
Effect and Mechanism of Herbal Medicines on Cisplatin-Induced Anorexia



Abstract
:1. Introduction
2. Results
2.1. 5-HT and Its Receptors
2.2. Inflammatory Cytokines

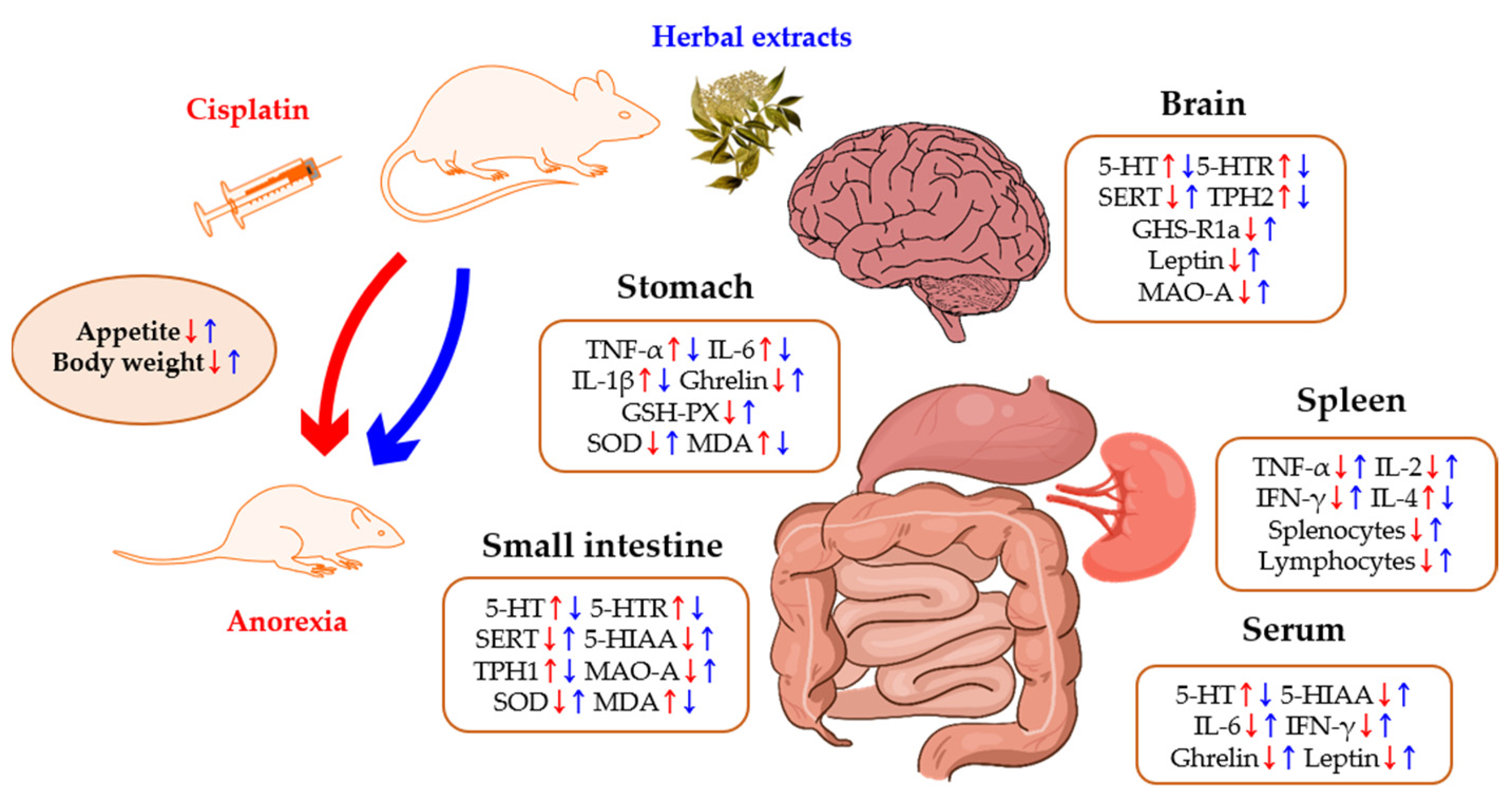{kind=link}
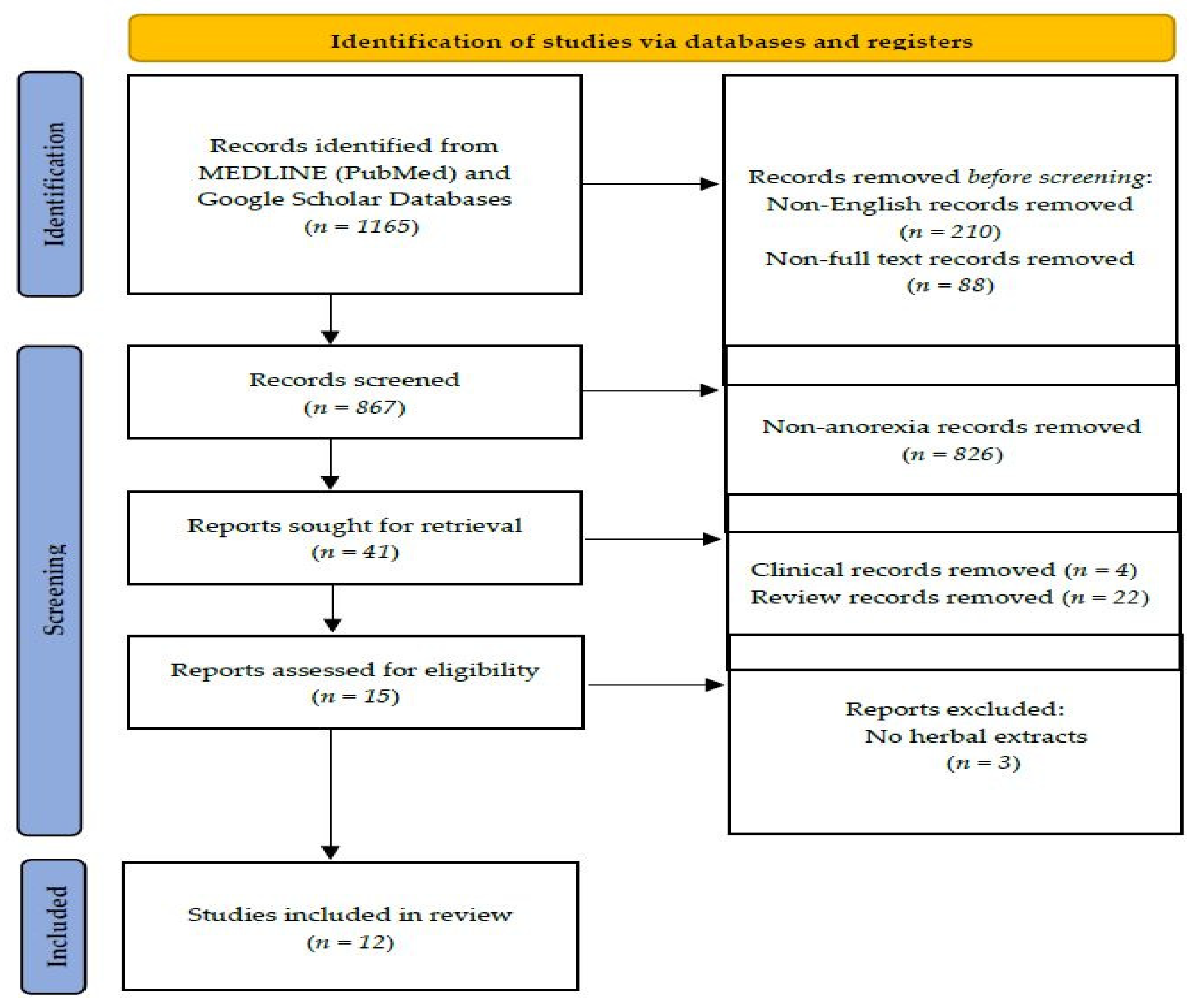{kind=link}
| Authors/ Years | Animal Type | Cisplatin Dosing | Herbal Medicine/ Dose | Findings | ||||
|---|---|---|---|---|---|---|---|---|
| Group | Food Intake | Kaolin Intake | Body Weight | Mechanisms of Actions | ||||
| Aung et al. 2003 [75] | Wistar rats | Single 3 mg/kg, i.p. | Scutellaria baicalensis roots [SbE] (water extract) 1, 3, and 10 mg/kg, i.p. | Cisplatin (n = 7) | ↓ | ↑ | - | - |
| Cisplatin + SbE (n = 6) | ↑ | ↓ | - | - | ||||
| Mehendale et al. 2005 [76] | Wistar rats | Single 3 mg/kg, i.p. | American ginseng berries [AGBE] (75% ethanol extract) 50, 100, and 150 mg/kg, i.p. | Cisplatin (n = 6) | ↓ | ↑ | - | - |
| Cisplatin + AGBE (n = 6) | ↑ | ↓ | - | - | ||||
| Cisplatin + Ginsenoside Re (5 mg/kg, i.p.) (n = 6) | - | ↓ | - | - | ||||
| Takeda et al. 2008 [45] | SD rats | Single 2 mg/kg, i.p. | Rikkunshito [RKT]; Atractylodis Lanceae Rhizoma, Ginseng Radix, Pinelliae Tuber, Hoelen, Zizyphi Fructus, Aurantii Nobilis Pericarpium, Glycyrrhizae Radix, Zingiberis Rhizoma). (water extract) 500, 1000 mg/kg, p.o. | Cisplatin (n = 4–5) | ↓ | - | - | Plasma-acylated ghrelin↓ Plasma-desacylated ghrelin↓ |
| 5-HT (4 & 8 mg/kg, i.p.) (n = 4–5) | - | - | - | Plasma-acylated ghrelin↓ Plasma-desacylated ghrelin↓ | ||||
| Cisplatin + RKT (n = 4–5) | ↑ | - | - | Plasma-acylated ghrelin↑ | ||||
| Cisplatin + RKT + GHS-R1a antagonist (0.4 μmol/rat, i.p.) (n = 4–5) | ↓ | - | - | - | ||||
| Cisplatin + HMF or Hesperetin or Isoliquiritigenin (n = 4–5) | - | - | - | Plasma-acylated ghrelin↑ | ||||
| HMF, Hesperetin, Isoliquiritigenin (n = 4–5) | - | - | - | 5-HT2B receptor: Ki: 0.21 ± 0.01 & 5.3 ± 0.1 & 3.3 μmol/L | ||||
| Hesperetin, Isoliquiritigenin (n = 4–5) | - | - | - | 5-HT2C receptor: Ki: 20.9 ± 7.1 & 3.5 ± 0.1 μmol/L | ||||
| Yakabi et al. 2010 [46] | SD rats | Single 2 mg/kg, i.p. | [RKT] (water extract) 500, 1000 mg/kg, p.o. | Cisplatin (n = 4–5) | ↓ | - | - | Hypothalamic GHS-R1a↓ |
| Cisplatin + Ghrelin (2 nmol/rat, I.C.V.) (n = 4–5) | ↓ | - | - | - | ||||
| 5-HT2C receptor agonist (9 mg/kg, i.p.) (n = 4–5) | ↓ | - | - | - | ||||
| 5-HT2C receptor agonist + RKT (n = 4–5) | ↑ | - | - | - | ||||
| Cisplatin + RKT (n = 4–5) | ↑ | - | - | Hypothalamic GHS-R1a↑ Effect abolished by GHS-R1a antagonist (1nmol/rat, I.C.V.) | ||||
| Cisplatin + Hesperidin or Isoliquiritigenin (n = 4–5) | ↑ | - | - | Effect abolished by GHS-R1a antagonist (1 nmol/rat, I.C.V.) | ||||
| Raghavendran et al. 2011 [77] | SD rats | Single Pre—7 mg/kg Post—6 mg/kg, i.p. | Korean ginseng roots [KG; the root of Panax ginseng CA Meyer] (water extract) Pre—25, 50, 100 mg/kg Post—12.5, 25, 50 mg/kg p.o. | Cisplatin (n = 8) | ↓ | ↑ | - | Total WBC↑ Neutrophil↑ Lymphocyte↑ Stomach injury↑ Small intestine injury↑ |
| Cisplatin + KG (pre-treatment) (n = 8) | ↑ | ↓ | - | Total WBC↓ Neutrophil↓ Lymphocyte↓ Stomach injury↓ Small intestine injury↓ | ||||
| Cisplatin + KG (post-treatment) (n = 8) | ↑ | ↓ | - | - | ||||
| Woo et al. 2016 [43] | Balb/c mice | Single 8 mg/kg, i.p. | Sip-jeon-dea-bo-tang [SJDBT; Angelicae Gigantis Radix, Astragali Radix, Atractylodis Rhizoma Alba, Cinnamomi Cortex, Cnidii Rhizoma, Paeoniae Radix, Ginseng Radix, Poria Sclerotium, Rehmannia Radix, Glycyrrhizae Radix et Rhizoma] (water extract) 678.4 mg/kg, p.o. | Cisplatin (n = 6) | ↓ | - | ↓ | Leptin↓ IL-6↓ |
| Cisplatin + SJDBT (Single) (n = 6) | ↑ | - | - | Leptin↑ IL-6↑ JAK1/STAT3↑ | ||||
| Cisplatin + SJDBT (multiple) (n = 6) | ↑ | - | ↑ | Leptin↑ IL-6↑ | ||||
| Kim et al. 2017 [47] | SD rats | Single 6 mg/kg, i.p. | Rhus verniciflua Stoke [RVX] (water extract) 25, 50, 100 mg/kg, p.o. | Cisplatin (n = 6) | ↓ | ↑ | ↓ | 5-HT↑ 5-HT3A receptor↑ SERT↓ TNF-α↑ IL-6↑ IL-1β↑ WBC↓ Lymphocyte↓ Bone-marrow tissue injury↑ Thymus weight↓ Spleen weight↓ |
| Cisplatin + RVX (n = 6) | ↑ | ↓ | ↑ | 5-HT↓ 5-HT3A receptor↓ SERT↑ TNF-α↓ IL-6↓ IL-1β↓ WBC↑ Lymphocyte↑ Bone-marrow tissue injury↓ Thymus weight↑ Spleen weights↑ | ||||
| Woo et al. 2017 [44] | Balb/c mice | Single 8 mg/kg, i.p. | LCBP-Anocure [LA; Atractylodis Rhizoma Alba (AJ), Angelicae Gigantis Radix (AG), Astragali Radix (AM), Lonicerae Flos (LJ), Taraxaci Herba (TP), Prunellae Spica (PV)]-16001, 16002, 16003 (water extract) 1000 mg/kg, p.o. | Cisplatin (n = 4) | ↓ | - | ↓ | Active ghrelin↓ Leptin↓ IL-6↓ p-JAK1↓p-STAT3↓ WBC↓ Neutrophil↓ |
| Cisplatin + LA16001 (n = 4) | ↑ | - | ↑ | Active ghrelin↑ Leptin↑ IL-6↑ p-JAK1↑ p-STAT3↑ WBC↑ Neutrophil↑ | ||||
| Cisplatin + LA16002 or LA16003 or SJDBT or AJ or AG or LJ or PV (n = 4) | ↑ | - | - | - | ||||
| Cisplatin + MGA (100 mg/kg, p.o.) or AM or TP (n = 4) | ↑ | - | ↑ | - | ||||
