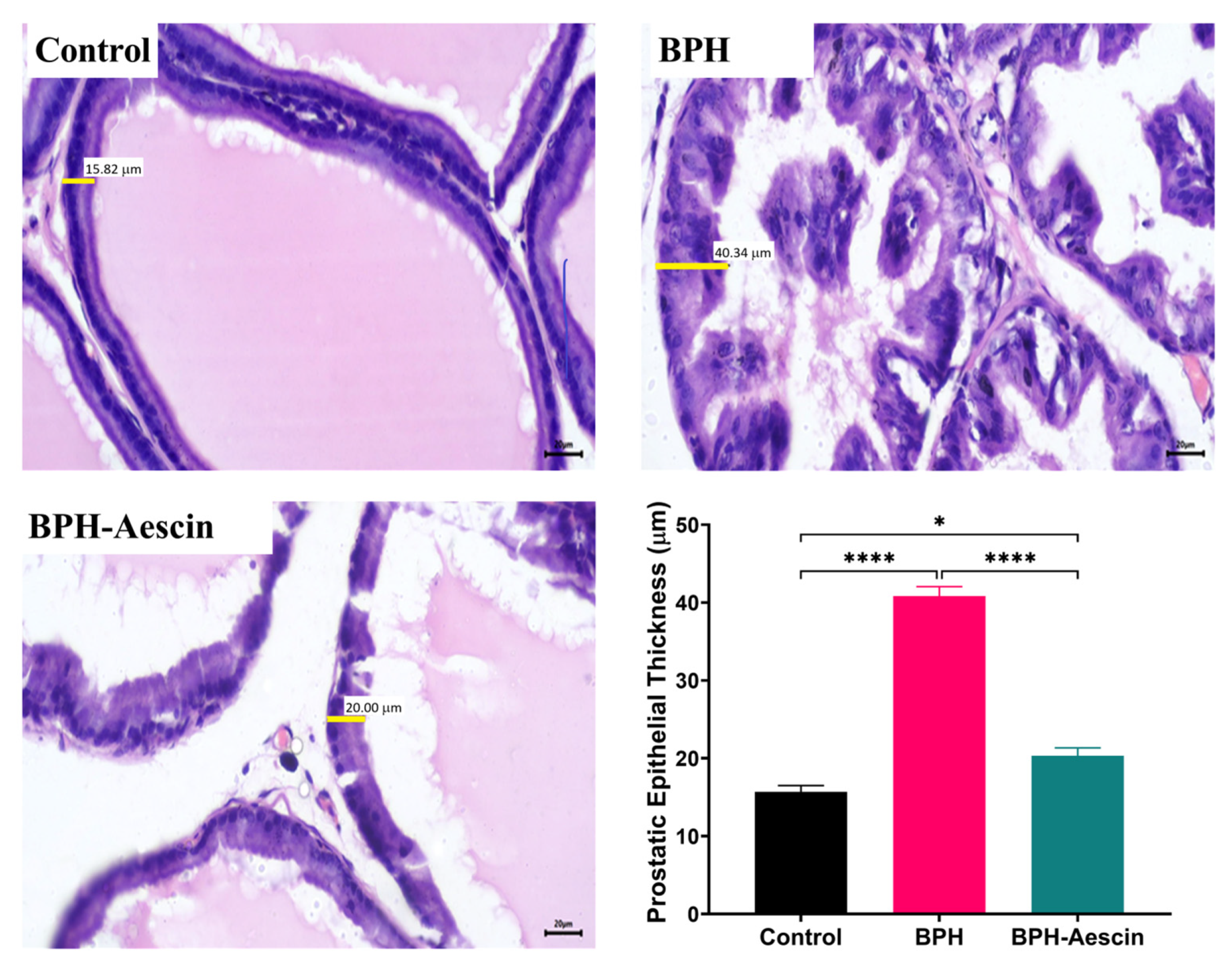
2.2. Aescin Prevents the Testosterone-Induced Prostatic Histopathological Abnormalities
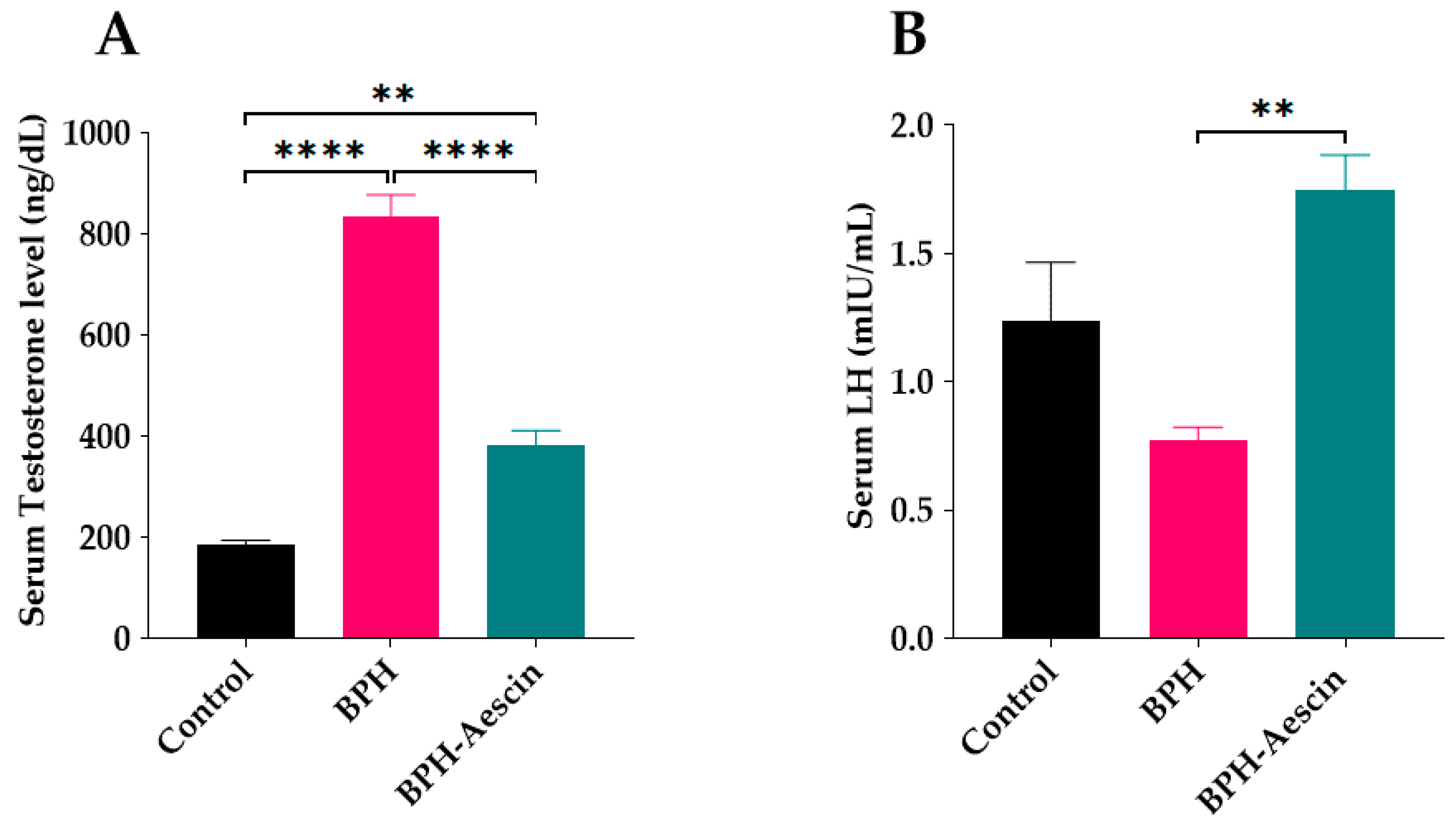
2.3. Effect of Aescin Treatment on Serum Testosterone and Luteinizing Hormone
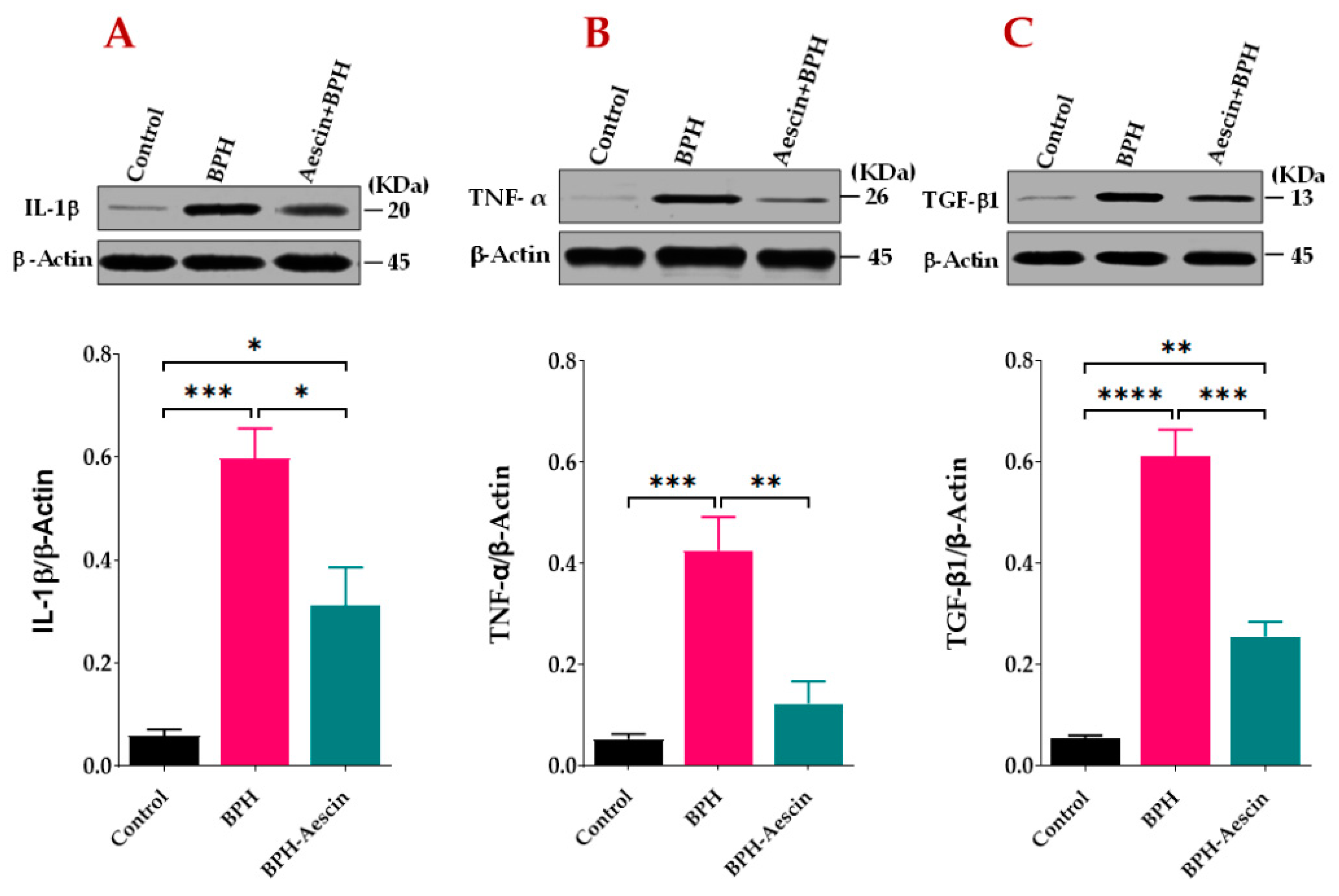
2.4. Effect of Aescin Treatment on Prostatic Expression of IL1-β, TNF-α, and TGF-β1
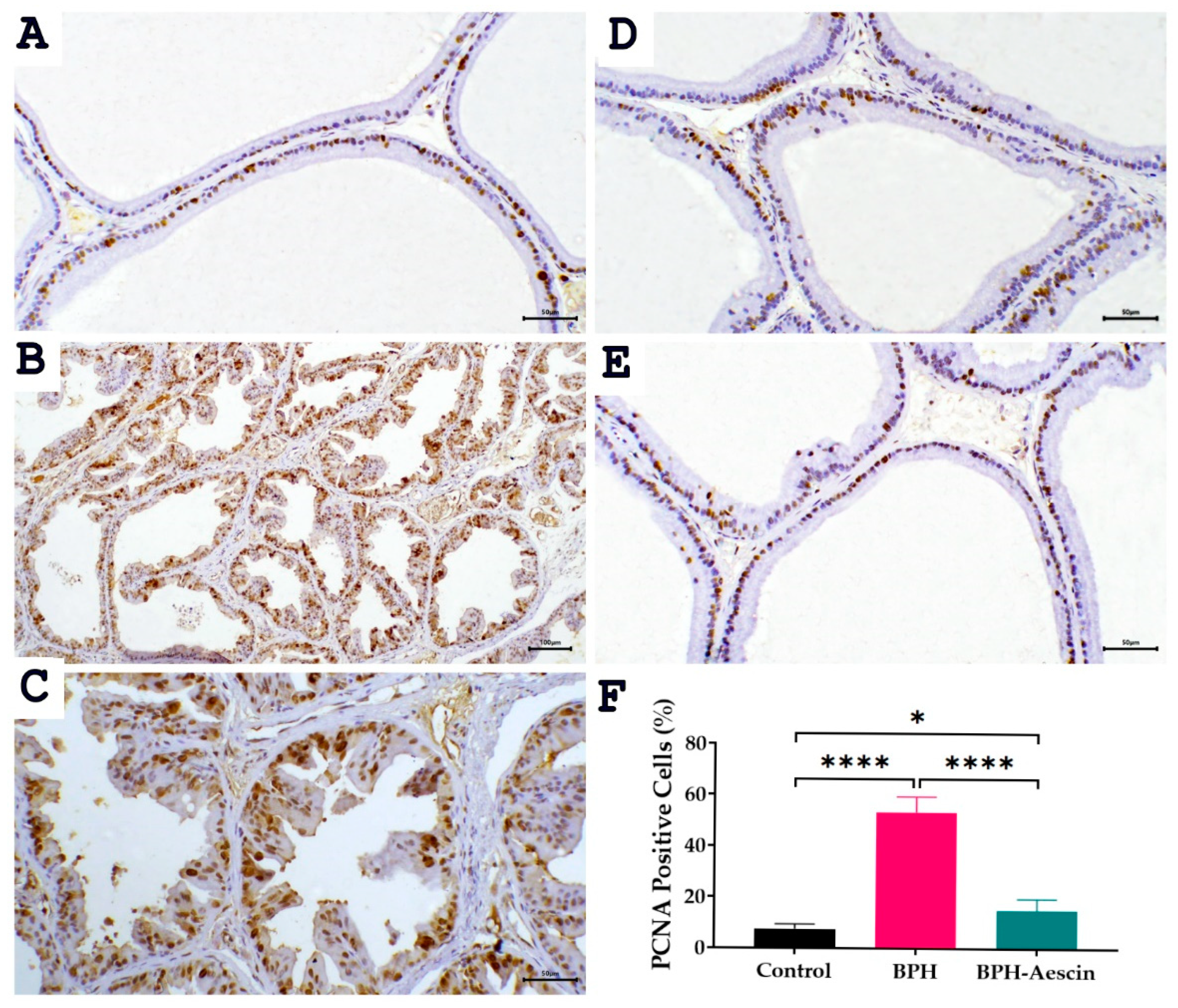
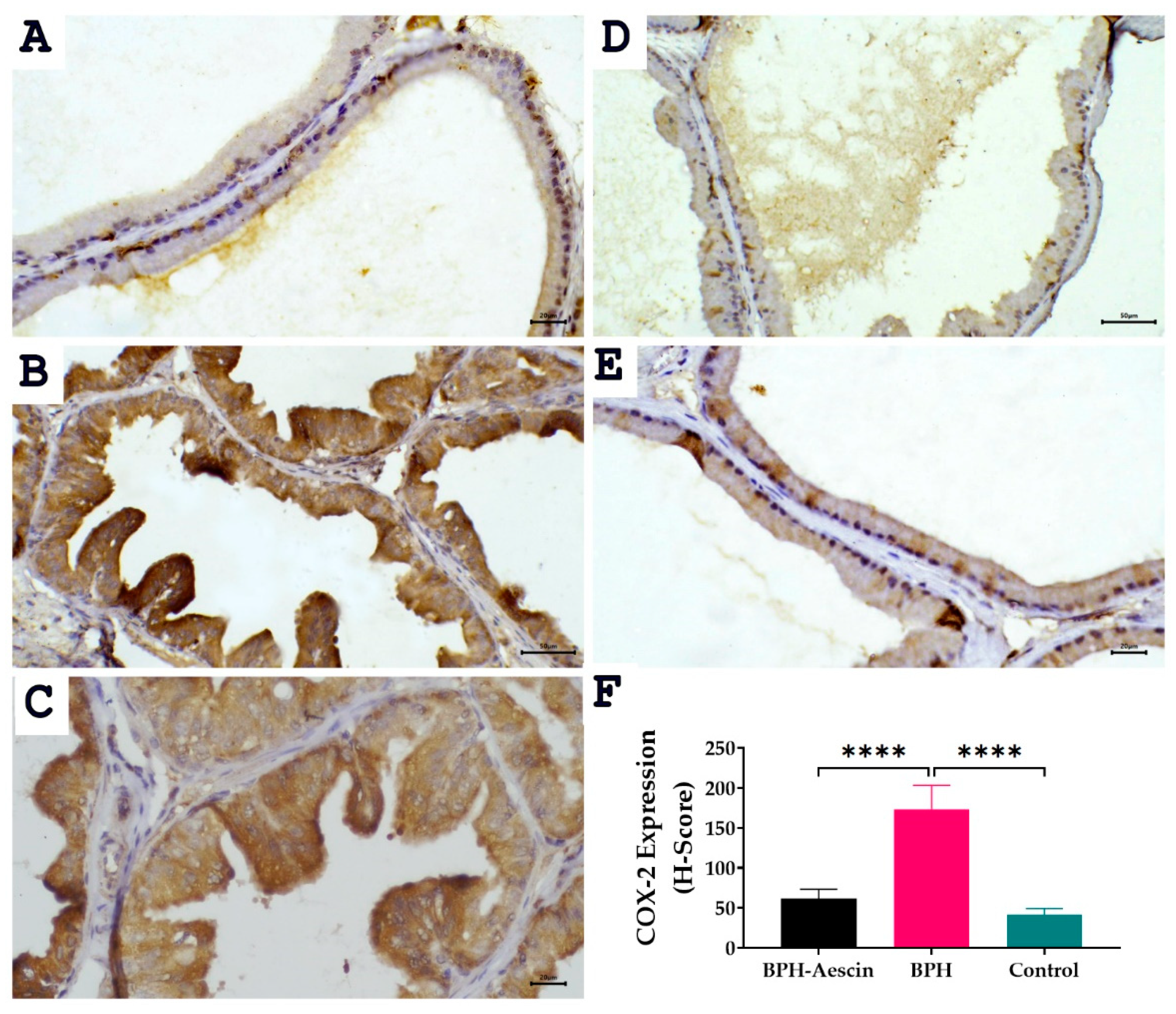
2.5. Effect of Aescin on the Expressions of PCNA and COX-2 in the Prostate of BPH Rats
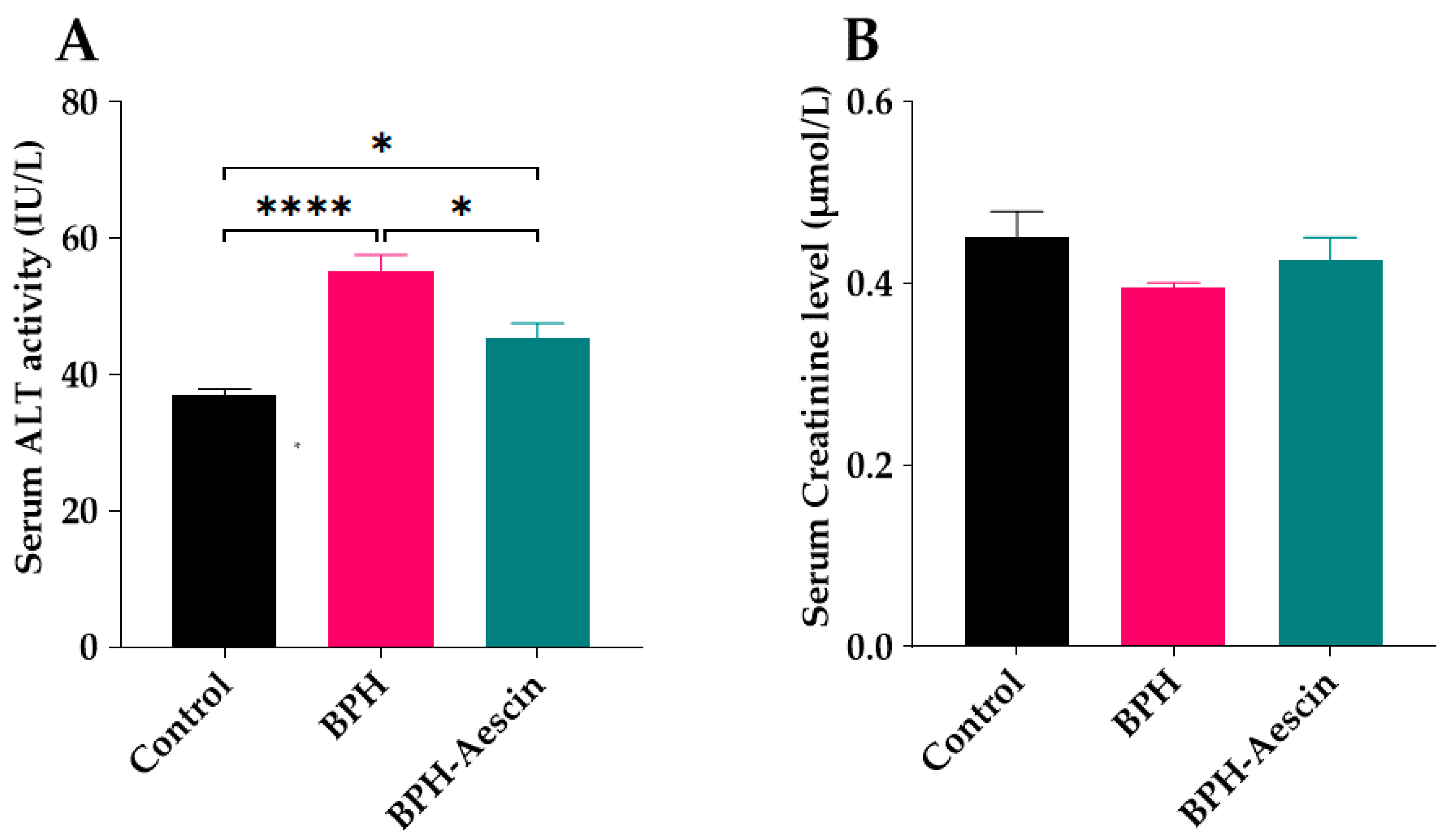
2.6. Effect of Aescin Treatment on Serum ALT Activity and Creatinine Level
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Chemicals
4.3. Experimental Design
4.4. Sample Collection and Preparation
4.5. Determination of Serum Levels of Testosterone and Luteinizing Hormone (LH)
4.6. Western Blotting Analysis of TNF-α, IL-1β, and TGF-β1
4.7. Histopathological Assessment
4.8. Immunohistochemical detection of PCNA and COX-2
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phua, T.J. The Etiology and Pathophysiology Genesis of Benign Prostatic Hyperplasia and Prostate Cancer: A New Perspective. Medicines 2021, 8, 30. [Google Scholar] [CrossRef]
- Park, S.; Ryu, J.-M.; Lee, M. Quality of life in older adults with Benign Prostatic hyperplasia. Healthcare 2020, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Maghsoudi, H.; Hallajzadeh, J.; Rezaeipour, M. Evaluation of the effect of polyphenol of escin compared with ibuprofen and dexamethasone in synoviocyte model for osteoarthritis: An in vitro study. Clin. Rheumatol. 2018, 37, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A. Benign prostatic hyperplasia (BPH) management in the primary care setting. Can. J. Urol. 2012, 19, 10–17. [Google Scholar] [PubMed]
- Traish, A.M. Health Risks Associated with Long-Term Finasteride and Dutasteride Use: It′s Time to Sound the Alarm. World J. Men’s Health 2020, 38, 323–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirshburg, J.M.; Kelsey, P.A.; Therrien, C.A.; Gavino, A.C.; Reichenberg, J.S. Adverse effects and safety of 5-alpha reductase inhibitors (finasteride, dutasteride): A systematic review. J. Clin. Aesthetic Dermatol. 2016, 9, 56. [Google Scholar]
