
Experimental Evidence for Diiodohydroxyquinoline-Induced Neurotoxicity: Characterization of Age and Gender as Predisposing Factors

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
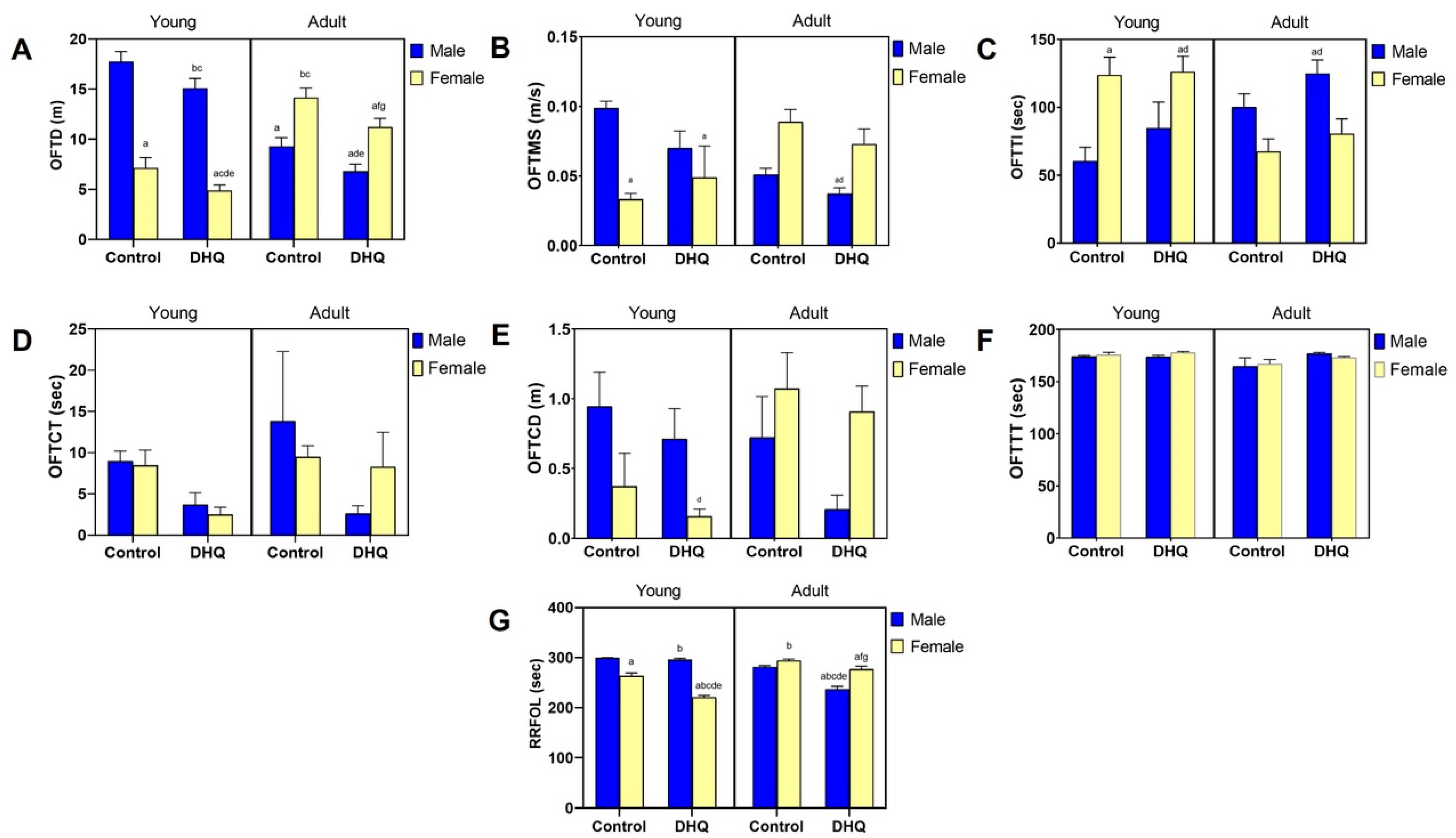
2.1. Motor Impairments Instigated by Diiodohydroxyquinoline Administration in Young/Adult Male/Female Rats