| Song et al. 2017 [48] | Wistar rats | Single 5 mg/kg, i.p. | He-Wei granules [HWKL; Pinelliae Tuber, Zingiberis Rhizoma Recens, Ginseng Radix et Rhizoma, Scutellariae Radix, Coptidis Rhizoma, Glycyrrhizae Radix et Rhizoma, Jujubae Fructus] (water extract) 1.18 (low), 2.36 (middle), 4.725 (high) mg/kg, i.g. | Cisplatin (n = 12) | ↓ | ↑ | ↓ | 5-HIAA↓ SP↑ 5-HT↑ 5-HIAA/5-HT↓ SERT↓ MAO-A↓ 5-HT3AR↑ PPTA↑ TPH-1↑ TPH-2↑ NK-1R↑ TPH-1↑ OB↑ GH↓ GH/OB↓ GPR39↑ EGFR↓ pERK1/2↓ GSH-PX↓ Stomach injury↑ SOD↓ MDA↑ Ileum injury↑ |
| Cisplatin + HWKL (n = 12) | ↑ | ↓ | ↑ | 5-HIAA↑ SP↓ 5-HT↓ 5-HIAA/5-HT↑ SERT↑ MAO-A↑ 5-HT3AR↓ PPTA↓ TPH-1↓ TPH-2↓ NK-1R↓ TPH-1↓ OB↓ GH↑ GH/OB↑ GPR39↓ EGFR↑ pERK1/2↑ GSH-PX↑ Stomach injury↓ SOD↑ MDA↓ Ileum injury↓ | ||||
| Kim et al. 2019 [78] | C57BL/6 mice | Multiple (3 times— D1, 6, and 11) 5 mg/kg, i.p. | HemoHIM [Atractylodis Rhizoma Alba, Angelica Gigas Nakai, Astragali Radix] (water extract) 100, 250, 500 mg/kg, p.o. | Cisplatin (n = 7) | - | - | ↓ | NK cell activity↓ Macrophage phagocytotic activity↓CD4+ T lymphocytes↓ Splenocytes↓ IL-2↓ IFN-γ↓ |
| Cisplatin + HemoHIM (n = 7) | - | - | ↑ | NK cell activity↑ Macrophage phagocytotic activity↑CD4+ T lymphocytes↑ Splenocytes↑ IL-2↑ IFN-γ↑ | ||||
| Chen et al. 2019 [79] | C57BL/6 mice | Multiple (9 times— 3 days/week) 5 mg/kg, i.p. | Zhen-Qi Sijunzi [ZQ-SJZ; Ginseng Radix, Atractylodis Rhizoma Alba, Poria Sclerotium, Glycyrrhizae Preparata, Hedysari Radix, Fructus Ligustri Lucidi] (water extract) 700 mg/kg, p.o. | Cisplatin (n = 18) | ↓ | - | ↓ | Intestinal mucosal damage↑ |
| Cisplatin + ZQ-SJZ (n = 18) | ↑ | - | ↑ | Intestinal mucosal damage↓ | ||||
| Goswami et al. 2019 [80] | C57BL/6 mice | Multiple (D0 and 4) 8 mg/kg, i.p. | Ninjin-yoeito [NYT; Angelicae Acutilobae Radix, Atractylodis Rhizoma Alba, Rehmanniae Radix, Poriae Cutis, Ginseng Radix, Citri Unshius Pericarpium Immaturus, Polygalae Radix, Paeoniae Radix, Astragali Radix, Schisandrae Fructus, Glycyrrhizae Radix et Rhizoma] (water extract) 1 g/kg, p.o. | Cisplatin (n = 6) | ↓ | - | ↓ | - |
| Cisplatin + NYT (n = 6) | ↑ | - | ↑ | IR-positive NPY neurons [Ca2+]i ↑ Ghrelin-responsive and unresponsive NPY neurons [Ca2+]i ↑ | ||||
| Pathways | Cisplatin | Herbal Medicines | Measured Locations | ||
|---|---|---|---|---|---|
| 5-HT | 5-HT and 5-HT3A receptor | ↑ | RVX [47] | ↓ | Small intestine |
| HWKL [48] | Ileum, medulla oblongata, serum | ||||
| TPH1 | HWKL [57] | Ileum | |||
| TPH2 | Medulla oblongata | ||||
| SERT | ↓ | RVX [47] | ↑ | Small intestine | |
| HWKL [48] | Medulla oblongata, ileum | ||||
| 5 HIAA | Ileum, serum | ||||
| Cytokine | IL-6 | ↓ | SJDBT [43], LA16001 [44] | ↑ | Fat, serum |
| ↑ | RVX [47] | ↓ | Stomach | ||
| IL-1β | |||||
| TNF-α | |||||
| ↓ | HemoHIM [78] | ↑ | Spleen | ||
| IL-2 | |||||
| IFN-γ | |||||
| IL-4 | ↑ | ↓ | |||
| WBC | Total Number | ↑ | KG [77] | ↓ | Serum |
| ↓ | RVX [47], LA16001 [44] | ↑ | |||
| Lymphocytes | ↑ | KG [77] | ↓ | ||
| ↓ | RVX [47] | ↑ | |||
| HemoHIM [78] | Spleen | ||||
| Neutrophils | ↑ | KG [77] | ↓ | Serum | |
| ↓ | LA16001 [44] | ↑ | |||
| NK cell activity | ↓ | HemoHIM [78] | ↑ | ||
| Macrophage Phagocytotic activity | Peritoneal cavity | ||||
| Splenocyte proliferation | Spleen | ||||
| Hormone | Ghrelin | ↓ | RKT [45], HWKL [48] | ↑ | Serum |
| LA16001 [44] | Stomach | ||||
| HWKL [48] | Antrum | ||||
| GHS-R1a | RKT [45] | Hypothalamus | |||
| Leptin | SJDBT [43] | Fat, serum | |||
| LA16001 [44] | Fat, hypothalamus | ||||
2.3. White Blood Cells (WBCs)
2.4. Ghrelin
2.5. Leptin
2.6. Most Used Herbal Medicines
3. Discussion
4. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dasari, S.; Bernard Tchounwou, P. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S. Cisplatin: The first metal based anticancer drug. Bioorganic Chem. 2019, 88, 102925. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.Y.; Woodward, N.; Coward, J.I.G. Cisplatin versus carboplatin: Comparative review of therapeutic management in solid malignancies. Crit. Rev. Oncol. /Hematol. 2016, 102, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florea, A.-M.; Büsselberg, D. Cisplatin as an Anti-Tumor Drug: Cellular Mechanisms of Activity, Drug Resistance and Induced Side Effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, L.; Ghiselli, S.; Guaran, V.; Chicca, M.; Simoni, E.; Olivetto, E.; Lelli, G.; Martini, A. Correlation of adverse effects of cisplatin administration in patients affected by solid tumours: A retrospective evaluation. Oncol. Rep. 2013, 29, 1285–1292. [Google Scholar] [CrossRef] [Green Version]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- Shahid, F.; Farooqui, Z.; Khan, F. Cisplatin-induced gastrointestinal toxicity: An update on possible mechanisms and on available gastroprotective strategies. Eur. J. Pharmacol. 2018, 827, 49–57. [Google Scholar] [CrossRef]
