
From the North Sea to Drug Repurposing, the Antiseizure Activity of Halimide and Plinabulin

 , , , ,
, , , ,  ,
,
Abstract
:
1. Introduction
2. Results
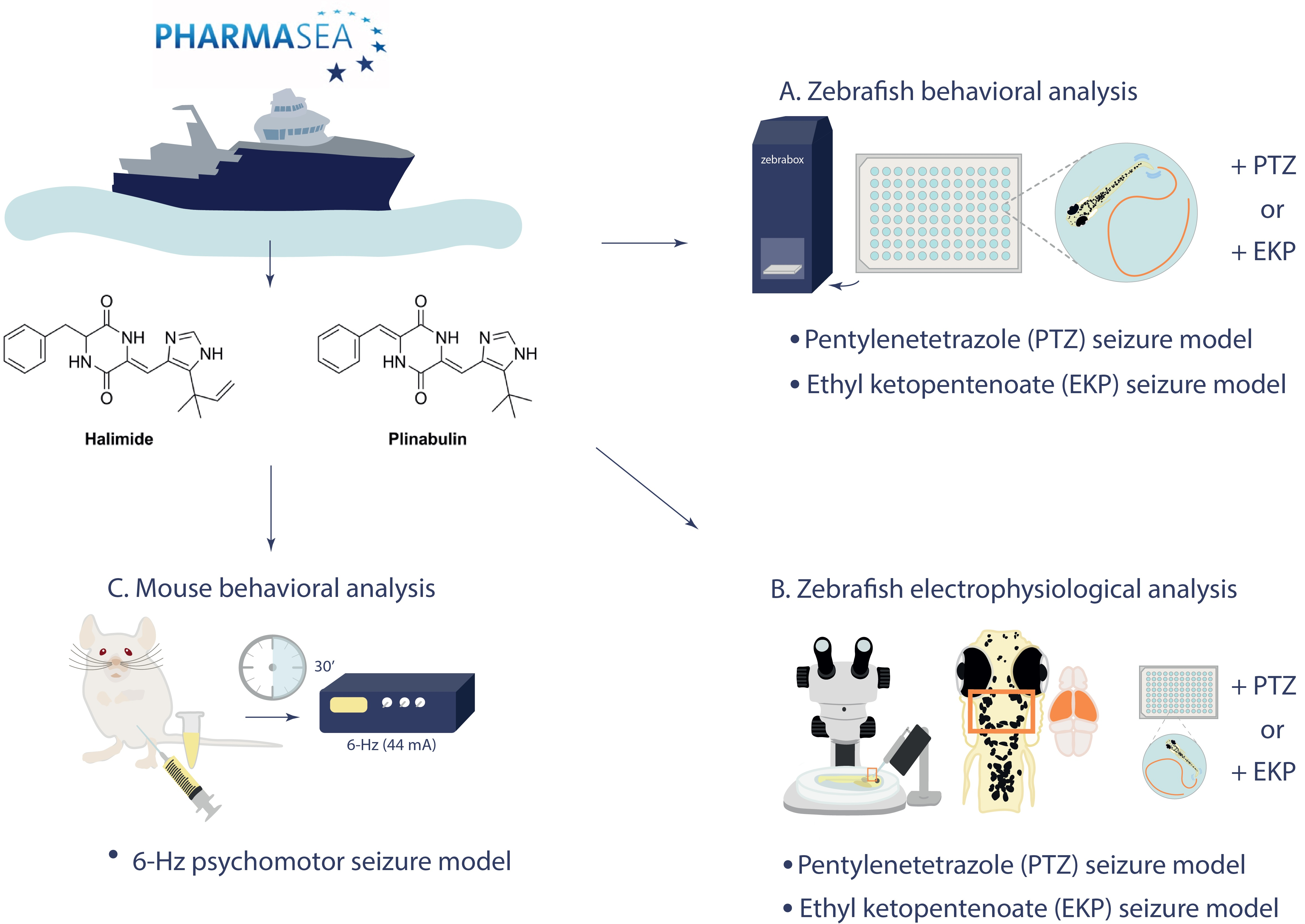
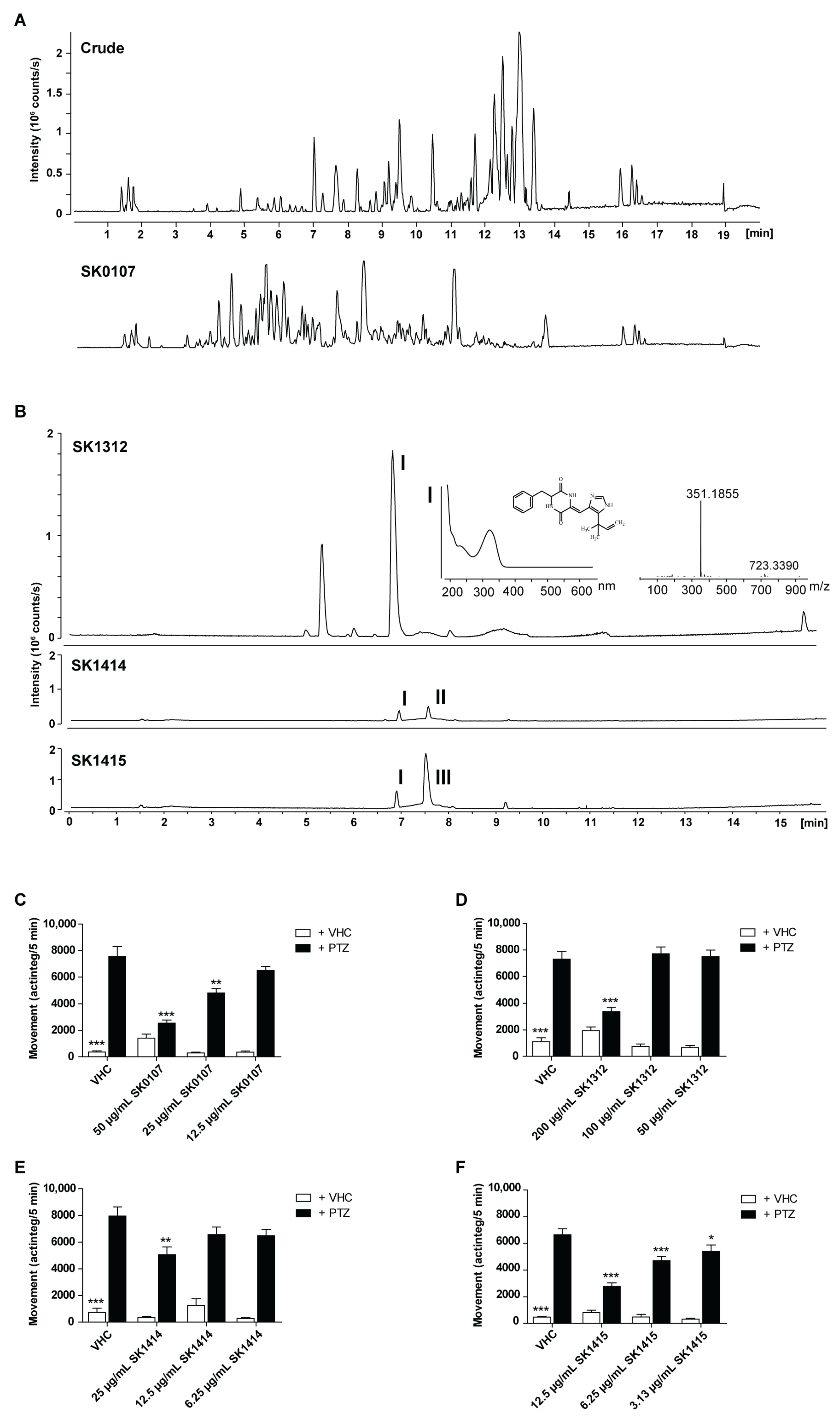
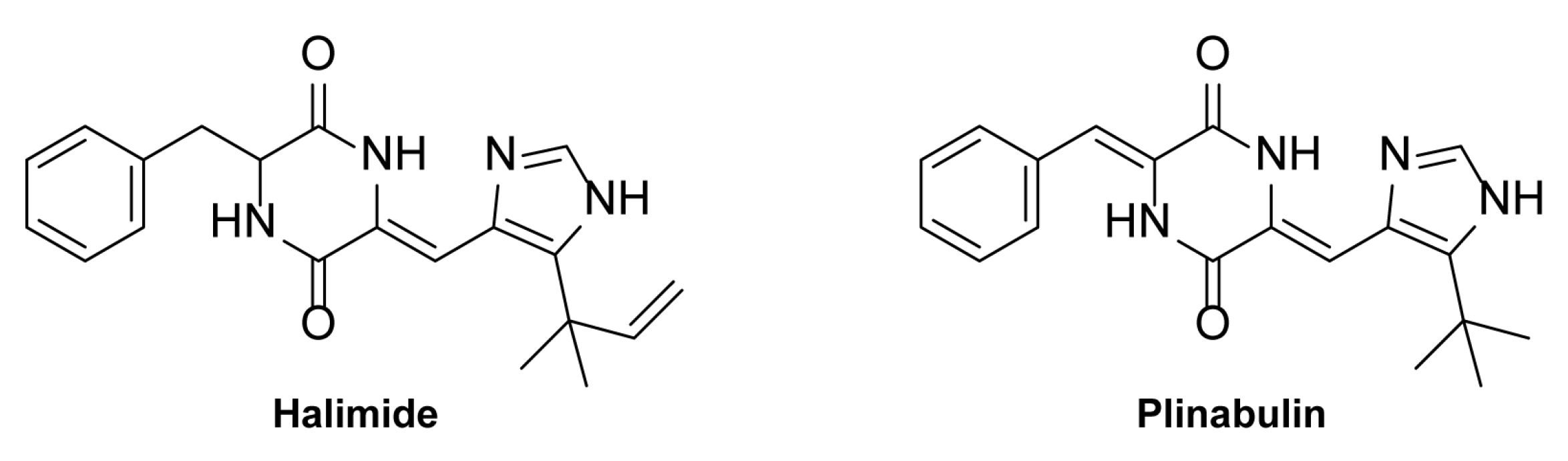
2.1. Marine Antiseizure Drug Discovery and Bioactivity-Guided Purification of Halimide
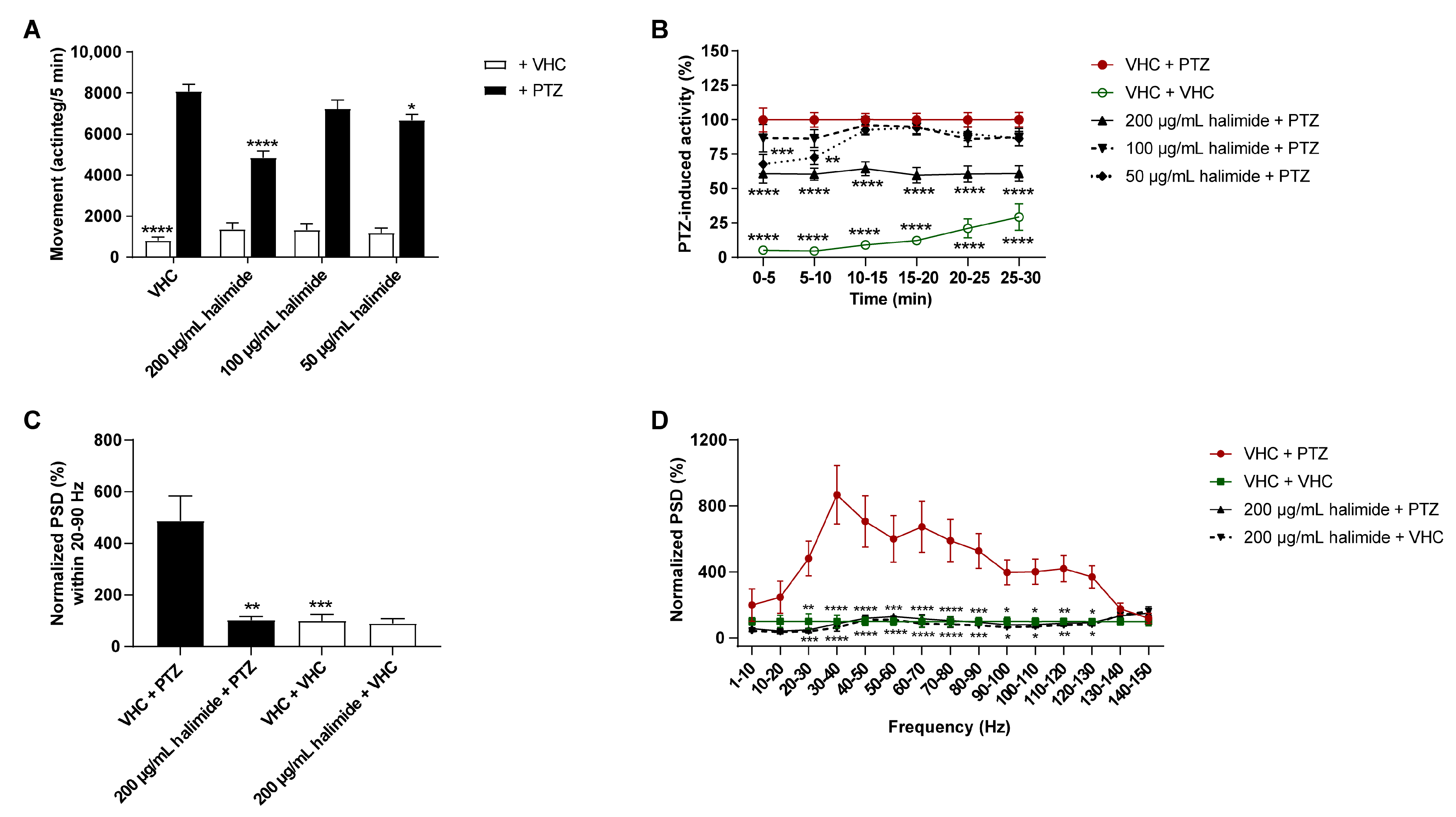
2.2. Halimide Ameliorates Seizures and Epileptiform Brain Activity in the Zebrafish PTZ Seizure Model
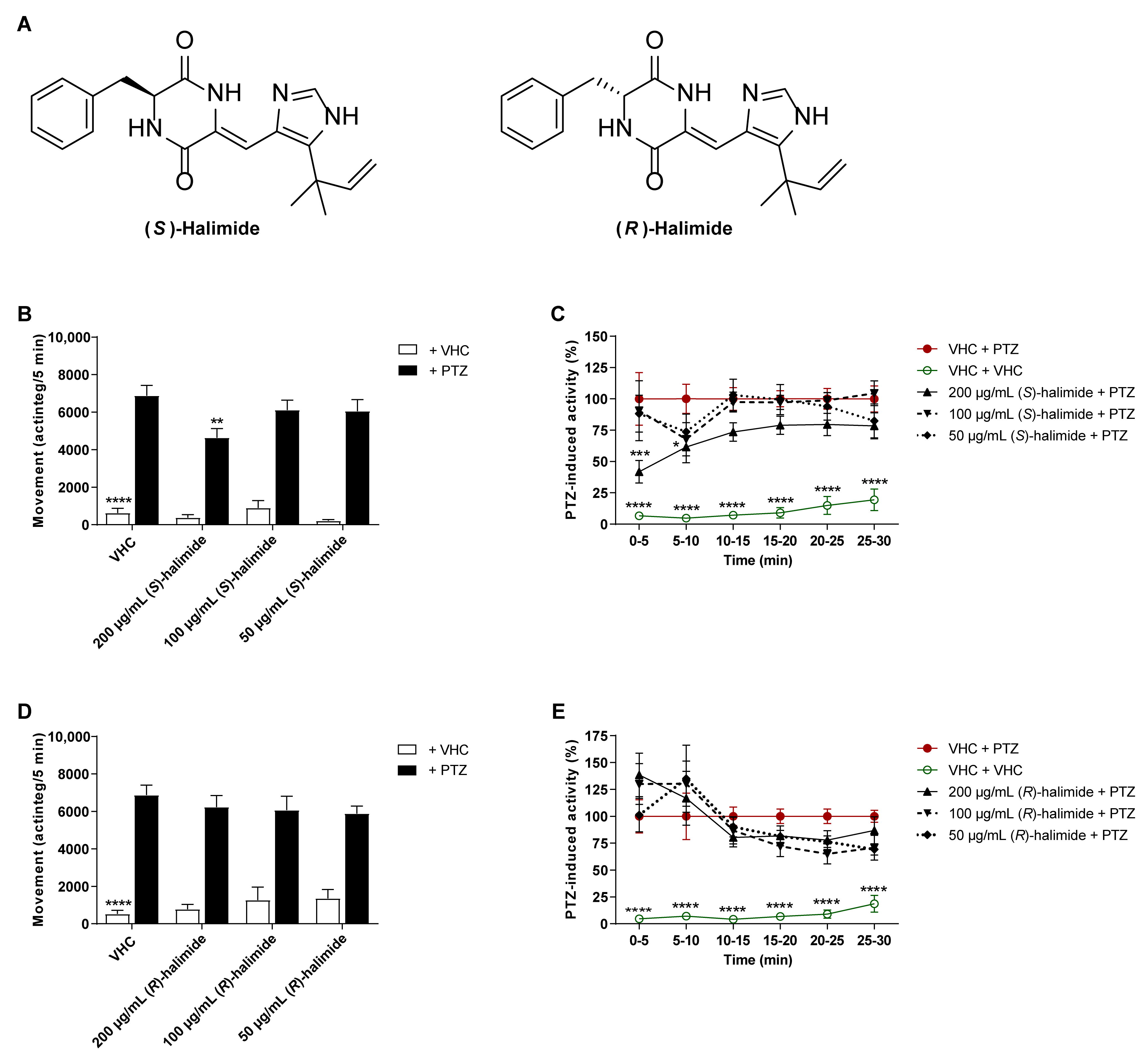
2.3. The S-Enantiomer of Halimide Is Active within the Scalemic Mixture
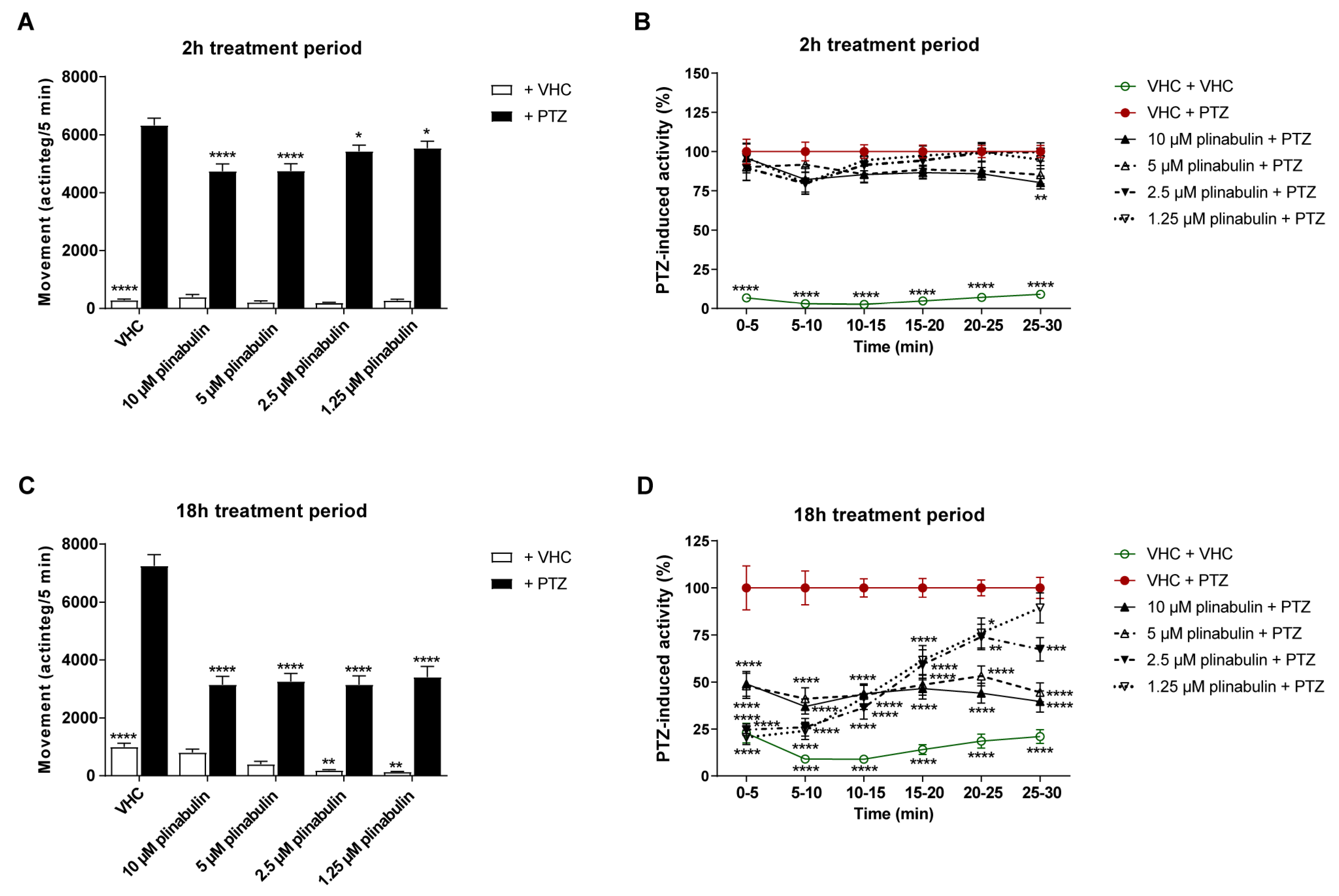
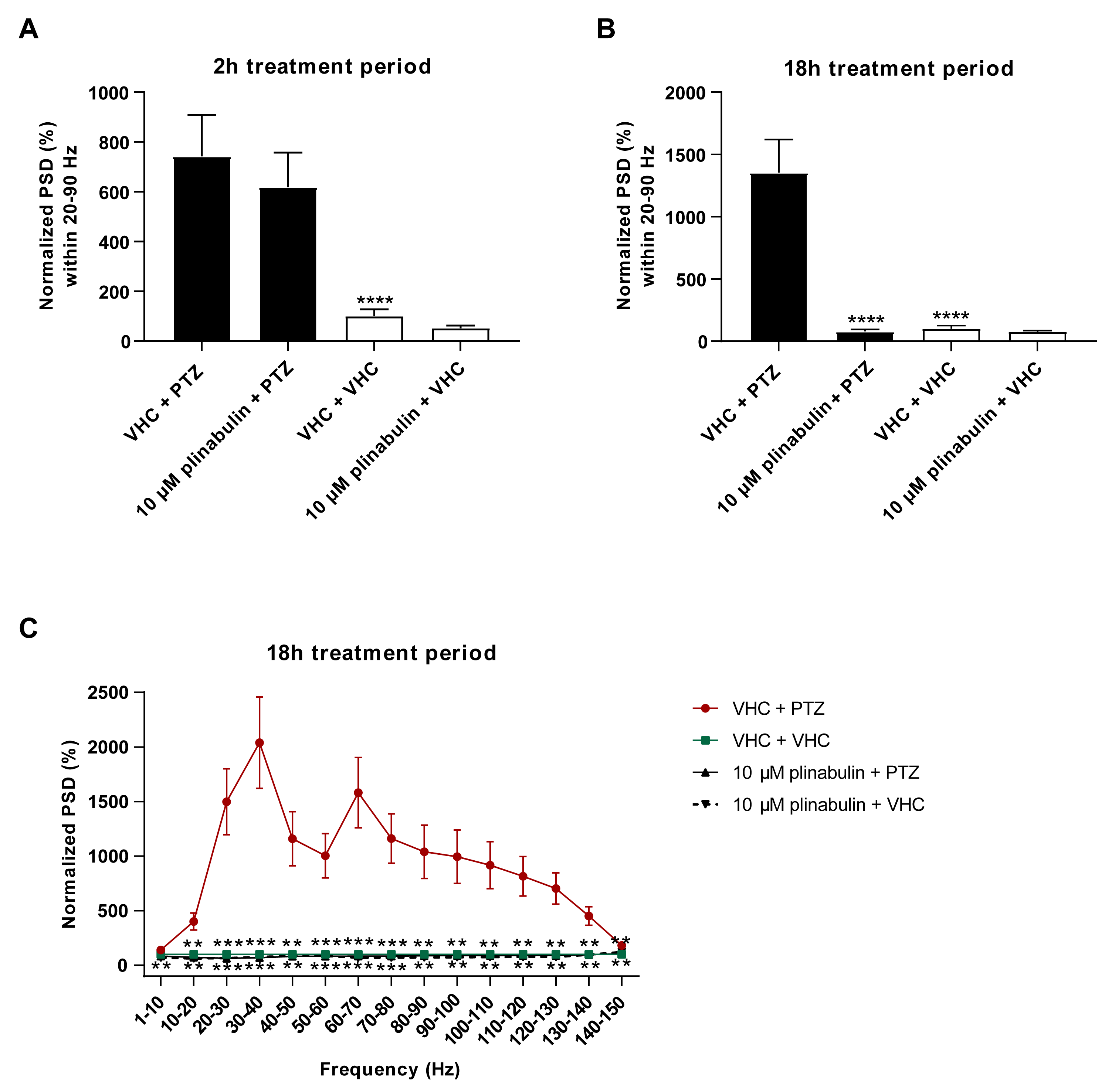
2.4. Plinabulin Ameliorates Seizures and Epileptiform Brain Activity in the Zebrafish PTZ Seizure Model
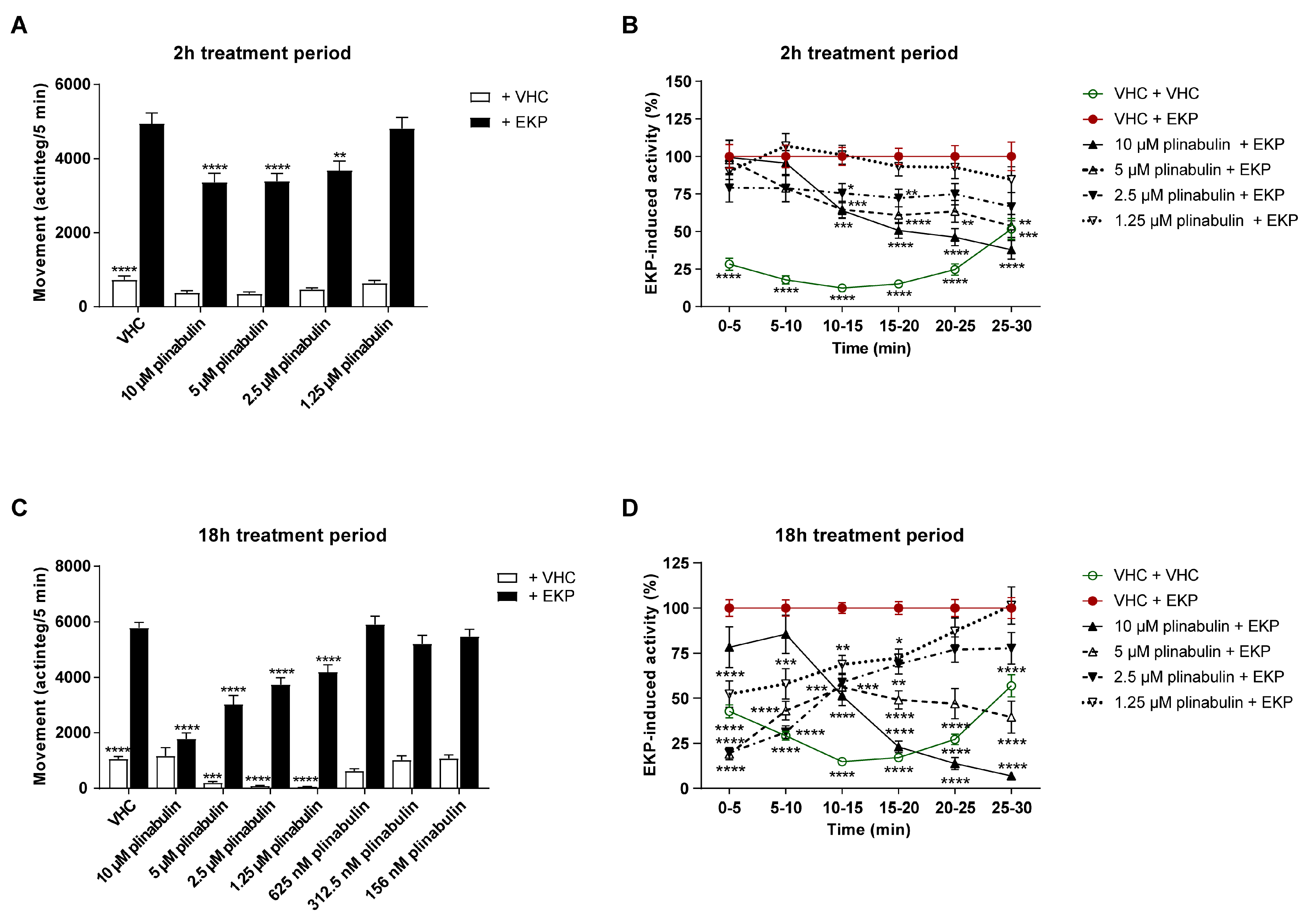