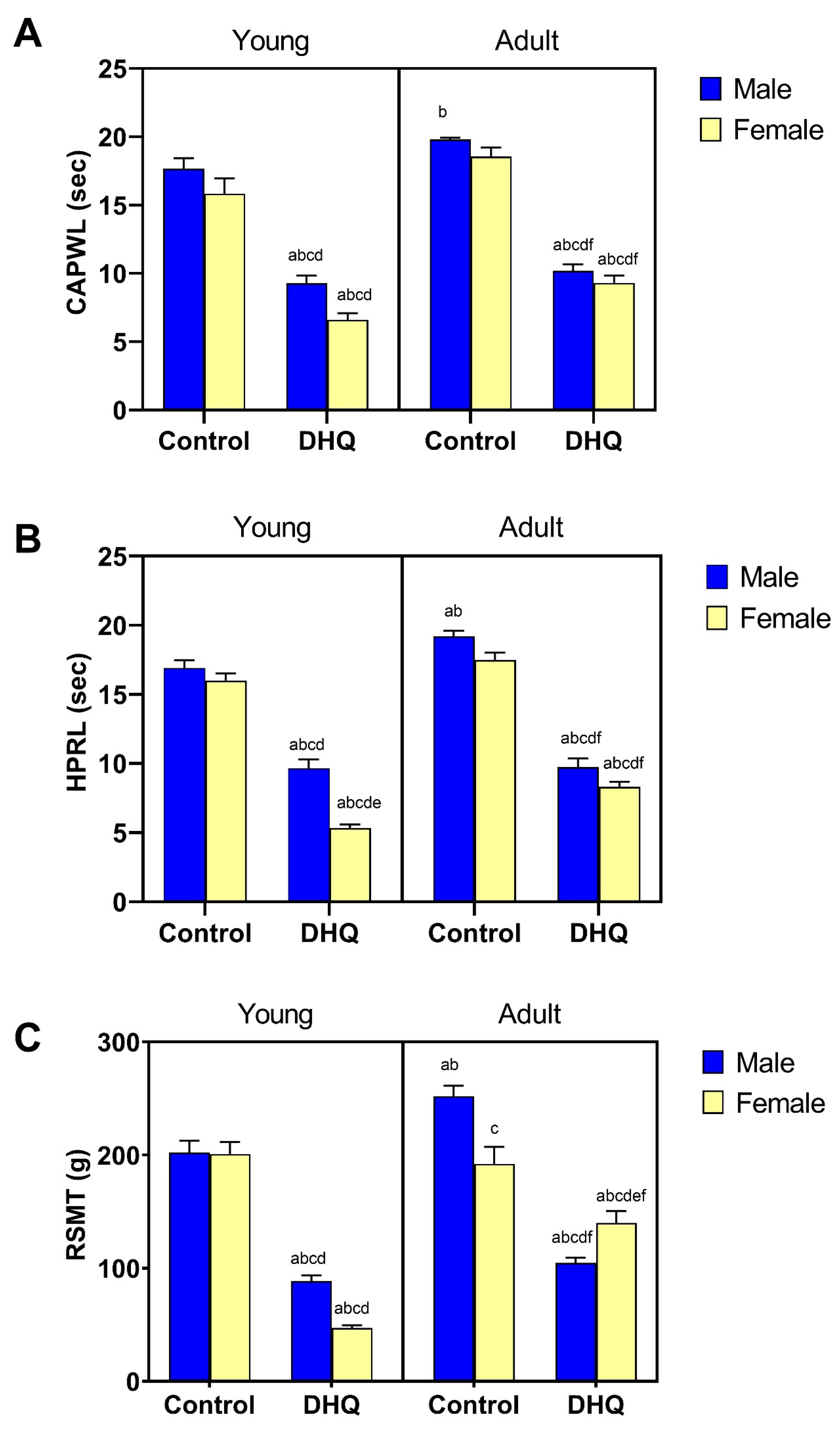
2.2. Sensory Impairments Instigated by Diiodohydroxyquinoline Administration in Young/Adult Male/Female Rats
