
Understanding the Anti-Diarrhoeal Properties of Incomptines A and B: Antibacterial Activity against Vibrio cholerae and Its Enterotoxin Inhibition

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Antibacterial Activity, Cholera Toxin-Induced Diarrhoea, and SDS-PAGE Analysis on Cholera Toxin of Sesquiterpene Lactones from Decachaeta incompta
2.2. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis on Cholera Toxin of Incomptine A (IA) and Incomptine B (IB)
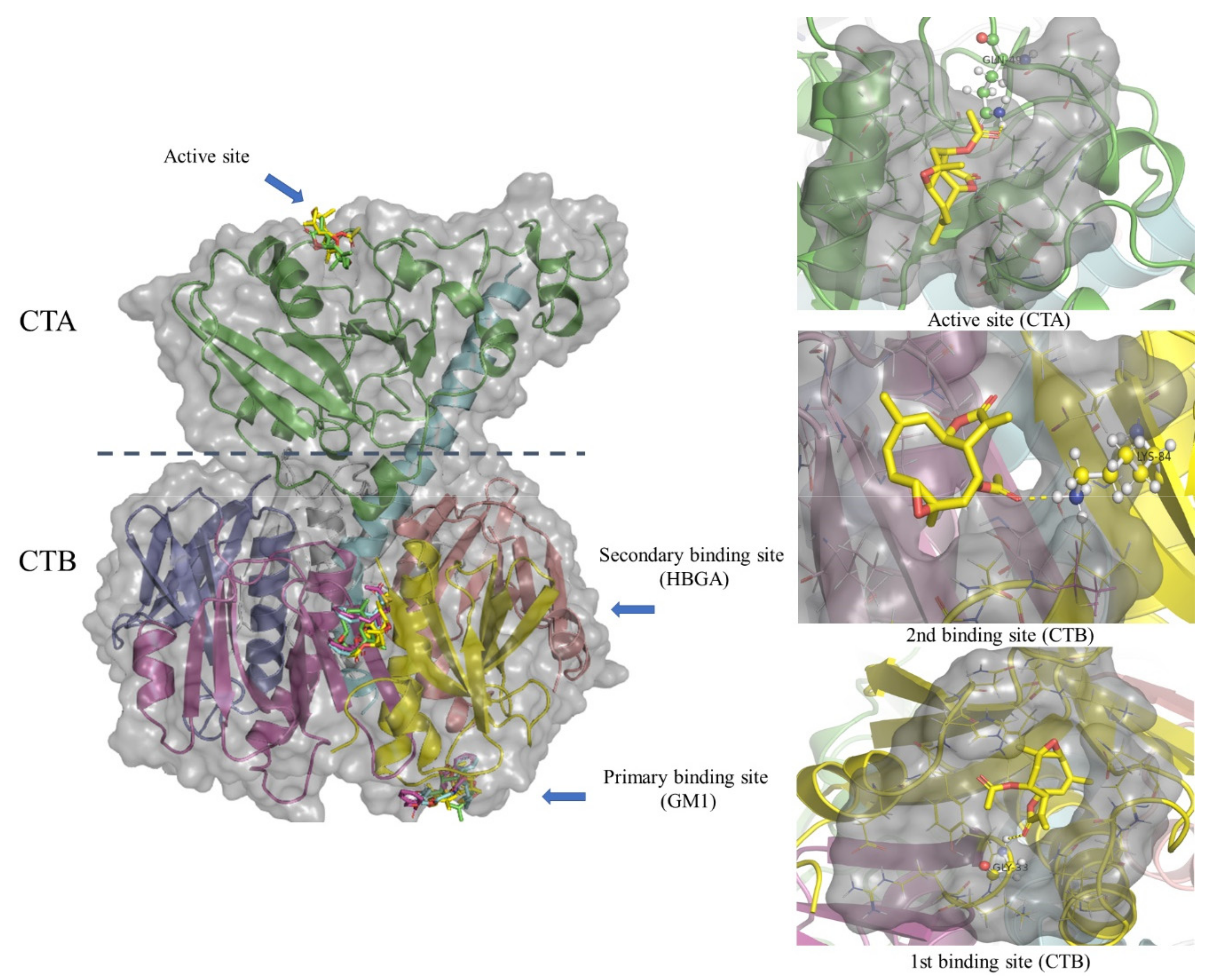
2.3. Molecular Docking Studies of Incomptine A (IA), Incomptine B (IB), and Racecadotril (REC) on Cholera Toxin
3. Discussion
4. Materials and Methods
4.1. Collection and Identification of Decachaeta incompta
4.2. Animals
4.3. Chemicals
4.4. Isolation of Sesquiterpene Lactones of the Aerial Parts from Decachaeta incompta
4.5. Antibacterial Assay
4.6. Cholera Toxin-Induced Diarrhoea
4.7. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.8. Statistical Analysis
5. Molecular Docking Analysis
Molecular Docking of Incomptine A (IA), Incomptine B (IB), and Racecadotril (REC)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casburn-Jones, A.C.; Farthing, M.J. Management of Infectious Diarrhoea. Gut 2004, 53, 296–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UNICEF/WHO 2009. Diarrhoea: Why Children are Still Dying and What Can be Done. WHO. Available online: http://apps.who.int.iris/bitstream/10665/4174/1/9789241598415_eng.pdf (accessed on 22 November 2016).
- Sánchez, J.; Holmgren, J. Virulence Factors, Pathogenesis and Vaccine Protection in Cholera and ETEC Diarrhea. Curr. Opin. Immunol. 2005, 17, 388–398. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cholera. 2018. Available online: http://www.who.int/cholera/task_force/GTFCC-Guidance-cholera-surveillance.pdf?ua=1 (accessed on 25 November 2021).
- SS (Secretaria de Salud). 2008. Sistema Nacional de Vigilancia Epidemiológica. Epidemiología 34. Semana 34. ISSN 1405-2636. Available online: www.sinave.gob.mx (accessed on 2 February 2022).