- Madersbacher, S.; Sampson, N.; Culig, Z. Pathophysiology of Benign Prostatic Hyperplasia and Benign Prostatic Enlargement: A Mini-Review. Gerontology 2019, 65, 458–464. [Google Scholar] [CrossRef]
- De Nunzio, C.; Presicce, F.; Tubaro, A. Inflammatory mediators in the development and progression of benign prostatic hyperplasia. Nat. Rev. Urol. 2016, 13, 613–626. [Google Scholar] [CrossRef]
- Devlin, C.M.; Simms, M.S.; Maitland, N.J. Benign prostatic hyperplasia—What do we know? BJU Int. 2021, 127, 389–399. [Google Scholar] [CrossRef]
- Cai, T.; Santi, R.; Tamanini, I.; Galli, I.C.; Perletti, G.; Bjerklund Johansen, T.E.; Nesi, G. Current Knowledge of the Potential Links between Inflammation and Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 3833. [Google Scholar] [CrossRef] [Green Version]
- Vasquez-Velasquez, C.; Gasco, M.; Fano-Sizgorich, D.; Gonzales, G.F. Inflammatory pathway employed by Red Maca to treat induced benign prostatic hyperplasia in rats. Andrologia 2020, 52, e13516. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhou, R.Y. Review of the Roles and Interaction of Androgen and Inflammation in Benign Prostatic Hyperplasia. Mediat. Inflamm. 2020, 2020, 7958316. [Google Scholar] [CrossRef] [PubMed]
- Adhyatma, K.P.; Prapiska, F.F.; Siregar, G.P.; Warli, S.M. Systemic Inflammatory Response in Predicting Prostate Cancer: The Diagnostic Value of Neutrophil-To-Lymphocyte Ratio. Open Access Maced. J. Med. Sci. 2019, 7, 1628–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandaglia, G.; Zaffuto, E.; Fossati, N.; Cucchiara, V.; Mirone, V.; Montorsi, F.; Briganti, A. The role of prostatic inflammation in the development and progression of benign and malignant diseases. Curr. Opin. Urol. 2017, 27, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kohnen, P.W.; Drach, G.W. Patterns of inflammation in prostatic hyperplasia: A histologic and bacteriologic study. J. Urol. 1979, 121, 755–760. [Google Scholar] [CrossRef]
- Bierhoff, E.; Vogel, J.; Benz, M.; Giefer, T.; Wernert, N.; Pfeifer, U. Stromal nodules in benign prostatic hyperplasia. Eur. Urol. 1996, 29, 345–354. [Google Scholar] [CrossRef]