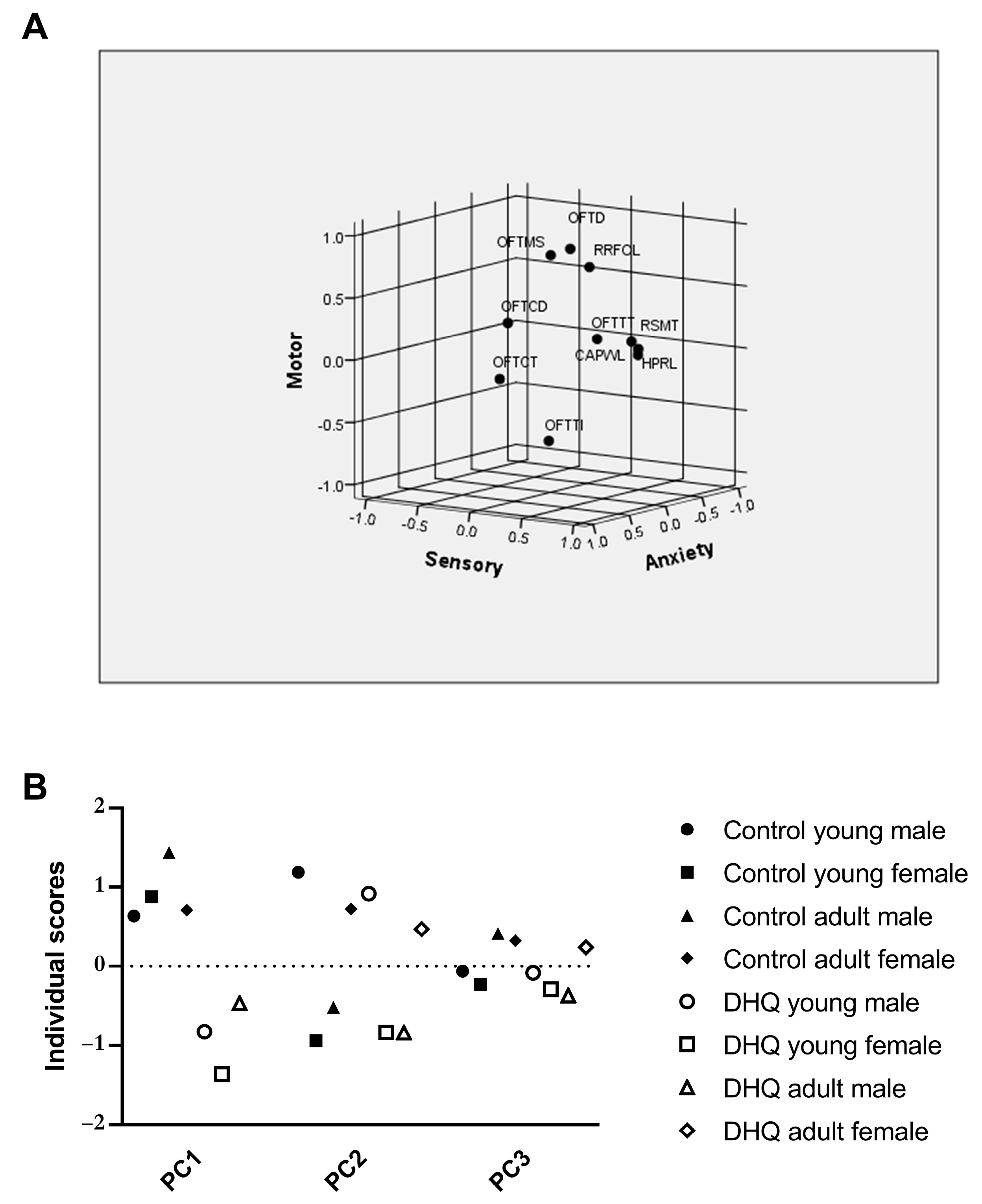
2.3. Linear Regression and Principal Component Analysis
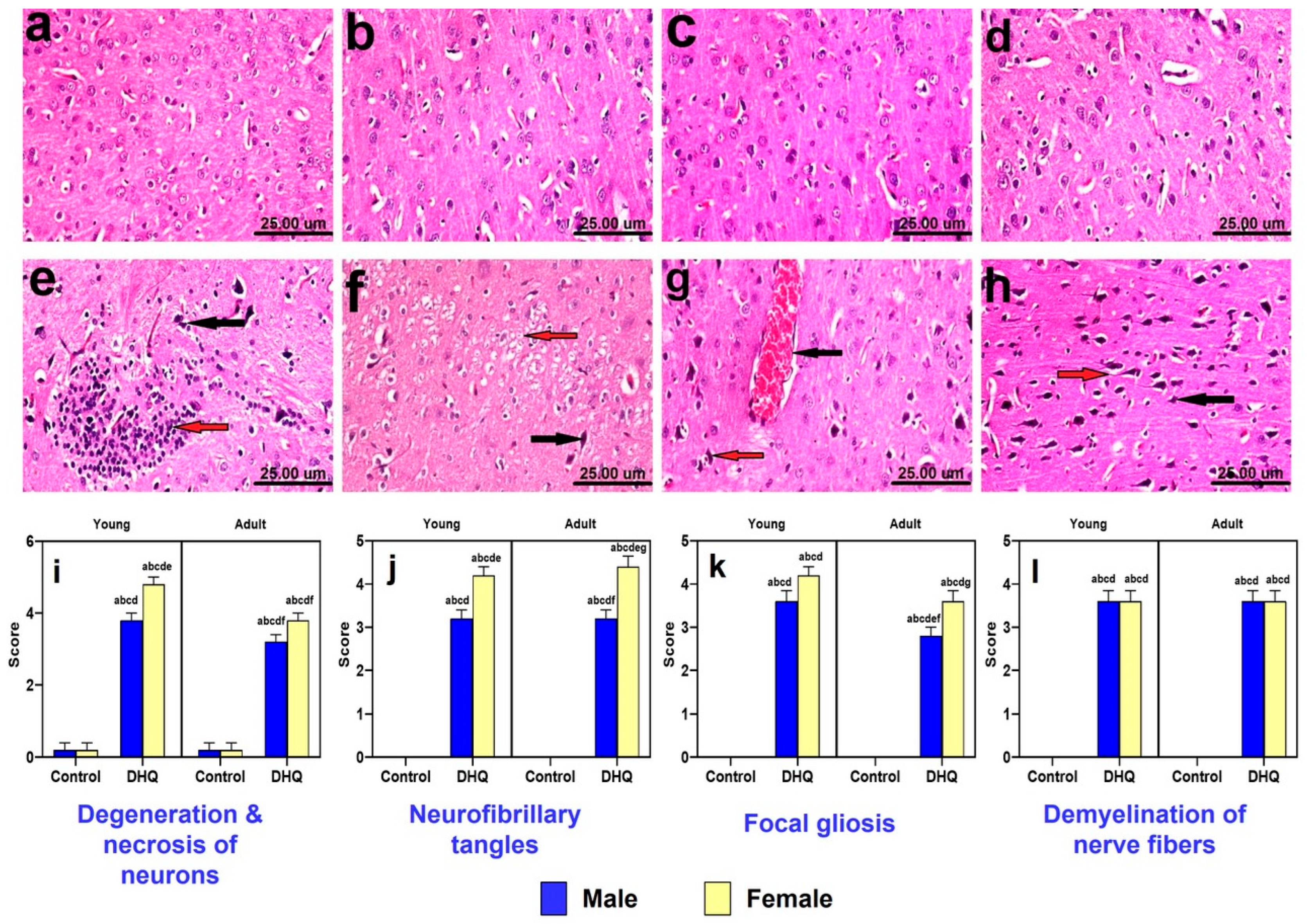
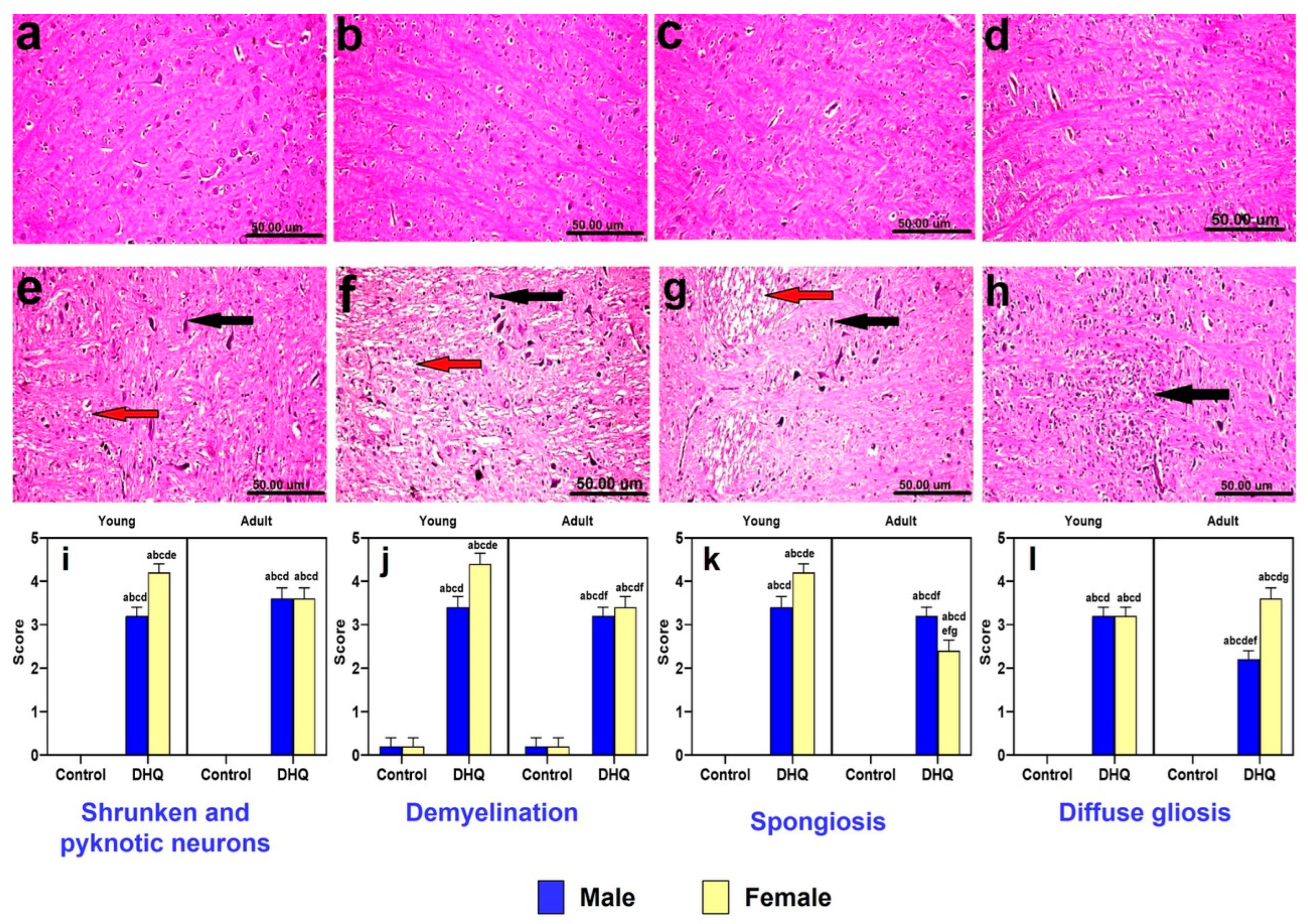