- Le-Rademacher, J.G.; Crawford, J.; Evans, W.J.; Jatoi, A. Overcoming obstacles in the design of cancer anorexia/weight loss trials. Crit. Rev. Oncol. /Hematol. 2017, 117, 30–37. [Google Scholar] [CrossRef]
- Bearcroft, C.P.; Domizio, P.; Mourad, F.H.; André, E.A.; Farthing, M.J.G. Cisplatin impairs fluid and electrolyte absorption in rat small intestine: A role for 5-hydroxytryptamine. Gut 1999, 44, 174. [Google Scholar] [CrossRef] [Green Version]
- Song, M.Y.; Ku, S.K.; Kim, H.J.; Han, J.S. Low molecular weight fucoidan ameliorating the chronic cisplatin-induced delayed gastrointestinal motility in rats. Food Chem. Toxicol. 2012, 50, 4468–4478. [Google Scholar] [CrossRef]
- Vera, G.; Chiarlone, A.; Martín, M.I.; Abalo, R. Altered feeding behaviour induced by long-term cisplatin in rats. Auton. Neurosci. 2006, 126-127, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Churm, D.; Andrew, I.M.; Holden, K.; Hildreth, A.J.; Hawkins, C. A questionnaire study of the approach to the anorexia-cachexia syndrome in patients with cancer by staff in a district general hospital. Supportive Care Cancer Off. J. Multinatl. Assoc. Supportive Care Cancer 2009, 17, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.P. Gastrointestinal Toxicity of Chemotherapeutic Agents. Semin. Oncol. 2006, 33, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Holland, J.F. Nutritional Consequences of Cancer Chemotherapy and Immunotherapy. Cancer Res. 1977, 37, 2395. [Google Scholar]
- Kokal, W.A. The impact of antitumor therapy on nutrition. Cancer 1985, 55, 273–278. [Google Scholar] [CrossRef]
- Hattori, T.; Yakabi, K.; Takeda, H. Cisplatin-induced anorexia and ghrelin. Vitam. Horm. 2013, 92, 301–317. [Google Scholar]
- Van Cutsem, E.; Arends, J. The causes and consequences of cancer-associated malnutrition. Eur. J. Oncol. Nurs. 2005, 9, S51–S63. [Google Scholar] [CrossRef]
- Sinno, M.H.; Coquerel, Q.; Boukhettala, N.; Coëffier, M.; Gallas, S.; Terashi, M.; Ibrahim, A.; Breuillé, D.; Déchelotte, P.; Fetissov, S.O. Chemotherapy-induced anorexia is accompanied by activation of brain pathways signaling dehydration. Physiol. Behav. 2010, 101, 639–648. [Google Scholar] [CrossRef]
- Jahn, P.; Renz, P.; Stukenkemper, J.; Book, K.; Kuss, O.; Jordan, K.; Horn, I.; Thoke-Colberg, A.; Schmoll, H.-J.; Landenberger, M. Reduction of chemotherapy-induced anorexia, nausea, and emesis through a structured nursing intervention: A cluster-randomized multicenter trial. Supportive Care Cancer 2009. [CrossRef]
- Mantovani, G.; Macciò, A.; Massa, E.; Madeddu, C. Managing cancer-related anorexia/cachexia. Drugs 2001, 61, 499–514. [Google Scholar] [CrossRef]
- Fernández-Lucas, M.; Díaz-Domínguez, M.E.; Ruiz-Roso, G.; Raoch, V.; Teruel-Briones, J.L.; Quereda-Rodríguez-Navarro, C. Anorexia and megestrol acetate: Treatment versus placebo controlled study. Nefrol. (Engl. Ed.) 2014, 34, 416–417. [Google Scholar]
- Aoyagi, T.; Terracina, K.P.; Raza, A.; Matsubara, H.; Takabe, K. Cancer cachexia, mechanism and treatment. World J. Gastrointest Oncol. 2015, 7, 17–29. [Google Scholar] [CrossRef]
- Cuvelier, G.D.; Baker, T.J.; Peddie, E.F.; Casey, L.M.; Lambert, P.J.; Distefano, D.S.; Wardle, M.G.; Mychajlunow, B.A.; Romanick, M.A.; Dix, D.B.; et al. A randomized, double-blind, placebo-controlled clinical trial of megestrol acetate as an appetite stimulant in children with weight loss due to cancer and/or cancer therapy. Pediatric Blood Cancer 2014, 61, 672–679. [Google Scholar] [CrossRef]
- Schacter, L.; Rozencweig, M.; Canetta, R.; Kelley, S.; Nicaise, C.; Smaldone, L. Megestrol acetate: Clinical experience. Cancer Treat. Rev. 1989, 16, 49–63. [Google Scholar] [CrossRef]
- Harrison, M.E.; Norris, M.L.; Robinson, A.; Spettigue, W.; Morrissey, M.; Isserlin, L. Use of cyproheptadine to stimulate appetite and body weight gain: A systematic review. Appetite 2019, 137, 62–72. [Google Scholar] [CrossRef]
- Najib, K.; Moghtaderi, M.; Karamizadeh, Z.; Fallahzadeh, E. Beneficial effect of cyproheptadine on body mass index in undernourished children: A randomized controlled trial. Iran J. Pediatr. 2014, 24, 753–758. [Google Scholar] [PubMed]
- Ruiz Garcia, V.; López-Briz, E.; Carbonell Sanchis, R.; Gonzalvez Perales, J.L.; Bort-Marti, S. Megestrol acetate for treatment of anorexia-cachexia syndrome. Cochrane Database Syst. Rev. 2013, 2013. [Google Scholar] [CrossRef]
- Pascual López, A.; Roqué i Figuls, M.; Urrútia Cuchi, G.; Graciela Berenstein, E.; Almenar Pasies, B.; Balcells Alegre, M.; Herdman, M. Systematic review of megestrol acetate in the treatment of anorexia-cachexia syndrome. J. Pain Symptom Manag. 2004, 27, 360–369. [Google Scholar] [CrossRef]
- Krasaelap, A.; Madani, S. Cyproheptadine: A Potentially Effective Treatment for Functional Gastrointestinal Disorders in Children. Pediatric Ann. 2017, 46, e120–e125. [Google Scholar] [CrossRef] [Green Version]