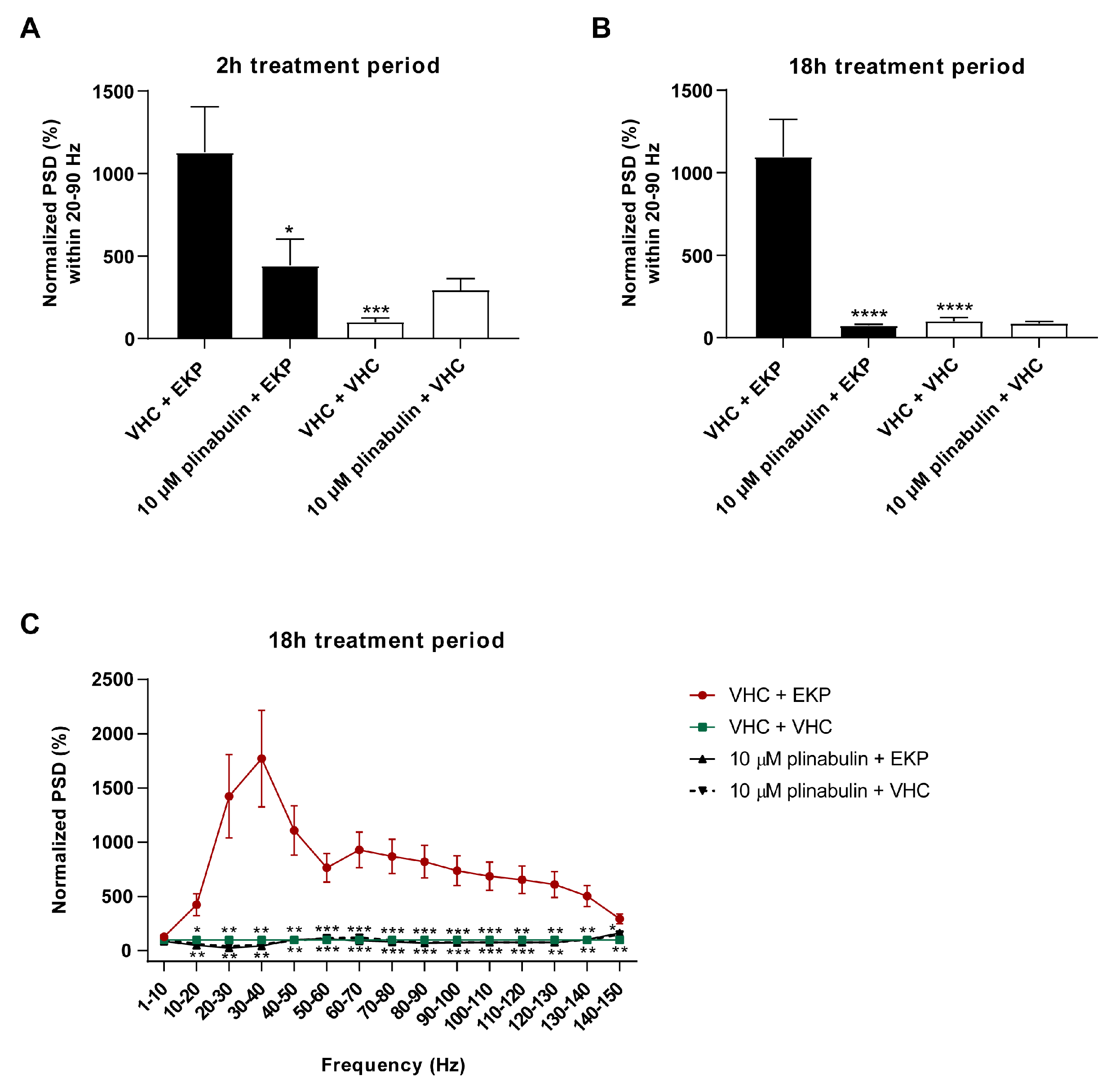
2.5. Plinabulin Ameliorates Seizures and Epileptiform Brain Activity in the Zebrafish EKP Seizure Model
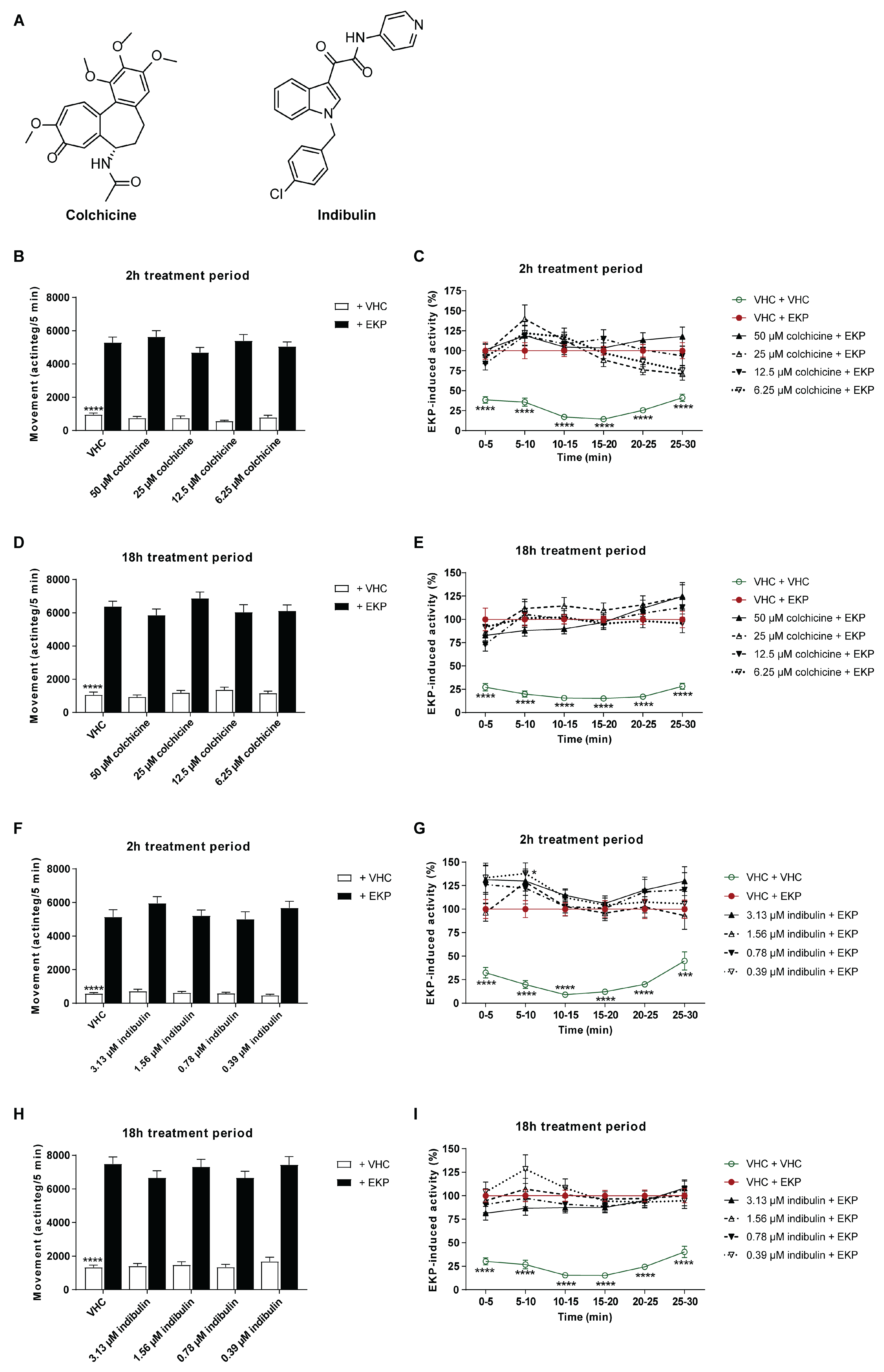
2.6. Microtubule Destabilization Is Insufficient for the Antiseizure Mechanism of Action of Plinabulin
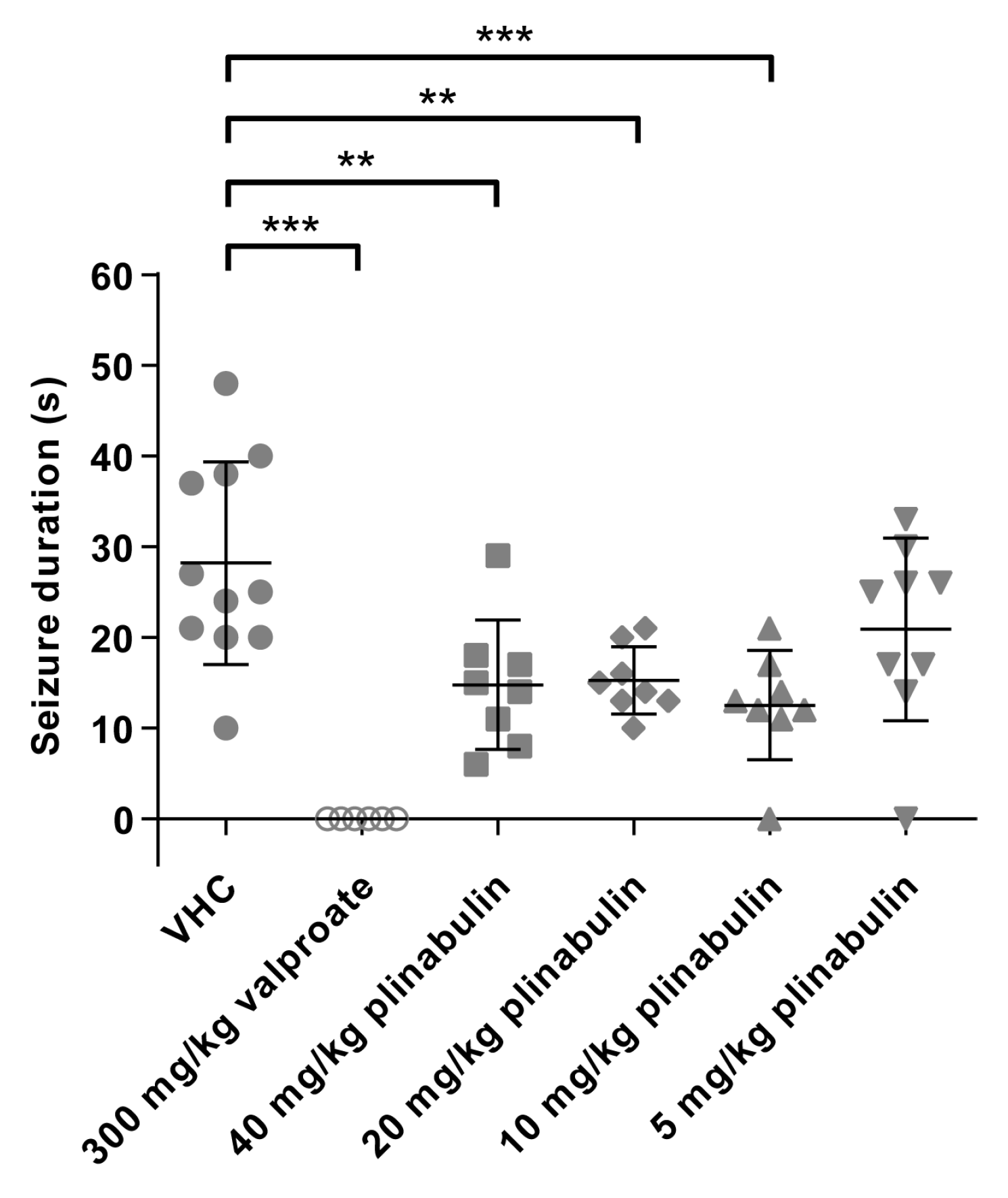
2.7. Plinabulin Ameliorates Seizures in the Mouse 6 Hz Psychomotor Seizure Model
3. Discussion
4. Materials and Methods
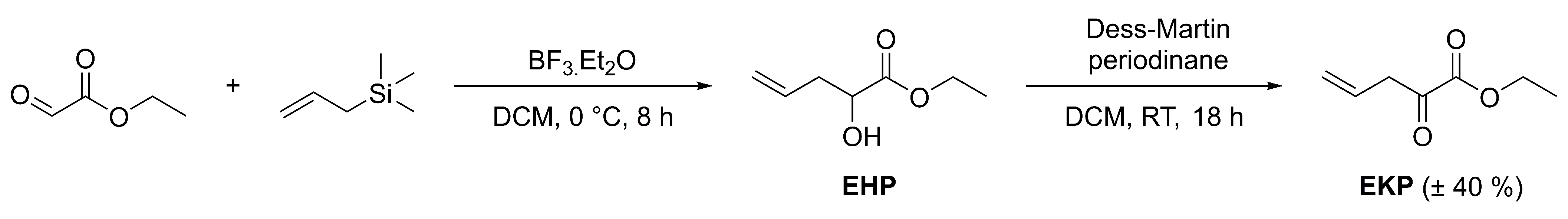
4.1. Chemical Experimental Procedures
4.2. Microbial Strain and Microbial Cultivation
4.3. Microbial Extraction and Isolation
Marfey’s Analysis
4.4. Compounds
4.5. Compound and Sample Preparation
4.6. Experimental Animals
4.6.1. Zebrafish
4.6.2. Mice
4.7. Toxicity Evaluation
4.8. Behavioral Analysis
4.9. Electrophysiology