2.4. Histopathological Alterations Instigated by Diiodohydroxyquinoline Administration in Cerebral Cortices of Young/Adult Male/Female Rats
2.5. Histopathological Alterations Instigated by Diiodohydroxyquinoline Administration in Striatum of Young/Adult Male/Female Rats
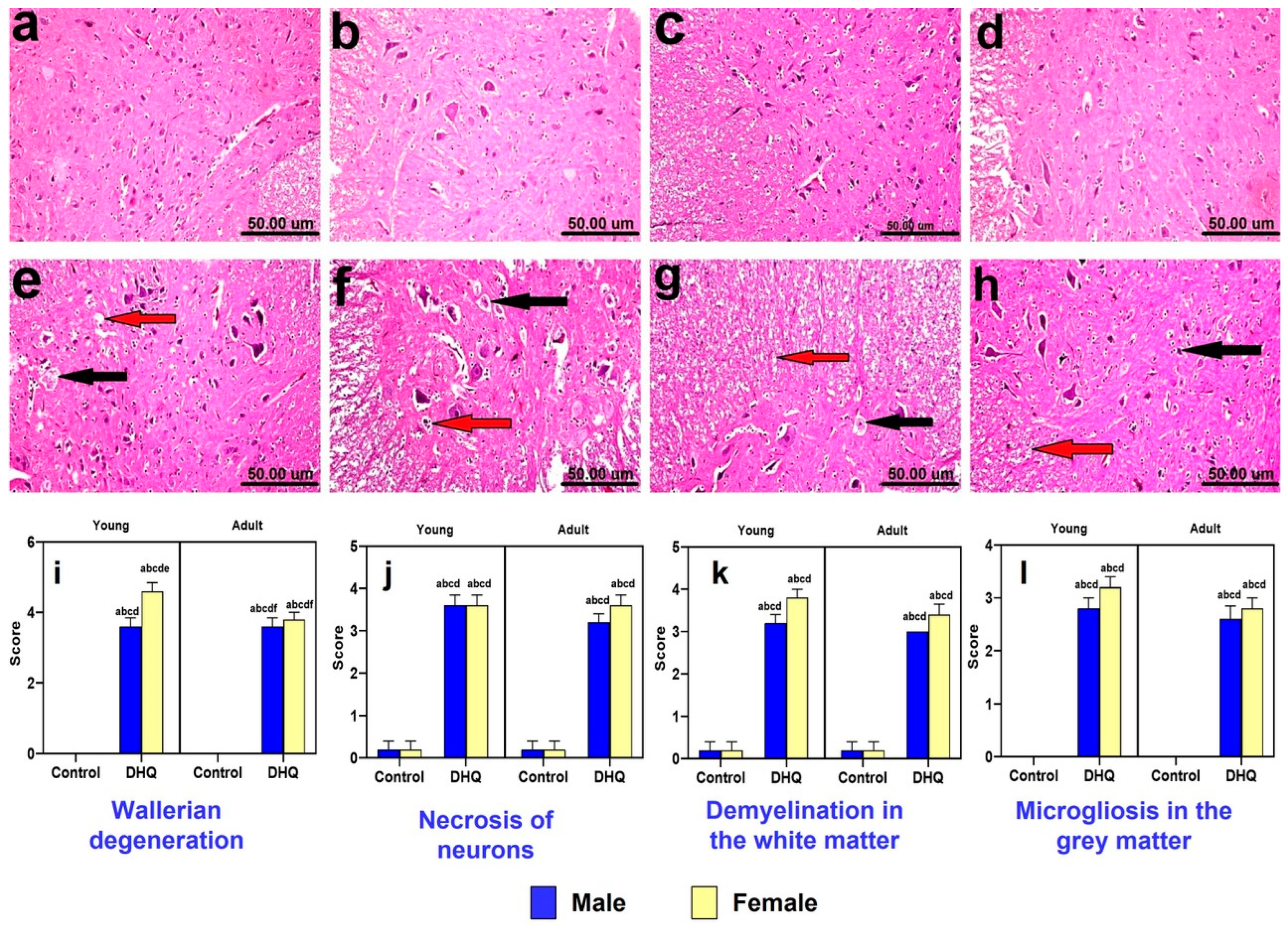