- He, Q.; Wang, Z.; Liu, G.; Daneshgari, F.; MacLennan, G.T.; Gupta, S. Metabolic syndrome, inflammation and lower urinary tract symptoms: Possible translational links. Prostate Cancer Prostatic Dis. 2016, 19, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, P.; Motley, S.S.; Koyama, T.; Kashyap, M.; Gingrich, J.; Yoshimura, N.; Fowke, J.H. Molecular correlates in urine for the obesity and prostatic inflammation of BPH/LUTS patients. Prostate 2018, 78, 17–24. [Google Scholar] [CrossRef]
- Bostanci, Y.; Kazzazi, A.; Momtahen, S.; Laze, J.; Djavan, B. Correlation between benign prostatic hyperplasia and inflammation. Curr. Opin. Urol. 2013, 23, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Naim, A.B.; Neamatallah, T.; Eid, B.G.; Esmat, A.; Alamoudi, A.J.; El-Aziz, A.; Gamal, S.; Ashour, O.M. 2-Methoxyestradiol Attenuates Testosterone-Induced Benign Prostate Hyperplasia in Rats through Inhibition of HIF-1α/TGF-β/Smad2 Axis. Oxidative Med. Cell. Longev. 2018, 2018, 4389484. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Lin, W.-J.; Izumi, K.; Wang, X.; Xu, D.; Fang, L.-Y.; Li, L.; Jiang, Q.; Jin, J.; Chang, C. Targeting androgen receptor to suppress macrophage-induced EMT and benign prostatic hyperplasia (BPH) development. Mol. Endocrinol. 2012, 26, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, Y.; Wang, Z.; O’Malley, K.J.; Tyagi, P.; DeFranco, D.B.; Gingrich, J.R.; Takahashi, R.; Majima, T.; Gotoh, M.; Yoshimura, N. Influence of E. coli-induced prostatic inflammation on expression of androgen-responsive genes and transforming growth factor beta 1 cascade genes in rats. Prostate 2015, 75, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Dai, W.; Li, J.; Li, Y. Synergistic effect of estradiol and testosterone protects against IL-6-induced cardiomyocyte apoptosis mediated by TGF-β1. Int. J. Clin. Exp. Pathol. 2018, 11, 10–26. [Google Scholar] [PubMed]
- Chen, D.; Fan, J.; Li, X.; Jiao, Z.; Tang, G.; Guo, X.; Chen, H.; Wang, J.; Men, T. Downregulation of miR-485-3p promotes proliferation, migration and invasion in prostate cancer through activation of TGF-β signaling. Histol. Histopathol. 2021, 18416. [Google Scholar] [CrossRef]
- Kyprianou, N.; Tu, H.; Jacobs, S.C. Apoptotic versus proliferative activities in human benign prostatic hyperplasia. Hum. Pathol. 1996, 27, 668–675. [Google Scholar] [CrossRef]
- Descazeaud, A.; Weinbreck, N.; Robert, G.; Vacherot, F.; Abbou, C.; Labrousse, F.; Allory, Y.; Rubin, M.; de la Taille, A. Transforming growth factor β-receptor II protein expression in benign prostatic hyperplasia is associated with prostate volume and inflammation. BJU Int. 2011, 108, E23–E28. [Google Scholar] [CrossRef] [Green Version]