4.10. Mouse 6 Hz (44 mA) Psychomotor Seizure Model
4.11. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Marcel, J.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The Marine Biodiscovery Pipeline and Ocean Medicines of Tomorrow. J. Mar. Biol. Assoc. UK 2016, 96, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Rateb, M.; Tabudravu, J.N.; Perez-Bonilla, M.; Dirkx, N.; Vallorani, R.; Diaz, C.; del Palacio, J.P.; Smith, A.J.; Ebel, R.; et al. Zebrafish-Based Discovery of Antiseizure Compounds from the Red Sea: Pseurotin A2 and Azaspirofuran A. ACS Chem. Neurosci. 2018, 9, 1652–1662. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Kildgaard, S.; Rasmussen, S.A.; Slezak, M.; Dirkx, N.; Partoens, M.; Esguerra, C.V.; Crawford, A.D.; Larsen, T.O.; de Witte, P.A.M. Zebrafish-Based Discovery of Antiseizure Compounds from the North Sea: Isoquinoline Alkaloids TMC-120A and TMC-120B. Mar. Drugs 2019, 17, 607. [Google Scholar] [CrossRef] [Green Version]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine Microorganisms as a Promising and Sustainable Source of Bioactive Molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Sonnenschein, C.E.; Stierhof, M.; Goralczyk, S.; Vabre, F.M.; Pellissier, L.; Hanssen, K.O.; de la Cruz, M.; Diaz, C.; de Witte, P.; Copmans, D.; et al. Pseudochelin A, a Siderophore of Pseudoalteromonas Piscicida S2040. Tetrahedron 2017, 73, 2633–2637. [Google Scholar] [CrossRef] [Green Version]
- Gnavi, G.; Esposito, F.P.; Festa, C.; Poli, A.; Tedesco, P.; Fani, R.; Monti, M.C.; de Pascale, D.; D’Auria, M.V.; Varese, G.C. The Antimicrobial Potential of Algicolous Marine Fungi for Counteracting Multidrug-Resistant Bacteria: Phylogenetic Diversity and Chemical Profiling. Res. Microbiol. 2016, 167, 492–500. [Google Scholar] [CrossRef]
- Tedesco, P.; Maida, I.; Esposito, F.P.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiros, M.; Sanchez, J.A.; Alonso, E.; Rateb, M.E.; Houssen, W.E.; Ebel, R.; Jaspars, M.; Alfonso, A.; Botana, L.M. Spongionella Secondary Metabolites Protect Mitochondrial Function in Cortical Neurons against Oxidative Stress. Mar. Drugs 2014, 12, 700–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, P.; Ma, H.; Zhu, W. Developments around the Bioactive Diketopiperazines: A Patent Review. Expert Opin. Ther. Pat. 2013, 23, 1415–1433. [Google Scholar] [CrossRef]
- Plinabulin. ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=plinabulin&cntry=&state=&city=&dist= (accessed on 16 June 2021).