- DiSaia, P.J.; Morrow, C.P. Unusual side effect of megestrol acetate. Am. J. Obstet. Gynecol. 1977, 129, 460–461. [Google Scholar] [CrossRef]
- Nelson, K.A.; Walsh, D.; Hussein, M. A phase II study of low-dose megestrol acetate using twice-daily dosing for anorexia in nonhormonally dependent cancer. Am. J. Hosp. Palliat. Care 2002, 19, 206–210. [Google Scholar] [CrossRef]
- von Mühlendahl, K.E.; Krienke, E.G. Toxicity of cyproheptadine. Side effects and accidental overdosage (author's transl). Mon. Fur Kinderheilkd. 1978, 126, 123–126. [Google Scholar]
- Wortsman, J.; Soler, N.G.; Hirschowitz, J. Side effects of cyproheptadine. Br. Med. J. 1978, 1, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoa, T.P.; Duong, T.B.T.; Thi, H.D.N.; Anna, M.P.; Ali, H.E.; Gianfranco, P. Herbal Medicine for Slowing Aging and Aging-associated Conditions: Efficacy, Mechanisms and Safety. Curr. Vasc. Pharmacol. 2020, 18, 369–393. [Google Scholar]
- Ardalan, M.-R.; Rafieian-Kopaei, M. Is the safety of herbal medicines for kidneys under question? J. Nephropharmacol. 2013, 2, 11–12. [Google Scholar] [PubMed]
- Langmead, L.; Rampton, D.S. Review article: Herbal treatment in gastrointestinal and liver disease--benefits and dangers. Aliment. Pharmacol. Ther. 2001, 15, 1239–1252. [Google Scholar] [CrossRef]
- Bahmani, M.; Zargaran, A.; Rafieian-Kopaei, M. Identification of medicinal plants of Urmia for treatment of gastrointestinal disorders. Rev. Bras. De Farmacogn. 2014, 24, 468–480. [Google Scholar] [CrossRef] [Green Version]
- Anheyer, D.; Frawley, J.; Koch, A.K.; Lauche, R.; Langhorst, J.; Dobos, G.; Cramer, H. Herbal Medicines for Gastrointestinal Disorders in Children and Adolescents: A Systematic Review. Pediatrics 2017, 139, e20170062. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Chen, J.; Xu, L.; Shi, X.; Zhou, X.; An, R. A Network Pharmacology Approach to Uncover the Molecular Mechanisms of Herbal Formula Ban-Xia-Xie-Xin-Tang. Evid.-Based Complementary Altern. Med. 2018, 2018, 4050714. [Google Scholar] [CrossRef]
- Jong, M.-S.; Hwang, S.-J.; Chen, Y.-C.; Chen, T.-J.; Chen, F.-J.; Chen, F.-P. Prescriptions of Chinese Herbal Medicine for Constipation Under the National Health Insurance in Taiwan. J. Chin. Med. Assoc. 2010, 73. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.-L.; Shibata, C.; Naito, H.; Ueno, T.; Funayama, Y.; Fukushima, K.; Matsuno, S.; Sasakimd, I. Intraduodenal and Intrajejunal Administration of the Herbal Medicine, Dai-Kenchu-Tou, Stimulates Small Intestinal Motility via Cholinergic Receptors in Conscious Dogs. Dig. Dis. Sci. 2001, 46, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Hayakawa, T.; Kase, Y.; Ishige, A.; Sasaki, H.; Nishikawa, S.; Kurosawa, S.; Yakabi, K.; Nakamura, T. Mechanisms for Contractile Effect of Dai-kenchu-to in Isolated Guinea Pig Ileum. Dig. Dis. Sci. 2001, 46, 250–256. [Google Scholar] [CrossRef]
- Woo, S.M.; Choi, Y.K.; Kim, A.J.; Yun, Y.J.; Shin, Y.C.; Cho, S.G.; Ko, S.G. Sip-jeon-dea-bo-tang, a traditional herbal medicine, ameliorates cisplatin-induced anorexia via the activation of JAK1/STAT3-mediated leptin and IL-6 production in the fat tissue of mice. Mol. Med. Rep. 2016, 13, 2967–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, S.M.; Lee, K.M.; Lee, G.R.; Park, J.Y.; Lee, H.J.; Bahn, H.J.; Yoon, H.S.; Kim, J.Y.; Shin, Y.C.; Cho, S.G.; et al. Novel herbal medicine LA16001 ameliorates cisplatin-induced anorexia. Mol. Med. Rep. 2018, 17, 2665–2672. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Sadakane, C.; Hattori, T.; Katsurada, T.; Ohkawara, T.; Nagai, K.; Asaka, M. Rikkunshito, an herbal medicine, suppresses cisplatin-induced anorexia in rats via 5-HT2 receptor antagonism. Gastroenterology 2008, 134, 2004–2013. [Google Scholar] [CrossRef]
- Yakabi, K.; Kurosawa, S.; Tamai, M.; Yuzurihara, M.; Nahata, M.; Ohno, S.; Ro, S.; Kato, S.; Aoyama, T.; Sakurada, T.; et al. Rikkunshito and 5-HT2C receptor antagonist improve cisplatin-induced anorexia via hypothalamic ghrelin interaction. Regul. Pept. 2010, 161, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, H.G.; Im, H.J.; Lee, J.S.; Lee, S.B.; Kim, W.Y.; Lee, H.W.; Lee, S.K.; Byun, C.K.; Son, C.G. Antiemetic and Myeloprotective Effects of Rhus verniciflua Stoke in a Cisplatin-Induced Rat Model. Evid.-Based Complementary Altern. Med. Ecam 2017, 2017, 9830342. [Google Scholar]
- Song, Z.H.; Chang, H.; Han, N.; Liu, Z.H.; Wang, Z.L.; Gao, H.; Yin, J. He-Wei granules inhibit chemotherapy-induced vomiting (CINV) in rats by reducing oxidative stress and regulating 5-HT, substance P, ghrelin and obestatin. Rsc. Adv. 2017, 7, 43866–43878. [Google Scholar] [CrossRef] [Green Version]