- Gallelli, L.; Cione, E.; Wang, T.; Zhang, L. Glucocorticoid-Like Activity of Escin: A New Mechanism for an Old Drug. Drug Des. Dev. Ther. 2021, 15, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Idris, S.; Mishra, A.; Khushtar, M. Phytochemical, ethanomedicinal and pharmacological applications of escin from Aesculus hippocastanum L. towards future medicine. J. Basic Clin. Physiol. Pharmacol. 2020, 31, 20190115. [Google Scholar] [CrossRef]
- Ezberci, F.; Ünal, E. Aesculus Hippocastanum (Aescin, Horse Chestnut) in the Management of Hemorrhoidal Disease. Turk. J. Colorectal Dis. 2018, 28, 54. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Jiang, N.; Wang, Z.; Chong, Y.; Fu, F. Anti-inflammatory effects of escin are correlated with the glucocorticoid receptor/NF-κB signaling pathway, but not the COX/PGF2α signaling pathway. Exp. Ther. Med. 2013, 6, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Ming, Z.J.; Hu, Y.; Qiu, Y.H.; Cao, L.; Zhang, X.G. Synergistic effects of beta-aescin and 5-fluorouracil in human hepatocellular carcinoma SMMC-7721 cells. Phytomedicine: Int. J. Phytother. Phytopharm. 2010, 17, 575–580. [Google Scholar] [CrossRef]
- Liu, Z.R.; Sun, L.Z.; Jia, T.H.; Jia, D.F. β-Aescin shows potent antiproliferative activity in osteosarcoma cells by inducing autophagy, ROS generation and mitochondrial membrane potential loss. J. BUON: Off. J. Balk. Union Oncol. 2017, 22, 1582–1586. [Google Scholar]
- Ramadoss, H.; Kathiresan, S.; Kathiresan, S. Anti-Proliferative Activity of Aescin on Human Oral Carcinoma (KB) Cells and Human Laryngeal Carcinoma (Hep-2) Cells-Via Cell Sensitivity Assays. Ann. Rom. Soc. Cell Biol. 2021, 25, 3559–3573. [Google Scholar]
- Li, B.; Wu, G.L.; Dai, W.; Wang, G.; Su, H.Y.; Shen, X.P.; Zhan, R.; Xie, J.M.; Wang, Z.; Qin, Z.H.; et al. Aescin-induced reactive oxygen species play a pro-survival role in human cancer cells via ATM/AMPK/ULK1-mediated autophagy. Acta Pharmacol. Sin. 2018, 39, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, Z.; Xie, J.M.; Wang, G.; Qian, L.Q.; Guan, X.M.; Shen, X.P.; Qin, Z.H.; Shen, G.H.; Li, X.Q.; et al. TIGAR knockdown enhanced the anticancer effect of aescin via regulating autophagy and apoptosis in colorectal cancer cells. Acta Pharmacol. Sin. 2019, 40, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.M.