- Komiazyk, M.; Palczewska, M.; Sitkiewicz, I.; Pikula, S.; Groves, P. Neutralization of Cholera Toxin by Rosaceae Family Plant Extracts. BMC Complement. Altern. Med. 2019, 19, 140. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Nelson, A.R.; Lopez, A.L.; Sack, D. Update Global Burden of Cholera in Endemic Countries. PLoS. Negl. Trop. Dis. 2015, 9, e0003832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G. Application of Sesquiterpene Lactone: A New Promising Way for Cancer Therapy Based on Anticancer Activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Adekenov, S.M. Natural Sesquiterpene Lactones as Renewable Chemical Materials for New Medicinal Products. Eurasian Chem. Technol. J. 2013, 15, 163–174. [Google Scholar] [CrossRef]
- Turner, B.L. A New Species of Decachaeta (Asteraceae: Eupatoriae), from Oaxaca, Mexico. Phytologia 2011, 93, 346–350. [Google Scholar]
- Huang, R.; Baker, H. Healing from the Forest: An Ethnobotanical and Chemical View of Guaymi Medicinal Plants; Results of the Ethnobiology 2001 Course in Costa Rica; Organization for Tropical Studies, Undergraduate Studies Abroad Programme: San José, Costa Rica, 2001; pp. 358–364. [Google Scholar]
- Calzada, F.; Yepez-Mulia, L.; Tapia-Contreras, A.; Ortega, A. Antiprotozoal and Antibacterial Properties of Decachaeta incompta. Rev. Latinoam. Quim. 2009, 37, 98–103. [Google Scholar]
- Bautista, E.; Calzada, F.; Lopez-Huerta, F.A.; Yepez-Mulia, L.; Ortega, O. Antiprotozoal Activity of 8-acyl and 8-alkyl Incomptine A Analogs. Bioorg. Med. Chem. 2014, 24, 3260–3262. [Google Scholar] [CrossRef]
- Rodriguez, E.; Towers, G.H.N.; Mitchell, J.C. Biological activities of Sesquiterpene Lactones. Phytochemistry 1976, 15, 1573–1580. [Google Scholar] [CrossRef]
- Repetto, M.G.; Boveris, A. Bioactivity of Sesquiterpenes: Compounds that Protec from Alcohol-Induced Gastric Mucosomal Lesions and Oxidative Damage. Mini-Rev. Med. Chem. 2010, 10, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, A.; Datta, A. Identification of Inhibitors against the Potential Ligandable Sites in the Active Cholera Toxin. Comput. Biol. Chem. 2015, 55, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Heim, J.B.; Hodnik, V.; Heggelund, J.E.; Anderluh, G.; Krengel, U. Crystal Structures of Cholera Toxin in Complex with Fucosylated Receptors Point to Importance of Secondary Binding Site. Sci. Rep. 2019, 9, 12243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzada, F.; Valdes, M.; Barbosa, E.; Velazquez, C.; Bautista, E. Evaluation of Antipropulsive Activity of Decachaeta incompta (DC) King and Robinson and its Sesquiterpene Lactones on Induced Hyperperistalsis in Rats. Pharmacogn. Mag. 2020, 16, 272–275. [Google Scholar] [CrossRef]