- Blayney, W.D.; Zhang, Q.; Feng, J.; Zhao, Y.; Bondarenko, I.; Vynnychenko, I.; Kovalenko, N.; Nair, S.; Ibrahim, E.; Udovista, D.P.; et al. Efficacy of Plinabulin Vs Pegfilgrastim for Prevention of Chemotherapy-Induced Neutropenia in Adults with Non-Small Cell Lung Cancer: A Phase 2 Randomized Clinical Trial. JAMA Oncol. 2020, 6, e204429. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Li, F.; Zhong, C.; Li, F.; Liu, Y.; Wang, S.; Zhao, J.; Li, W. Structure-Based Design and Synthesis of Novel Furan-Diketopiperazine-Type Derivatives as Potent Microtubule Inhibitors for Treating Cancer. Bioorg. Med. Chem. 2020, 28, 115435. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.M.; Spear, M.A.; Yee, L.K.; Mita, A.C.; Heath, E.I.; Papadopoulos, K.P.; Federico, K.C.; Reich, S.D.; Romero, O.; Malburg, L.; et al. Phase 1 First-in-Human Trial of the Vascular Disrupting Agent Plinabulin(Npi-2358) in Patients with Solid Tumors or Lymphomas. Clin. Cancer Res. 2010, 16, 5892–5899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millward, M.; Mainwaring, P.; Mita, A.; Federico, K.; Lloyd, G.K.; Reddinger, N.; Nawrocki, S.; Mita, M.; Spear, M.A. Phase 1 Study of the Novel Vascular Disrupting Agent Plinabulin (Npi-2358) and Docetaxel. Investig. New Drugs 2012, 30, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Crawford, D.A.; Esguerra, C.V.; de Witte, P.A. Fishing for Drugs from Nature: Zebrafish as a Technology Platform for Natural Product Discovery. Planta Med. 2008, 74, 624–632. [Google Scholar] [CrossRef] [Green Version]
- MacRae, A.C.; Peterson, R.T. Zebrafish as Tools for Drug Discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Copmans, D.; Siekierska, A.; de Witte, P.A.M. Zebrafish Models of Epilepsy and Epileptic Seizures. In Models of Seizures and Epilepsy; Pitkänen, A., Buckmaster, P.S., Galanopoulou, A.S., Moshé, S.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 369–384. [Google Scholar]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish Disease Models in Drug Discovery: From Preclinical Modelling to Clinical Trials. Nat. Rev. Drug Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef]
- Ngugi, K.A.; Bottomley, C.; Kleinschmidt, I.; Sander, J.W.; Newton, C.R. Estimation of the Burden of Active and Life-Time Epilepsy: A Meta-Analytic Approach. Epilepsia 2010, 51, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Trevick, S. The Epidemiology of Global Epilepsy. Neurol. Clin. 2016, 34, 837–847. [Google Scholar] [CrossRef]
- Fisher, S.R.; Boas, W.V.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J., Jr. Epileptic Seizures and Epilepsy: Definitions Proposed by the International League against Epilepsy (Ilae) and the International Bureau for Epilepsy (Ibe). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Loscher, W.; Potschka, H.; Sisodiya, S.M.; Vezzani, A. Drug Resistance in Epilepsy: Clinical Impact, Potential Mechanisms, and New Innovative Treatment Options. Pharmacol. Rev. 2020, 72, 606–638. [Google Scholar] [CrossRef] [PubMed]
- Loscher, W.; Klein, P. The Feast and Famine: Epilepsy Treatment and Treatment Gaps in Early 21st Century. Neuropharmacology 2020, 170, 108055. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, K.; Kohno, S.; Asari, T.; Harada, T.; Katada, J.; Muramatsu, M.; Kawashima, H.; Sekiya, H.; Uno, I. (−)-Phenylahistin: A New Mammalian Cell Cycle Inhibitor Produced by Aspergillus Ustus. Bioorg. Med. Chem. Lett. 1997, 7, 2847–2852. [Google Scholar] [CrossRef]
- Kanoh, K.; Kohno, S.; Katada, J.; Hayashi, Y.; Muramatsu, M.; Uno, I. Antitumor Activity of Phenylahistin in Vitro and in Vivo. Biosci. Biotechnol. Biochem. 1999, 63, 1130–1133. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, K.; Kohno, S.; Katada, J.; Takahashi, J.; Uno, I. (-)-Phenylahistin Arrests Cells in Mitosis by Inhibiting Tubulin Polymerization. J. Antibiot. 1999, 52, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Vanmeert, M.; Siekierska, A.; Ny, A.; John, J.; Callewaert, G.; Lescrinier, E.; Dehaen, W.; de Witte, P.A.M.; Kaminski, R.M. Inhibition of Glutamate Decarboxylase (Gad) by Ethyl Ketopentenoate (Ekp) Induces Treatment-Resistant Epileptic Seizures in Zebrafish. Sci. Rep. 2017, 7, 7195. [Google Scholar] [CrossRef] [Green Version]
- Barton, E.M.; Klein, B.D.; Wolf, H.H.; White, H.S. Pharmacological Characterization of the 6 Hz Psychomotor Seizure Model of Partial Epilepsy. Epilepsy Res. 2001, 47, 217–227. [Google Scholar] [CrossRef]
- Kehne, H.J.; Klein, B.D.; Raeissi, S.; Sharma, S. The National Institute of Neurological Disorders and Stroke (Ninds) Epilepsy Therapy Screening Program (Etsp). Neurochem. Res. 2017, 42, 1894–1903. [Google Scholar] [CrossRef] [Green Version]
- Kokel, D.; Bryan, J.; Laggner, C.; White, R.; Cheung, C.Y.; Mateus, R.; Healey, D.; Kim, S.; Werdich, A.A.; Haggarty, S.J.; et al. Rapid Behavior-Based Identification of Neuroactive Small Molecules in the Zebrafish. Nat. Chem. Biol. 2010, 6, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Kokel, D.; Dunn, T.W.; Ahrens, M.B.; Alshut, R.; Cheung, C.Y.; Saint-Amant, L.; Bruni, G.; Mateus, R.; van Ham, T.J.; Shiraki, T.; et al. Identification of Nonvisual Photomotor Response Cells in the Vertebrate Hindbrain. J. Neurosci. 2013, 33, 3834–3843. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Meinl, T.; Dietz, C.; van Leeuwen, M.; Ortmann, J.; Berthold, M.R.; de Witte, P.A. A Knime-Based Analysis of the Zebrafish Photomotor Response Clusters the Phenotypes of 14 Classes of Neuroactive Molecules. J. Biomol. Screen. 2016, 21, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Baraban, C.S.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole Induced Changes in Zebrafish Behavior, Neural Activity and C-Fos Expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Afrikanova, T.; Serruys, A.S.; Buenafe, O.E.; Clinckers, R.; Smolders, I.; de Witte, P.A.; Crawford, A.D.; Esguerra, C.V. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor Versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Copmans, D.; Partoens, M.; Hunyadi, B.; Luyten, W.; de Witte, P. Zebrafish-Based Screening of Antiseizure Plants Used in Traditional Chinese Medicine: Magnolia Officinalis Extract and Its Constituents Magnolol and Honokiol Exhibit Potent Anticonvulsant Activity in a Therapy-Resistant Epilepsy Model. ACS Chem. Neurosci. 2020, 11, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Britton, W.J.; Frey, L.C.; Hopp, J.L.; Korb, P.; Koubeissi, M.Z.; Lievens, W.E.; Pestana-Knight, E.M.; Louis, E.K.S. Electroencephalography (EEG): An Introductory Text and Atlas of Normal and Abnormal Findings in Adults, Children, and Infants; St. Louis, E.K., Frey, L.C., Eds.; American Epilepsy Society: Chicago, IL, USA, 2016. [Google Scholar]