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, X.; Gan, Y.; Li, B.S.; Wang, K.N.; He, Y. Dihydroartemisinin attenuates benign prostatic hyperplasia in rats by inhibiting prostatic epithelial cell proliferation. Ann. Transl. Med. 2021, 9, 1246. [Google Scholar] [CrossRef]
- Karunasagara, S.; Hong, G.L.; Jung, D.Y.; Kim, K.H.; Cho, K.; Jung, J.Y. Protective effects of combination of Stauntonia hexaphylla and Cornus officinalis on testosterone-induced benign prostatic hyperplasia through inhibition of 5α- reductase type 2 and induced cell apoptosis. PLoS ONE 2020, 15, e0236879. [Google Scholar] [CrossRef] [PubMed]
- An, Y.J.; Lee, J.Y.; Kim, Y.; Jun, W.; Lee, Y.H. Cranberry Powder Attenuates Benign Prostatic Hyperplasia in Rats. J. Med. Food 2020, 23, 1296–1302. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, L.M.; Wang, Y.F.; Yang, H.Y.; Huang, X.Y.; Zhou, W.; Sun, H.X. Effect of sodium aescinate treatment on PCOS rat model with insulin resistance. Bratisl. Lek. Listy 2017, 118, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.B. Nonclassical Testosterone Signaling: A New Pathway Controlling Spermatogenesis? Biol. Reprod. 2016, 94, 43. [Google Scholar] [CrossRef]
- Walker, W.H. Androgen Actions in the Testis and the Regulation of Spermatogenesis. Adv. Exp. Med. Biol. 2021, 1288, 175–203. [Google Scholar] [CrossRef]
- Ammar, A.E.; Esmat, A.; Hassona, M.D.; Tadros, M.G.; Abdel-Naim, A.B.; Guns, E.S.T. The effect of pomegranate fruit extract on testosterone-induced BPH in rats. Prostate 2015, 75, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Lin, Y.S.; Weng, W.C.; Panny, L.; Chen, H.L.; Tung, M.C.; Ou, Y.C.; Lin, C.C.; Yang, C.H. Phloretin Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia in Rats by Regulating the Inflammatory Response, Oxidative Stress and Apoptosis. Life 2021, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lu, C.; Zhang, L.; Zhang, J.; Du, Y.; Duan, S.; Wang, T.; Fu, F. Oral Administration of Escin Inhibits Acute Inflammation and Reduces Intestinal Mucosal Injury in Animal Models. Evid.