- Hasler, W.L. Serotonin and the GI tract. Curr. Gastroenterol. Rep. 2009, 11, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Haleem, D.J. Serotonin neurotransmission in anorexia nervosa. Behav. Pharmacol. 2012, 23, 478–495. [Google Scholar] [CrossRef]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The diverse metabolic roles of peripheral serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef]
- Simansky, K.J. Serotonergic control of the organization of feeding and satiety. Behav. Brain Res. 1995, 73, 37–42. [Google Scholar] [CrossRef]
- Jason, C.G.H.; Joanne, A.H.; Clare, L.L.; John, E.B. Serotonin (5-HT) Drugs: Effects on Appetite Expression and Use for the Treatment of Obesity. Curr. Drug Targets 2005, 6, 201–213. [Google Scholar]
- Samanin, R.; Mennini, T.; Ferraris, A.; Bendotti, C.; Borsini, F.; Garattini, S. m-Chlorophenylpiperazine: A central serotonin agonist causing powerful anorexia in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1979, 308, 159–163. [Google Scholar] [CrossRef]
- Gershon, M. Serotonin receptors and transporters—roles in normal and abnormal gastrointestinal motility. Aliment. Pharmacol. Ther. 2004, 20, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Yang, E.; Kim, S.; Park, C.-S.; Park, Y.S.; Jin, Y.-H. Parapheromones suppress chemotherapy side effects. J. Pharmacol. Exp. Ther. 2018, 367, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Jayamohananan, H.; Manoj Kumar, M.K.; T P, A. 5-HIAA as a Potential Biological Marker for Neurological and Psychiatric Disorders. Adv. Pharm. Bull. 2019, 9, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenfors, C.; Ross, S.B. Changes in extracellular 5-HIAA concentrations as measured by in vivo microdialysis technique in relation to changes in 5-HT release. Psychopharmacology 2004, 172, 119–128. [Google Scholar] [CrossRef]
- Asakawa, A.; Inui, A.; Kaga, T.; Katsuura, G.; Fujimiya, M.; Fujino, M.A.; Kasuga, M. Antagonism of ghrelin receptor reduces food intake and body weight gain in mice. Gut 2003, 52, 947. [Google Scholar] [CrossRef] [Green Version]
- Karagiannides, I.; Torres, D.; Tseng, Y.-H.; Bowe, C.; Carvalho, E.; Espinoza, D.; Pothoulakis, C.; Kokkotou, E. Substance P as a novel anti-obesity target. Gastroenterology 2008, 134, 747–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Asano, K.; Tasaka, A.; Ogura, Y.; Kim, S.; Ito, Y.; Yamatodani, A. Involvement of substance P in the development of cisplatin-induced acute and delayed pica in rats. Br. J. Pharmacol. 2014, 171, 2888–2899. [Google Scholar] [CrossRef] [Green Version]
- Rojas, C.; Li, Y.; Zhang, J.; Stathis, M.; Alt, J.; Thomas, A.G.; Cantoreggi, S.; Sebastiani, S.; Pietra, C.; Slusher, B.S. The antiemetic 5-HT3 receptor antagonist Palonosetron inhibits substance P-mediated responses in vitro and in vivo. J. Pharmacol. Exp. Ther. 2010, 335, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Gautron, L.; Layé, S. Neurobiology of inflammation-associated anorexia. Front Neurosci 2010, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Goehler, L.E.; Busch, C.R.; Tartaglia, N.; Relton, J.; Sisk, D.; Maier, S.F.; Watkins, L.R. Blockade of cytokine induced conditioned taste aversion by subdiaphragmatic vagotomy: Further evidence for vagal mediation of immune-brain communication. Neurosci. Lett. 1995, 185, 163–166. [Google Scholar] [CrossRef]
- Solmi, M.; Veronese, N.; Favaro, A.; Santonastaso, P.; Manzato, E.; Sergi, G.; Correll, C.U. Inflammatory cytokines and anorexia nervosa: A meta-analysis of cross-sectional and longitudinal studies. Psychoneuroendocrinology 2015, 51, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Langhans, W.; Savoldelli, D.; Weingarten, S. Comparison of the feeding responses to bacterial lipopolysaccharide and interleukin-1β. Physiol. Behav. 1993, 53, 643–649. [Google Scholar] [CrossRef]
- Bretdibat, J.L.; Bluthe, R.M.; Kent, S.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide and Interleukin-1 Depress Food-Motivated Behavior in Mice by a Vagal-Mediated Mechanism. Brain Behav. Immun. 1995, 9, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layé, S.; Gheusi, G.; Cremona, S.; Combe, C.; Kelley, K.; Dantzer, R.; Parnet, P. Endogenous brain IL-1 mediates LPS-induced anorexia and hypothalamic cytokine expression. Am. J. Physiology. Regul. Integr. Comp. Physiol. 2000, 279, R93–R98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, M.H.; Arnold, M.; Langhans, W. TNF-α tolerance blocks LPS-induced hypophagia but LPS tolerance fails to prevent TNF-α-induced hypophagia. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1998, 274, R741–R745. [Google Scholar] [CrossRef]
- Porter, M.H.; Hrupka, B.J.; Altreuther, G.; Arnold, M.; Langhans, W. Inhibition of TNF-α production contributes to the attenuation of LPS-induced hypophagia by pentoxifylline. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2000, 279, R2113–R2120. [Google Scholar] [CrossRef]