2.6. Histopathological Alterations Instigated by Diiodohydroxyquinoline Administration in Spinal Cords of Young/Adult Male/Female Rats
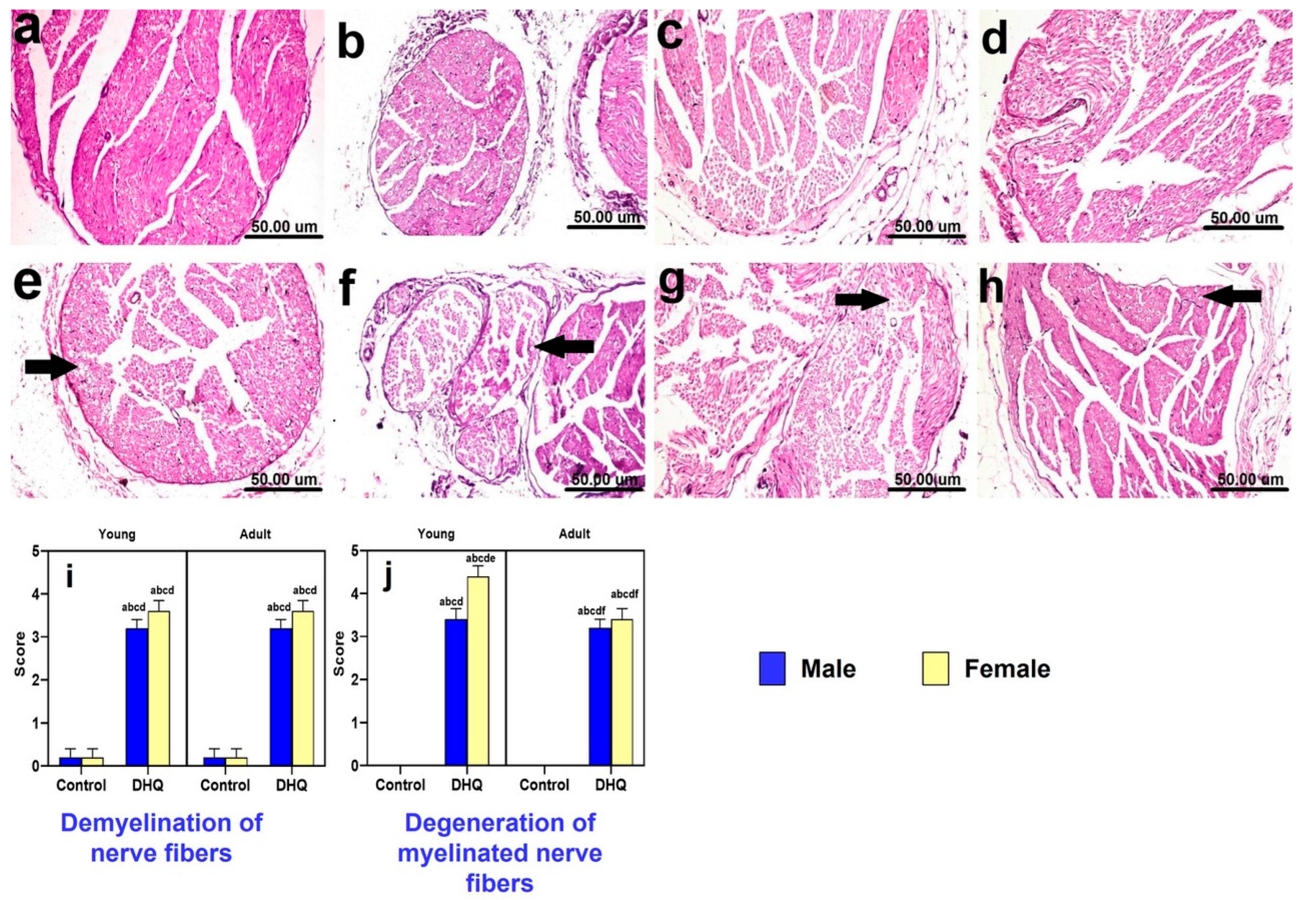