-Based Complementary Altern. Med.: Ecam 2015, 2015, 503617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Kuang, F.; Ju, G. Aescin reduces oxidative stress and provides neuroprotection in experimental traumatic spinal cord injury. Free. Radic. Biol. Med. 2016, 99, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Torrealba, N.; Rodríguez-Berriguete, G.; Fraile, B.; Olmedilla, G.; Martínez-Onsurbe, P.; Guil-Cid, M.; Paniagua, R.; Royuela, M. Expression of several cytokines in prostate cancer: Correlation with clinical variables of patients. Relationship with biochemical progression of the malignance. Cytokine 2017, 89, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, J.; Li, Y.; Liu, H.; Hassan, H.M.; He, W.; Li, M.; Zhou, Y.; Fu, X.; Zhan, J.; et al. Upregulated bone morphogenetic protein 5 enhances proliferation and epithelial-mesenchymal transition process in benign prostatic hyperplasia via BMP/Smad signaling pathway. Prostate 2021, 81, 1435–1449. [Google Scholar] [CrossRef]
- Shoieb, S.M.; Esmat, A.; Khalifa, A.E.; Abdel-Naim, A.B. Chrysin attenuates testosterone-induced benign prostate hyperplasia in rats. Food Chem. Toxicol. 2018, 111, 650–659. [Google Scholar] [CrossRef]
- Al-Trad, B.; Aljabali, A.; Al Zoubi, M.; Shehab, M.; Omari, S. Effect of gold nanoparticles treatment on the testosterone-induced benign prostatic hyperplasia in rats. Int. J. Nanomed. 2019, 14, 3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y.; Chen, L.; Ren, N.; Li, B.; Wu, Y.; Rankin, G.O.; Rojanasakul, Y.; Wang, Y.; Chen, Y.C. Standardized saponin extract from Baiye no. 1 tea (Camellia sinensis) flowers induced S phase cell cycle arrest and apoptosis via AKT-MDM2-p53 signaling pathway in ovarian cancer cells. Molecules 2020, 25, 3515. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Dwivedi, J.; Jain, P.K.; Satpathy, S.; Patra, A. Medicinal plants for treatment of cancer: A brief review. Pharmacogn. J. 2016, 8, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.Q.; Xu, S.Q.; Cheng, J.; Cao, X.L.; Zhang, Y.; Zhou, W.P.; Huang, Y.J.; Wang, J.; Hu, X.M. Anti-inflammatory effect of external use of escin on cutaneous inflammation: Possible involvement of glucocorticoids receptor. Chin. J. Nat. Med. 2018, 16, 105–112. [Google Scholar] [CrossRef]