- Bodnar, R.J.; Pasternak, G.W.; Mann, P.E.; Paul, D.; Warren, R.; Donner, D.B. Mediation of Anorexia by Human Recombinant Tumor Necrosis Factor through a Peripheral Action in the Rat. Cancer Res. 1989, 49, 6280. [Google Scholar] [PubMed]
- Fattori, E.; Cappelletti, M.; Costa, P.; Sellitto, C.; Cantoni, L.; Carelli, M.; Faggioni, R.; Fantuzzi, G.; Ghezzi, P.; Poli, V. Defective inflammatory response in interleukin 6-deficient mice. J. Exp. Med. 1994, 180, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.M.; Vogt, S.K.; Burney, M.W.; Muglia, L.J. COX-2 inhibition attenuates anorexia during systemic inflammation without impairing cytokine production. Am. J. Physiol.-Endocrinol. Metab. 2002, 282, E650–E656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, T.P.; Marks, D.L. Pathophysiology and treatment of inflammatory anorexia in chronic disease. J. Cachexia Sarcopenia Muscle 2010, 1, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, H.H.; Dey, L.; Mehendale, S.; Xie, J.T.; Wu, J.A.; Yuan, C.S. Scutellaria baicalensis extract decreases cisplatin-induced pica in rats. Cancer Chemother. Pharmacol. 2003, 52, 453–458. [Google Scholar] [CrossRef]
- Mehendale, S.; Aung, H.; Wang, A.; Yin, J.J.; Wang, C.Z.; Xie, J.T.; Yuan, C.S. American ginseng berry extract and ginsenoside Re attenuate cisplatin-induced kaolin intake in rats. Cancer Chemother. Pharmacol. 2005, 56, 63–69. [Google Scholar] [CrossRef]
- Raghavendran, H.R.; Rekha, S.; Shin, J.W.; Kim, H.G.; Wang, J.H.; Park, H.J.; Choi, M.K.; Cho, J.H.; Son, C.G. Effects of Korean ginseng root extract on cisplatin-induced emesis in a rat-pica model. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 215–221. [Google Scholar] [CrossRef]
- Kim, S.K.; Kwon, D.A.; Lee, H.S.; Kim, H.K.; Kim, W.K. Preventive Effect of the Herbal Preparation, HemoHIM, on Cisplatin-Induced Immune Suppression. Evid.-Based Complementary Altern. Med. Ecam 2019, 2019, 3494806. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Yang, T.T.; Cheng, T.S.; Hsiao, T.F.; Chang, P.M.H.; Leu, J.Y.; Wang, F.S.; Hsu, S.L.; Huang, C.Y.F.; Lai, J.M. Modified Sijunzi decoction can alleviate cisplatin-induced toxicity and prolong the survival time of cachectic mice by recovering muscle atrophy. J. Ethnopharmacol. 2019, 233, 47–55. [Google Scholar] [CrossRef]
- Goswami, C.; Dezaki, K.; Wang, L.; Inui, A.; Seino, Y.; Yada, T. Ninjin-yoeito activates ghrelin-responsive and unresponsive NPY neurons in the arcuate nucleus and counteracts cisplatin-induced anorexia. Neuropeptides 2019, 75, 58–64. [Google Scholar] [CrossRef]
- De Filippo, E.; Marra, M.; Alfinito, F.; Di Guglielmo, M.L.; Majorano, P.; Cerciello, G.; De Caprio, C.; Contaldo, F.; Pasanisi, F. Hematological complications in anorexia nervosa. Eur. J. Clin. Nutr. 2016, 70, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Hütter, G.; Ganepola, S.; Hofmann, W.-K. The hematology of anorexia nervosa. Int. J. Eat. Disord. 2009, 42, 293–300. [Google Scholar] [CrossRef]
- Janssen, E.M.; Lemmens, E.E.; Wolfe, T.; Christen, U.; von Herrath, M.G.; Schoenberger, S.P. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 2003, 421, 852–856. [Google Scholar] [CrossRef]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.; Bowers, C.Y.; Broglio, F. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Cummings, D.E.; Shannon, M.H. Roles for Ghrelin in the Regulation of Appetite and Body Weight. Arch. Surg. 2003, 138, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Wren, A.M.; Small, C.J.; Ward, H.L.; Murphy, K.G.; Dakin, C.L.; Taheri, S.; Kennedy, A.R.; Roberts, G.H.; Morgan, D.G.A.; Ghatei, M.A.; et al. The Novel Hypothalamic Peptide Ghrelin Stimulates Food Intake and Growth Hormone Secretion. Endocrinology 2000, 141, 4325–4328. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Ghrelin and des-acyl ghrelin: Two major forms of rat ghrelin peptide in gastrointestinal tissue. Biochem. Biophys. Res. Commun. 2000, 279, 909–913. [Google Scholar] [CrossRef]
- Tschöp, M.; Smiley, D.L.; Heiman, M.L. Ghrelin induces adiposity in rodents. Nature 2000, 407, 908–913. [Google Scholar] [CrossRef]
- Asakawa, A.; Inui, A.; Kaga, T.; Yuzuriha, H.; Nagata, T.; Ueno, N.; Makino, S.; Fujimiya, M.; Niijima, A.; Fujino, M.A.; et al. Ghrelin is an appetite-stimulatory signal from stomach with structural resemblance to motilin. Gastroenterology 2001, 120, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, M.; Murakami, N.; Date, Y.; Kojima, M.; Matsuo, H.; Kangawa, K.; Matsukura, S. A role for ghrelin in the central regulation of feeding. Nature 2001, 409, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Shinomiya, T.; Fukunaga, M.; Akamizu, T.; Irako, T.; Yokode, M.; Kangawa, K.; Nakai, Y.; Nakai, Y. Plasma acylated ghrelin levels correlate with subjective symptoms of functional dyspepsia in female patients. Scand. J. Gastroenterol. 2005, 40, 648–653. [Google Scholar] [CrossRef]