- Zdebik, A.A.; Mahmood, F.; Stanescu, H.C.; Kleta, R.; Bockenhauer, D.; Russell, C. Epilepsy in Kcnj10 Morphant Zebrafish Assessed with a Novel Method for Long-Term Eeg Recordings. PLoS ONE 2013, 8, e79765. [Google Scholar] [CrossRef]
- Yakushiji, F.; Muguruma, K.; Hayashi, Y.; Shirasaka, T.; Kawamata, R.; Tanaka, H.; Yoshiwaka, Y.; Taguchi, A.; Takayama, K.; Hayashi, Y. Click Strategy Using Disodium Salts of Amino Acids Improves the Water Solubility of Plinabulin and Kpu-300. Bioorg. Med. Chem. 2017, 25, 3623–3630. [Google Scholar] [CrossRef]
- Ding, Z.; Cheng, H.; Wang, S.; Hou, Y.; Zhao, J.; Guan, H.; Li, W. Development of Mbri-001, a Deuterium-Substituted Plinabulin Derivative as a Potent Anti-Cancer Agent. Bioorg. Med. Chem. Lett. 2017, 27, 1416–1419. [Google Scholar] [CrossRef]
- Joubert, B.; Belbezier, A.; Haesebaert, J.; Rheims, S.; Ducray, F.; Picard, G.; Rogemond, V.; Psimaras, D.; Berzero, G.; Desestret, V.; et al. Long-Term Outcomes in Temporal Lobe Epilepsy with Glutamate Decarboxylase Antibodies. J. Neurol. 2020, 267, 2083–2089. [Google Scholar] [CrossRef]
- Baizabal-Carvallo, J.F. The Neurological Syndromes Associated with Glutamic Acid Decarboxylase Antibodies. J. Autoimmun. 2019, 101, 35–47. [Google Scholar] [CrossRef]
- Lloyd, G.K.; Bossi, L.; Morselli, P.L.; Munari, C.; Rougier, M.; Loiseau, H. Alterations of Gaba-Mediated Synaptic Transmission in Human Epilepsy. Adv. Neurol. 1986, 44, 1033–1044. [Google Scholar] [CrossRef]
- Errichiello, L.; Perruolo, G.; Pascarella, A.; Formisano, P.; Minetti, C.; Striano, S.; Zara, F.; Striano, P. Autoantibodies to Glutamic Acid Decarboxylase (Gad) in Focal and Generalized Epilepsy: A Study on 233 Patients. J. Neuroimmunol. 2009, 211, 120–123. [Google Scholar] [CrossRef]
- Peltola, J.; Kulmala, P.; Isojarvi, J.; Saiz, A.; Latvala, K.; Palmio, J.; Savola, K.; Knip, M.; Keranen, T.; Graus, F. Autoantibodies to Glutamic Acid Decarboxylase in Patients with Therapy-Resistant Epilepsy. Neurology 2000, 55, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, B.; Lloyd, G.K.; Miller, B.R.; Palladino, M.A.; Kiso, Y.; Hayashi, Y.; Neuteboom, S.T. Npi-2358 Is a Tubulin-Depolymerizing Agent: In-Vitro Evidence for Activity as a Tumor Vascular-Disrupting Agent. Anticancer Drugs 2006, 17, 25–31. [Google Scholar] [CrossRef]
- Pereira, B.R.; Evdokimov, N.M.; Lefranc, F.; Valentao, P.; Kornienko, A.; Pereira, D.M.; Andrade, P.B.; Gomes, N.G.M. Marine-Derived Anticancer Agents: Clinical Benefits, Innovative Mechanisms, and New Targets. Mar. Drugs 2019, 17, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.A.; Bandi, M.; Raje, N.; Richardson, P.; Palladino, M.A.; Chauhan, D.; Anderson, K.C. A Novel Vascular Disrupting Agent Plinabulin Triggers Jnk-Mediated Apoptosis and Inhibits Angiogenesis in Multiple Myeloma Cells. Blood 2011, 117, 5692–5700. [Google Scholar] [CrossRef] [PubMed]
- Pascart, T.; Richette, P. Colchicine in Gout: An Update. Curr. Pharm. Des. 2018, 24, 684–689. [Google Scholar] [CrossRef]
- Oostendorp, L.R.; Witteveen, P.O.; Schwartz, B.; Vainchtein, L.D.; Schot, M.; Nol, A.; Rosing, H.; Beijnen, J.H.; Voest, E.E.; Schellens, J.H. Dose-Finding and Pharmacokinetic Study of Orally Administered Indibulin (D-24851) to Patients with Advanced Solid Tumors. Investig. New Drugs 2010, 28, 163–170. [Google Scholar] [CrossRef]
- Saeedian Moghadam, E.; Hamel, E.; Shahsavari, Z.; Amini, M. Synthesis and Anti-Breast Cancer Activity of Novel Indibulin Related Diarylpyrrole Derivatives. DARU J. Pharm. Sci. 2019, 27, 179–189. [Google Scholar] [CrossRef]
- Wilcox, S.K.; Dixon-Salazar, T.; Sills, G.J.; Ben-Menachem, E.; White, H.S.; Porter, R.J.; Dichter, M.A.; Moshe, S.L.; Noebels, J.L.; Privitera, M.D.; et al. Issues Related to Development of New Antiseizure Treatments. Epilepsia 2013, 54 (Suppl. S4), 24–34. [Google Scholar] [CrossRef] [Green Version]
- Fisher, S.R.; Cross, J.H.; D’Souza, C.; French, J.A.; Haut, S.R.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshe, S.L.; et al. Instruction Manual for the Ilae 2017 Operational Classification of Seizure Types. Epilepsia 2017, 58, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Holcomb, J.M.; Dean, R.S. Psychomotor Seizures. In Encyclopedia of Child Behavior and Development; Goldstein, S., Naglieri, J.A., Eds.; Springer: Boston, MA, USA, 2011; pp. 1191–1192. [Google Scholar] [CrossRef]
- Orellana-Paucar, M.A.; Afrikanova, T.; Thomas, J.; Aibuldinov, Y.K.; Dehaen, W.; de Witte, P.A.; Esguerra, C.V. Insights from Zebrafish and Mouse Models on the Activity and Safety of Ar-Turmerone as a Potential Drug Candidate for the Treatment of Epilepsy. PLoS ONE 2013, 8, e81634. [Google Scholar] [CrossRef]
- Molinski, F.T.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug Development from Marine Natural Products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.D.; Cragg, G.M. Drugs and Drug Candidates from Marine Sources: An Assessment of the Current State of Play. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Gerwick, H.W.; Moore, B.S. Lessons from the Past and Charting the Future of Marine Natural Products Drug Discovery and Chemical Biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F. Have Marine Natural Product Drug Discovery Efforts Been Productive and How Can We Improve Their Efficiency? Expert Opin. Drug Discov. 2019, 14, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Clinical Pipeline Marine Pharmacology. Midwestern University. Available online: https://www.midwestern.edu/departments/marinepharmacology/clinical-pipeline (accessed on 5 November 2021).