- Ding, Y.X.; Eerduna, G.W.; Duan, S.J.; Li, T.; Liu, R.X.; Zhang, L.M.; Wang, T.; Fu, F.H. Escin ameliorates the impairments of neurological function and blood brain barrier by inhibiting systemic inflammation in intracerebral hemorrhagic mice. Exp. Neurol. 2021, 337, 113554. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhao, S.; Wang, Y.; Yang, Y.; Yao, L.; Chu, L.; Du, H.; Fu, F. Protective effects of escin against indomethacin-induced gastric ulcer in mice. Toxicol. Mech. Methods 2014, 24, 560–566. [Google Scholar] [CrossRef]
- Segain, J.-P.; de la Blétière, D.R.; Sauzeau, V.; Bourreille, A.; Hilaret, G.; Cario-Toumaniantz, C.; Pacaud, P.; Galmiche, J.-P.; Loirand, G. Rho kinase blockade prevents inflammation via nuclear factor κB inhibition: Evidence in Crohn’s disease and experimental colitis. Gastroenterology 2003, 124, 1180–1187. [Google Scholar] [CrossRef]
- Jiang, N.; Xin, W.; Wang, T.; Zhang, L.; Fan, H.; Du, Y.; Li, C.; Fu, F. Protective effect of aescin from the seeds of Aesculus hippocastanum on liver injury induced by endotoxin in mice. Phytomedicine: Int. J. Phytother. Phytopharm. 2011, 18, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Zhang, L.; Fan, H.; Jiang, N.; Wang, T.; Fu, F. Escin attenuates acute lung injury induced by endotoxin in mice. Eur. J. Pharm. Sci.: Off. J. Eur. Fed. Pharm. Sci. 2011, 42, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, G.; Kaufman, M.J.; Pope, H.G., Jr. Public health impact of androgens. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 218–223. [Google Scholar] [CrossRef]
- Mullen, C.; Whalley, B.J.; Schifano, F.; Baker, J.S. Anabolic androgenic steroid abuse in the United Kingdom: An update. Br. J. Pharmacol. 2020, 177, 2180–2198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H.; Fan, H.; Wang, T.; Jiang, N.; Yu, P.; Fu, F. The potent anti-inflammatory agent escin does not increase corticosterone secretion and immune cell apoptosis in mice. Fitoterapia 2011, 82, 861–867. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Wang, T.; Jiang, N.; Yu, P.; Liu, F.; Chong, Y.; Fu, F. Potent anti-inflammatory agent escin does not affect the healing of tibia fracture and abdominal wound in an animal model. Exp. Ther. Med. 2012, 3, 735–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, J.Y.; Zhang, M.J.; Wang, Y.Y.; Liu, Y.H. The positive clinical therapeutically effects of Escin on advanced thyroid cancer. Cancer Med. 2017, 6, 937–943. [Google Scholar] [CrossRef]