- Isomoto, H.; Ueno, H.; Nishi, Y.; Yasutake, T.; Tanaka, K.; Kawano, N.; Ohnita, K.; Mizuta, Y.; Inoue, K.; Nakazato, M.; et al. Circulating Ghrelin Levels in Patients with Various Upper Gastrointestinal Diseases. Dig. Dis. Sci. 2005, 50, 833–838. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Kohno, D.; Nakata, M.; Maekawa, F.; Fujiwara, K.; Maejima, Y.; Kuramochi, M.; Shimazaki, T.; Okano, H.; Onaka, T.; Yada, T. Leptin suppresses ghrelin-induced activation of neuropeptide Y neurons in the arcuate nucleus via phosphatidylinositol 3-kinase-and phosphodiesterase 3-mediated pathway. Endocrinology 2007, 148, 2251–2263. [Google Scholar] [CrossRef]
- Gropp, E.; Shanabrough, M.; Borok, E.; Xu, A.W.; Janoschek, R.; Buch, T.; Plum, L.; Balthasar, N.; Hampel, B.; Waisman, A. Agouti-related peptide–expressing neurons are mandatory for feeding. Nat. Neurosci. 2005, 8, 1289–1291. [Google Scholar] [CrossRef]
- Luquet, S.; Perez, F.A.; Hnasko, T.S.; Palmiter, R.D. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science 2005, 310, 683–685. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Hebebrand, J.; Muller, T.; Holtkamp, K.; Herpertz-Dahlmann, B. The role of leptin in anorexia nervosa: Clinical implications. Mol. Psychiatry 2007, 12, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, L.S. Cancer Drug Resistance Research Perspectives; Nova Publishers: New York, NY, USA, 2007. [Google Scholar]
- Garcia, J.M.; Scherer, T.; Chen, J.-a.; Guillory, B.; Nassif, A.; Papusha, V.; Smiechowska, J.; Asnicar, M.; Buettner, C.; Smith, R.G. Inhibition of cisplatin-induced lipid catabolism and weight loss by ghrelin in male mice. Endocrinology 2013, 154, 3118–3129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakabi, K.; Sadakane, C.; Noguchi, M.; Ohno, S.; Ro, S.; Chinen, K.; Aoyama, T.; Sakurada, T.; Takabayashi, H.; Hattori, T. Reduced Ghrelin Secretion in the Hypothalamus of Rats due to Cisplatin-Induced Anorexia. Endocrinology 2010, 151, 3773–3782. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-T.; Ko, J.-L.; Liu, T.-C.; Chao, P.-T.; Ou, C.-C. Protective effect of D-methionine on body weight loss, anorexia, and nephrotoxicity in cisplatin-induced chronic toxicity in rats. Integr. Cancer Ther. 2018, 17, 813–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- de Matos Feijó, F.; Bertoluci, M.C.; Reis, C. Serotonin and hypothalamic control of hunger: A review. Rev. Da Assoc. Médica Bras. (Engl. Ed.) 2011, 57, 74–77. [Google Scholar] [CrossRef]
- Halford, J.C.; Blundell, J.E. Separate systems for serotonin and leptin in appetite control. Ann. Med. 2000, 32, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Barnes, N.M.; Ge, J.; Jones, W.G.; Naylor, R.J.; Rudd, J.A. Cisplatin induced emesis: Preliminary results indicative of changes in plasma levels of 5-hydroxytryptamine. Br. J. Cancer 1990, 62, 862–864. [Google Scholar] [CrossRef] [Green Version]
- Cubeddu, L.X.; Hoffmann, I.S.; Fuenmayor, N.T.; Finn, A.L. Efficacy of Ondansetron (Gr 38032F) and the Role of Serotonin in Cisplatin-Induced Nausea and Vomiting. N. Engl. J. Med. 1990, 322, 810–816. [Google Scholar] [CrossRef]
- Wilder-Smith, O.H.G.; Borgeat, A.; Chappuis, P.; Fathi, M.; Forni, M. Urinary serotonin metabolite excretion during cisplatin chemotherapy. Cancer 1993, 72, 2239–2241. [Google Scholar] [CrossRef]
- Schwörer, H.; Racké, K.; Kilbinger, H. Cisplatin increases the release of 5-hydroxytryptamine (5-HT) from the isolated vascularly perfused small intestine of the guinea-pig: Involvement of 5-HT3 receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1991, 344, 143–149. [Google Scholar] [CrossRef]
- Kamato, T.; Ito, H.; Nagakura, Y.; Nishida, A.; Yuki, H.; Yamano, M.; Miyata, K. Mechanisms of cisplatin- and m-chlorophenylbiguinide-induced emesis in ferrets. Eur. J. Pharmacol. 1993, 238, 369–376. [Google Scholar] [CrossRef]
- Murotani, T.; Ishizuka, T.; Isogawa, Y.; Karashima, M.; Yamatodani, A. Possible involvement of serotonin 5-HT2 receptor in the regulation of feeding behavior through the histaminergic system. Neuropharmacology 2011, 61, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Banas, S.M.; Doly, S.; Boutourlinsky, K.; Diaz, S.L.; Belmer, A.; Callebert, J.; Collet, C.; Launay, J.-M.; Maroteaux, L. Deconstructing Antiobesity Compound Action: Requirement of Serotonin 5-HT2B Receptors for Dexfenfluramine Anorectic Effects. Neuropsychopharmacology 2011, 36, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Abdallah, L.; Goulding, E.H.; Bonasera, S.J.; Tecott, L.H. Hyperactivity and Reduced Energy Cost of Physical Activity in Serotonin 5-HT2C Receptor Mutant Mice. Diabetes 2003, 52, 315. [Google Scholar]