- Zhang, G.; Li, J.; Zhu, T.; Gu, Q.; Li, D. Advanced Tools in Marine Natural Drug Discovery. Curr. Opin. Biotechnol. 2016, 42, 13–23. [Google Scholar] [CrossRef]
- Pitchai, A.; Rajaretinam, R.K.; Freeman, J.L. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines 2019, 6, 61. [Google Scholar] [CrossRef] [Green Version]
- West, H.K.; Crawford, A.D. Marine Biodiscovery Goes Deeper: Using in Vivo Bioassays Based on Model Organisms to Identify Biomedically Relevant Marine Metabolites. Planta Med. 2016, 82, 754–760. [Google Scholar] [CrossRef]
- Long, M.S.; Liang, F.Y.; Wu, Q.; Lu, X.L.; Yao, X.L.; Li, S.C.; Li, J.; Su, H.; Pang, J.Y.; Pei, Z. Identification of Marine Neuroactive Molecules in Behaviour-Based Screens in the Larval Zebrafish. Mar. Drugs 2014, 12, 3307–3322. [Google Scholar] [CrossRef] [Green Version]
- Bosse, D.G.; Urcino, C.; Watkins, M.; Salcedo, P.F.; Kozel, S.; Chase, K.; Cabang, A.; Espino, S.S.; Safavi-Hemami, H.; Raghuraman, S.; et al. Discovery of a Potent Conorfamide from Conus Episcopatus Using a Novel Zebrafish Larvae Assay. J. Nat. Prod. 2021, 84, 1232–1243. [Google Scholar] [CrossRef]
- Dawidowski, M.; Herold, F.; Chodkowski, A.; Kleps, J.; Szulczyk, P.; Wilczek, M. Synthesis and Anticonvulsant Activity of Novel 2,6-Diketopiperazine Derivatives. Part 1: Perhydropyrrole[1,2-a]Pyrazines. Eur. J. Med. Chem. 2011, 46, 4859–4869. [Google Scholar] [CrossRef] [PubMed]
- Dawidowski, M.; Herold, F.; Chodkowski, A.; Kleps, J. Synthesis and Anticonvulsant Activity of Novel 2,6-Diketopiperazine Derivatives. Part 2: Perhydropyrido[1,2-a]Pyrazines. Eur. J. Med. Chem. 2012, 48, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Cornacchia, C.; Cacciatore, I.; Baldassarre, L.; Mollica, A.; Feliciani, F.; Pinnen, F. 2,5-Diketopiperazines as Neuroprotective Agents. Mini Rev. Med. Chem. 2012, 12, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Faden, I.A.; Knoblach, S.M.; Cernak, I.; Fan, L.; Vink, R.; Araldi, G.L.; Fricke, S.T.; Roth, B.L.; Kozikowski, A.P. Novel Diketopiperazine Enhances Motor and Cognitive Recovery after Traumatic Brain Injury in Rats and Shows Neuroprotection in Vitro and in Vivo. J. Cereb. Blood Flow Metab. 2003, 23, 342–354. [Google Scholar] [CrossRef] [Green Version]
- Faden, I.A.; Fox, G.B.; Di, X.; Knoblach, S.M.; Cernak, I.; Mullins, P.; Nikolaeva, M.; Kozikowski, A.P. Neuroprotective and Nootropic Actions of a Novel Cyclized Dipeptide after Controlled Cortical Impact Injury in Mice. J. Cereb. Blood Flow Metab. 2003, 23, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Faden, I.A.; Knoblach, S.M.; Movsesyan, V.A.; Lea, P.M.T.; Cernak, I. Novel Neuroprotective Tripeptides and Dipeptides. Ann. N. Y. Acad. Sci. 2005, 1053, 472–481. [Google Scholar] [CrossRef]
- Sourbron, J.; Schneider, H.; Kecskes, A.; Liu, Y.; Buening, E.M.; Lagae, L.; Smolders, I.; de Witte, P. Serotonergic Modulation as Effective Treatment for Dravet Syndrome in a Zebrafish Mutant Model. ACS Chem. Neurosci. 2016, 7, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, H.; Gigant, B.; Yu, Y.; Wu, Y.; Chen, X.; Lai, Q.; Yang, Z.; Chen, Q.; Yang, J. Structures of a Diverse Set of Colchicine Binding Site Inhibitors in Complex with Tubulin Provide a Rationale for Drug Discovery. FEBS J. 2016, 283, 102–111. [Google Scholar] [CrossRef]
- Choi, Y.B.; Suh, S.W. Antimicrotubule Agent-Induced Zinc Neurotoxicity. Biol. Pharm. Bull. 2018, 41, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, S.A.; Fernandez-Rodriguez, L.; Zhao, Y.; Monaco, G.; Trefny, M.P.; Yoshida, N.; Martin, K.; Sharma, A.; Olieric, N.; Shah, P.; et al. Gef-H1 Signaling Upon Microtubule Destabilization Is Required for Dendritic Cell Activation and Specific Anti-Tumor Responses. Cell Rep. 2019, 28, 3367–3380.e8. [Google Scholar] [CrossRef] [Green Version]
- La Sala, G.; Olieric, N.; Sharma, A.; Viti, F.; Perez, F.D.B.; Huang, L.; Tonra, J.R.; Lloyd, G.K.; Decherchi, S.; Diaz, J.F.; et al. Structure, Thermodynamics, and Kinetics of Plinabulin Binding to Two Tubulin Isotypes. Chem 2019, 5, 2969–2986. [Google Scholar] [CrossRef]
- Grisham, R.; Ky, B.; Tewari, K.S.; Chaplin, D.J.; Walker, J. Clinical Trial Experience with Ca4p Anticancer Therapy: Focus on Efficacy, Cardiovascular Adverse Events, and Hypertension Management. Gynecol. Oncol. Res. Pract. 2018, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, R.; Marx, G.; Gillett, M.; Kark, A.; Arunanthy, S. Colchicine-Induced Bone Marrow Suppression: Treatment with Granulocyte Colony-Stimulating Factor. J. Emerg. Med. 2000, 18, 435–440. [Google Scholar] [CrossRef]
- Gambino, G.; Rizzo, V.; Giglia, G.; Ferraro, G.; Sardo, P. Microtubule Dynamics and Neuronal Excitability: Advances on Cytoskeletal Components Implicated in Epileptic Phenomena. Cell. Mol. Neurobiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lasser, M.; Tiber, J.; Lowery, L.A. The Role of the Microtubule Cytoskeleton in Neurodevelopmental Disorders. Front. Cell. Neurosci. 2018, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Broekaart, M.D.W.; Anink, J.J.; Baayen, J.C.; Idema, S.; de Vries, H.E.; Aronica, E.; Gorter, J.A.; van Vliet, E.A. Activation of the Innate Immune System Is Evident Throughout Epileptogenesis and Is Associated with Blood-Brain Barrier Dysfunction and Seizure Progression. Epilepsia 2018, 59, 1931–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, Y.; Aks, S.E.; Hutson, J.R.; Juurlink, D.N.; Nguyen, P.; Dubnov-Raz, G.; Pollak, U.; Koren, G.; Bentur, Y. Colchicine Poisoning: The Dark Side of an Ancient Drug. Clin. Toxicol. 2010, 48, 407–414. [Google Scholar] [CrossRef]
- Subbiah, M.I.; Lenihan, D.J.; Tsimberidou, A.M. Cardiovascular Toxicity Profiles of Vascular-Disrupting Agents. Oncologist 2011, 16, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Orellana-Paucar, A.M.; Steurs, G.; Zhang, Y.; Ny, A.; Foubert, K.; Exarchou, V.; Siekierska, A.; Kim, Y.; de Borggraeve, W.; et al. Methylated Flavonoids as Anti-Seizure Agents: Naringenin 4′,7-Dimethyl Ether Attenuates Epileptic Seizures in Zebrafish and Mouse Models. Neurochem. Int. 2018, 112, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Hunyadi, B.; Siekierska, A.; Sourbron, J.; Copmans, D.; de Witte, P.A.M. Automated Analysis of Brain Activity for Seizure Detection in Zebrafish Models of Epilepsy. J. Neurosci. Methods 2017, 287, 13–24. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Halimide | ||
|---|---|---|
| Position | δ1H (Mult, J) | δ13C |
| 1 | - | - |
| 2 | 7.67 s | 135.1 |
| 3 | - | - |
| 4 | - | 132.4 |
| 5 | - | 138.7 |
| 6 | 6.60 s | 107.2 |
| 7 | - | 124.4 |
| 8 | - | - |
| 9 | - | 167.4 |
| 10 | 4.46 t(4.4) | 58.1 |
| 11 | - | - |
| 12 | - | 162.3 |
| 13 | 3.06 dd(13.6,4.4) | 41.4 |
| 3.27 dd(13.6,4.4) | ||
| 14 | - | 135.9 |
| 15/19 | 7.18 m | 131.5 |
| 16/18 | 7.20 m | 129.6 |
| 17 | 7.15 m | 128.4 |
| 20 | - | 38.8 |
| 21 | 5.99 dd(17.5,10.6) | 146.6 |
| 22 | 5.02 dd(17.5,1.0) | 113.1 |
| 5.08 dd(10.6,1.0) | ||
| 23/24 | 1.41 d(6.6) | 28.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copmans, D.; Kildgaard, S.; Roux, E.; Partoens, M.; Steurs, G.; Wang, X.; De Borggraeve, W.M.; Esguerra, C.V.; Crawford, A.D.; Larsen, T.O.; et al. From the North Sea to Drug Repurposing, the Antiseizure Activity of Halimide and Plinabulin. Pharmaceuticals 2022, 15, 247. https://doi.org/10.3390/ph15020247
Copmans D, Kildgaard S, Roux E, Partoens M, Steurs G, Wang X, De Borggraeve WM, Esguerra CV, Crawford AD, Larsen TO, et al. From the North Sea to Drug Repurposing, the Antiseizure Activity of Halimide and Plinabulin. Pharmaceuticals. 2022; 15(2):247. https://doi.org/10.3390/ph15020247
Chicago/Turabian StyleCopmans, Daniëlle, Sara Kildgaard, Emma Roux, Michèle Partoens, Gert Steurs, Xinhui Wang, Wim M. De Borggraeve, Camila V. Esguerra, Alexander D. Crawford, Thomas O. Larsen, and et al. 2022. "From the North Sea to Drug Repurposing, the Antiseizure Activity of Halimide and Plinabulin" Pharmaceuticals 15, no. 2: 247. https://doi.org/10.3390/ph15020247
APA StyleCopmans, D., Kildgaard, S., Roux, E., Partoens, M., Steurs, G., Wang, X., De Borggraeve, W. M., Esguerra, C. V., Crawford, A. D., Larsen, T. O., & de Witte, P. A. M. (2022). From the North Sea to Drug Repurposing, the Antiseizure Activity of Halimide and Plinabulin. Pharmaceuticals, 15(2), 247. https://doi.org/10.3390/ph15020247