- Wendel, G.H.; Maria, A.O.M.; Guzman, J.A.; Giordano, O.; Pelzer, L.E. Antidiarrheal Activity of Dehydroleucodine Isolated from Artemisia douglasiana. Fitoterapia 2008, 79, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Duraipandiyan, V.; Al-Harbi, N.A.; Ignacimuthu, S.; Muthukumar, C. Antimicrobial Activity of Sesquiterpene Lactones Isolated from Traditional Medicinal Plants, Costus speciosus (Koen ex. Retz.) Sm. BMC Complement. Altern. Med. 2012, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Bautista, E.; Ortega Hernandez, A.; Macias, F.A. Phytotoxicity Evaluation of Sesquiterpene Lactones and Diterpenes from Species of the Decachaeta, Salvia and Podachaenium Genera. Phytochem. Lett. 2016, 18, 68–76. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999. Norma Oficial Mexicana NOM-062-ZOO-1999: Especificaciones Técnicas Para la Producción, Cuidado y uso de los Animales de Laboratorio. 1999. Available online: http://publico.senasica.gob.mx/?doc=743 (accessed on 7 December 2021).
- Torres, J.; González-Arroyo, S.; Pérez, R.; Muñoz, O. Inappropriate Treatment in Children with Bloody Diarrhea: Clinical and Microbiological Studies. Arch. Med. Res. 1995, 26, 23–29. [Google Scholar]
- Alanís, A.D.; Calzada, F.; Cervantes, J.A.; Torres, J.; Ceballos, G.M. Antibacterial Properties of Some Plants Used in Mexican Traditional Medicine for the Treatment of Gastrointestinal Disorders. J. Ethnopharmacol. 2005, 100, 153–157. [Google Scholar] [CrossRef]
- Laemmli, U. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Molecular Operating Environment (MOE), 2019.01; Chemical Computing Group ULC: Montreal, QC, Canada, 2021.
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, G.; Evrard-Todeschi, N.; Jean-Pierre, G.; Bertho, G. Automatic Clustering of Docking Poses in Virtual Screening Process Using Self-organizing map. Bioinformatics. 2010, 26, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molecular Operating Environment (MOE), 2020.09; Chemical Computing Group ULC: Montreal, QC, Canada, 2022.