- Nonogaki, K.; Strack, A.M.; Dallman, M.F.; Tecott, L.H. Leptin-independent hyperphagia and type 2 diabetes in mice with a mutated serotonin 5-HT 2C receptor gene. Nat. Med. 1998, 4, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.; Cunningham, K.; Tyers, M. Selectivity of 5-HT3 receptor antagonists and anti-emetic mechanisms of action. Anti-Cancer Drugs 1992, 3, 79–85. [Google Scholar] [CrossRef]
- Hesketh, P.J. Clinical science review: Comparative review of 5-HT3 receptor antagonists in the treatment of acute chemotherapy-induced nausea and vomiting. Cancer Investig. 2000, 18, 163–173. [Google Scholar] [CrossRef]
- Raymond, N.C.; Dysken, M.; Bettin, K.; Eckert, E.D.; Crow, S.J.; Markus, K.; Pomeroy, C. Cytokine production in patients with anorexia nervosa, bulimia nervosa, and obesity. Int. J. Eat. Disord. 2000, 28, 293–302. [Google Scholar] [CrossRef]
- Kim, J.H.; Yi, Y.-S.; Kim, M.-Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Yoo, S.R.; Park, H.J.; Lee, N.H.; Shin, J.W.; Sathyanath, R.; Cho, J.H.; Son, C.G. Antioxidant effects of Panax ginseng C.A. Meyer in healthy subjects: A randomized, placebo-controlled clinical trial. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 2229–2235. [Google Scholar] [CrossRef]
- Kiefer, D.; Pantuso, T. Panax ginseng. Am. Fam. Physician 2003, 68, 1539–1542. [Google Scholar]
- Ahn, J.-Y.; Choi, I.-S.; Shim, J.-Y.; Yun, E.-K.; Yun, Y.-S.; Jeong, G.; Song, J.-Y. The immunomodulator ginsan induces resistance to experimental sepsis by inhibiting Toll-like receptor-mediated inflammatory signals. Eur. J. Immunol. 2006, 36, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Keum, Y.S.; Han, S.S.; Chun, K.S.; Park, K.K.; Park, J.H.; Lee, S.K.; Surh, Y.J. Inhibitory effects of the ginsenoside Rg3 on phorbol ester-induced cyclooxygenase-2 expression, NF-kappaB activation and tumor promotion. Mutat. Res. 2003, 523-524, 75–85. [Google Scholar] [CrossRef]
- Ok, S.; Oh, S.R.; Jung, T.S.; Jeon, S.O.; Jung, J.W.; Ryu, D.S. Effects of Angelica gigas Nakai as an Anti-Inflammatory Agent in In Vitro and In Vivo Atopic Dermatitis Models. Evid.-Based Complementary Altern. Med. Ecam 2018, 2018, 2450712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-G.; Jan, Y.-S.; Tsai, P.-W.; Norimoto, H.; Michihara, S.; Murayama, C.; Wang, C.-C. Anti-inflammatory and Antinociceptive Constituents of Atractylodes japonica Koidzumi. J. Agric. Food Chem. 2016, 64, 2254–2262. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.E.; Kim, J.N.; Kwon, M.J.; Lee, J.R.; Kim, S.C.; Nam, J.H.; Kim, B.J. The traditional medicine bojungikki-tang increases intestinal motility. Pharmacogn. Mag. 2021, 17, 1. [Google Scholar]
- Lee, M.-Y.; Shin, I.-S.; Jeon, W.-Y.; Seo, C.-S.; Ha, H.; Huh, J.-I.; Shin, H.-K. Protective effect of Bojungikki-tang, a traditional herbal formula, against alcohol-induced gastric injury in rats. J. Ethnopharmacol. 2012, 142, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.-p.; Chen, F.-j.; Jong, M.-s.; Tsai, H.-l.; Wang, J.-r.; Hwang, S.-j. Modern use of Chinese herbal formulae fromShang-Han Lun. Chin. Med. J. 2009, 122, 1889–1894. [Google Scholar]
- Murata, P.; Kase, Y.; Ishige, A.; Sasaki, H.; Kurosawa, S.; Nakamura, T. The herbal medicine Dai-kenchu-to and one of its active components [6]-shogaol increase intestinal blood flow in rats. Life Sci. 2002, 70, 2061–2070. [Google Scholar] [CrossRef] [Green Version]
- Shibata, C.; Sasaki, I.; Naito, H.; Ueno, T.; Matsuno, S. The herbal medicine Dai-Kenchu-Tou stimulates upper gut motility through cholinergic and 5-hydroxytryptamine 3 receptors in conscious dogs. Surgery 1999, 126, 918–924. [Google Scholar] [CrossRef]
- Kawasaki, N.; Nakada, K.; Suzuki, Y.; Furukawa, Y.; Hanyu, N.; Kashiwagi, H. Effect of Dai-kenchu-to on gastrointestinal motility and gastric emptying. Int. J. Surg. 2009, 7, 218–222. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, K.; Kurita, N.; Higashijima, J.; Miyatani, T.; Miyamoto, H.; Nishioka, M.; Shimada, M. Kampo medicine “Dai-kenchu-to” prevents bacterial translocation in rats. Dig. Dis. Sci. 2008, 53, 1824–1831. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, D.; Kim, B.; Ko, S.-G.; Kim, W. Effect and Mechanism of Herbal Medicines on Cisplatin-Induced Anorexia. Pharmaceuticals 2022, 15, 208. https://doi.org/10.3390/ph15020208
Min D, Kim B, Ko S-G, Kim W. Effect and Mechanism of Herbal Medicines on Cisplatin-Induced Anorexia. Pharmaceuticals. 2022; 15(2):208. https://doi.org/10.3390/ph15020208
Chicago/Turabian StyleMin, Daeun, Bonglee Kim, Seong-Gyu Ko, and Woojin Kim. 2022. "Effect and Mechanism of Herbal Medicines on Cisplatin-Induced Anorexia" Pharmaceuticals 15, no. 2: 208. https://doi.org/10.3390/ph15020208
APA StyleMin, D., Kim, B., Ko, S. -G., & Kim, W. (2022). Effect and Mechanism of Herbal Medicines on Cisplatin-Induced Anorexia. Pharmaceuticals, 15(2), 208. https://doi.org/10.3390/ph15020208