- Du, Y.; Song, Y.; Zhang, L.; Zhang, M.; Fu, F. Combined treatment with low dose prednisone and escin improves the anti-arthritic effect in experimental arthritis. Int. Immunopharmacol. 2016, 31, 257–265. [Google Scholar] [CrossRef]
- Ewees, M.G.; Messiha, B.A.S.; Abdel-Bakky, M.S.; Bayoumi, A.M.A.; Abo-Saif, A.A. Tempol, a superoxide dismutase mimetic agent, reduces cisplatin-induced nephrotoxicity in rats. Drug. Chem. Toxicol. 2019, 42, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Specht, E.; Kaemmerer, D.; Sänger, J.; Wirtz, R.M.; Schulz, S.; Lupp, A. Comparison of immunoreactive score, HER2/neu score and H score for the immunohistochemical evaluation of somatostatin receptors in bronchopulmonary neuroendocrine neoplasms. Histopathology 2015, 67, 368–377. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raafat, M.; Kamel, A.A.; Shehata, A.H.; Ahmed, A.-S.F.; Bayoumi, A.M.A.; Moussa, R.A.; Abourehab, M.A.S.; El-Daly, M. Aescin Protects against Experimental Benign Prostatic Hyperplasia and Preserves Prostate Histomorphology in Rats via Suppression of Inflammatory Cytokines and COX-2. Pharmaceuticals 2022, 15, 130. https://doi.org/10.3390/ph15020130
Raafat M, Kamel AA, Shehata AH, Ahmed A-SF, Bayoumi AMA, Moussa RA, Abourehab MAS, El-Daly M. Aescin Protects against Experimental Benign Prostatic Hyperplasia and Preserves Prostate Histomorphology in Rats via Suppression of Inflammatory Cytokines and COX-2. Pharmaceuticals. 2022; 15(2):130. https://doi.org/10.3390/ph15020130
Chicago/Turabian StyleRaafat, Mohamed, Amr A. Kamel, Alaa H. Shehata, Al-Shaimaa F. Ahmed, Asmaa M. A. Bayoumi, Rabab A. Moussa, Mohammed A. S. Abourehab, and Mahmoud El-Daly. 2022. "Aescin Protects against Experimental Benign Prostatic Hyperplasia and Preserves Prostate Histomorphology in Rats via Suppression of Inflammatory Cytokines and COX-2" Pharmaceuticals 15, no. 2: 130. https://doi.org/10.3390/ph15020130
APA StyleRaafat, M., Kamel, A. A., Shehata, A. H., Ahmed, A. -S. F., Bayoumi, A. M. A., Moussa, R. A., Abourehab, M. A. S., & El-Daly, M. (2022). Aescin Protects against Experimental Benign Prostatic Hyperplasia and Preserves Prostate Histomorphology in Rats via Suppression of Inflammatory Cytokines and COX-2. Pharmaceuticals, 15(2), 130. https://doi.org/10.3390/ph15020130