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Vibrio cholerae | Cholera Toxin-Induced Diarrhoea |
|---|---|---|
| MIC (mg/mL) a | ED50 (mg/kg) b | |
| Incomptine A (IA) | 0.15 | 8.1 ± 1.3 |
| Incomptine B (IB) | 0.05 | 80.0 ± 3.7 |
| Chloramphenicol (CLO) | > 2.0 | - |
| Racecadotril (REC) | 71.8 ± 2.7 |
| Compound | Subunit A (CTA) | Subunit B (CTA) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Active Site | Primary Binding Site (GM1) | Secondary Binding Site (HBGA) | |||||||
| ΔG a | H-BR | NPI | ΔG | H-BR | NPI | ΔG | H-BR | NPI | |
| IA | −5.9 | Gln49 | Ser61, Leu71, Thr75 | −6.6 | Gly33 | Tyr12, His13, Asn14 | −6.5 | Lys84 | Gln3, Asn4, Ile5 |
| Glu110, Glu112 | Ala32, Glu51, Glu56 | Phe25, Ser26, Thr28 | |||||||
| His57, Glu61, Trp88 | Thr41, Ile47, Glu83 | ||||||||
| Lys91 | Ser100, Ala102 | ||||||||
| IB | −6.6 | Gln49 | Ser61, Leu71, Thr75 | −6.1 | -- | Tyr12, His13, Asn14 | −6.1 | -- | Gln3, Asn4, Ile5 |
| Arg54 | Glu110, Glu112 | Ala32, Glu51, Glu56 | Phe25, Ser26, Thr28 | ||||||
| His57, Glu61, Trp88 | Thr41, Ile47, Glu83 | ||||||||
| Lys91 | Ser100, Ala102 | ||||||||
| R-REC | −5.8 | --- | Ser61, Ser68, Leu71 | −6.8 | Glu61 | Tyr12, His13, Asn14 | −5.3 | Thr41 | Gln3, Asn4, Ile5 |
| Thr75, Asp109, Glu110 | Trp88 | Ala32, Glu51, Glu56 | Phe25, Ser26, Thr28 | ||||||
| Glu112 | His57, Ile58, Lys91 | Ile47, Glu83, Ser100 | |||||||
| Ala102 | |||||||||
| S-REC | −5.7 | -- | Ser61, Ser68, Leu71 | −7.0 | His13 | Tyr12, Asn14, Ala32 | −5.4 | Thrt41 | Gln3, Asn4, Ile5 |
| Thr75, Asp109, Glu110 | Trp88 | Glu51, Glu56, His57 | Phe25, Ser26, Thr28 | ||||||
| Glu112 | Ile58, Gln61, Lys91 | Ile47, Glu83, Ser100 | |||||||
| Ala102 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calzada, F.; Bautista, E.; Hidalgo-Figueroa, S.; García-Hernández, N.; Velázquez, C.; Barbosa, E.; Valdes, M.; Solares-Pascasio, J.I. Understanding the Anti-Diarrhoeal Properties of Incomptines A and B: Antibacterial Activity against Vibrio cholerae and Its Enterotoxin Inhibition. Pharmaceuticals 2022, 15, 196. https://doi.org/10.3390/ph15020196
Calzada F, Bautista E, Hidalgo-Figueroa S, García-Hernández N, Velázquez C, Barbosa E, Valdes M, Solares-Pascasio JI. Understanding the Anti-Diarrhoeal Properties of Incomptines A and B: Antibacterial Activity against Vibrio cholerae and Its Enterotoxin Inhibition. Pharmaceuticals. 2022; 15(2):196. https://doi.org/10.3390/ph15020196
Chicago/Turabian StyleCalzada, Fernando, Elihu Bautista, Sergio Hidalgo-Figueroa, Normand García-Hernández, Claudia Velázquez, Elizabeth Barbosa, Miguel Valdes, and Jesús Iván Solares-Pascasio. 2022. "Understanding the Anti-Diarrhoeal Properties of Incomptines A and B: Antibacterial Activity against Vibrio cholerae and Its Enterotoxin Inhibition" Pharmaceuticals 15, no. 2: 196. https://doi.org/10.3390/ph15020196
APA StyleCalzada, F., Bautista, E., Hidalgo-Figueroa, S., García-Hernández, N., Velázquez, C., Barbosa, E., Valdes, M., & Solares-Pascasio, J. I. (2022). Understanding the Anti-Diarrhoeal Properties of Incomptines A and B: Antibacterial Activity against Vibrio cholerae and Its Enterotoxin Inhibition. Pharmaceuticals, 15(2), 196. https://doi.org/10.3390/ph15020196