2.7. Histopathological Alterations Instigated by Diiodohydroxyquinoline Administration in Sciatic Nerves of Young/Adult Male/Female Rats
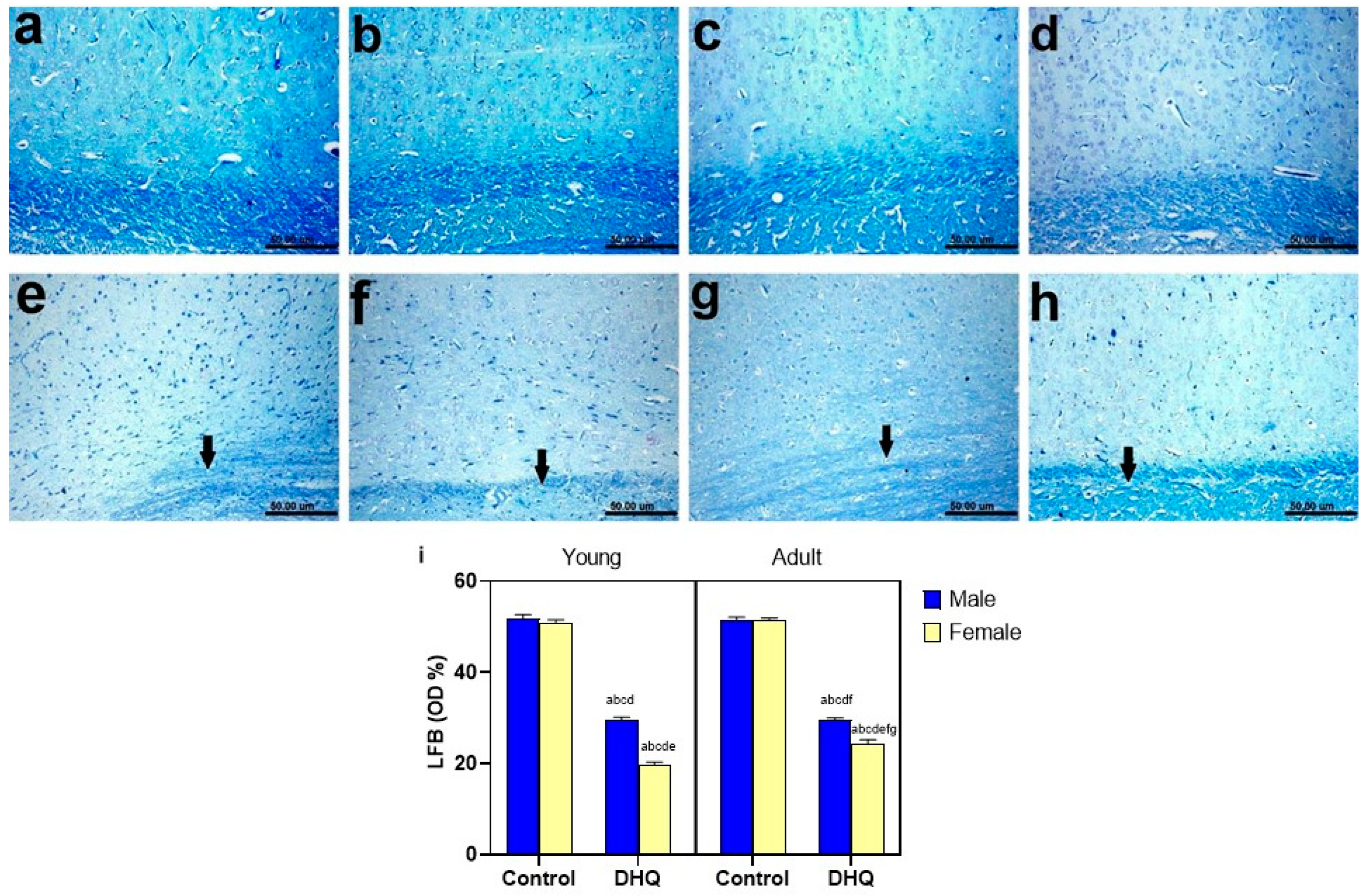
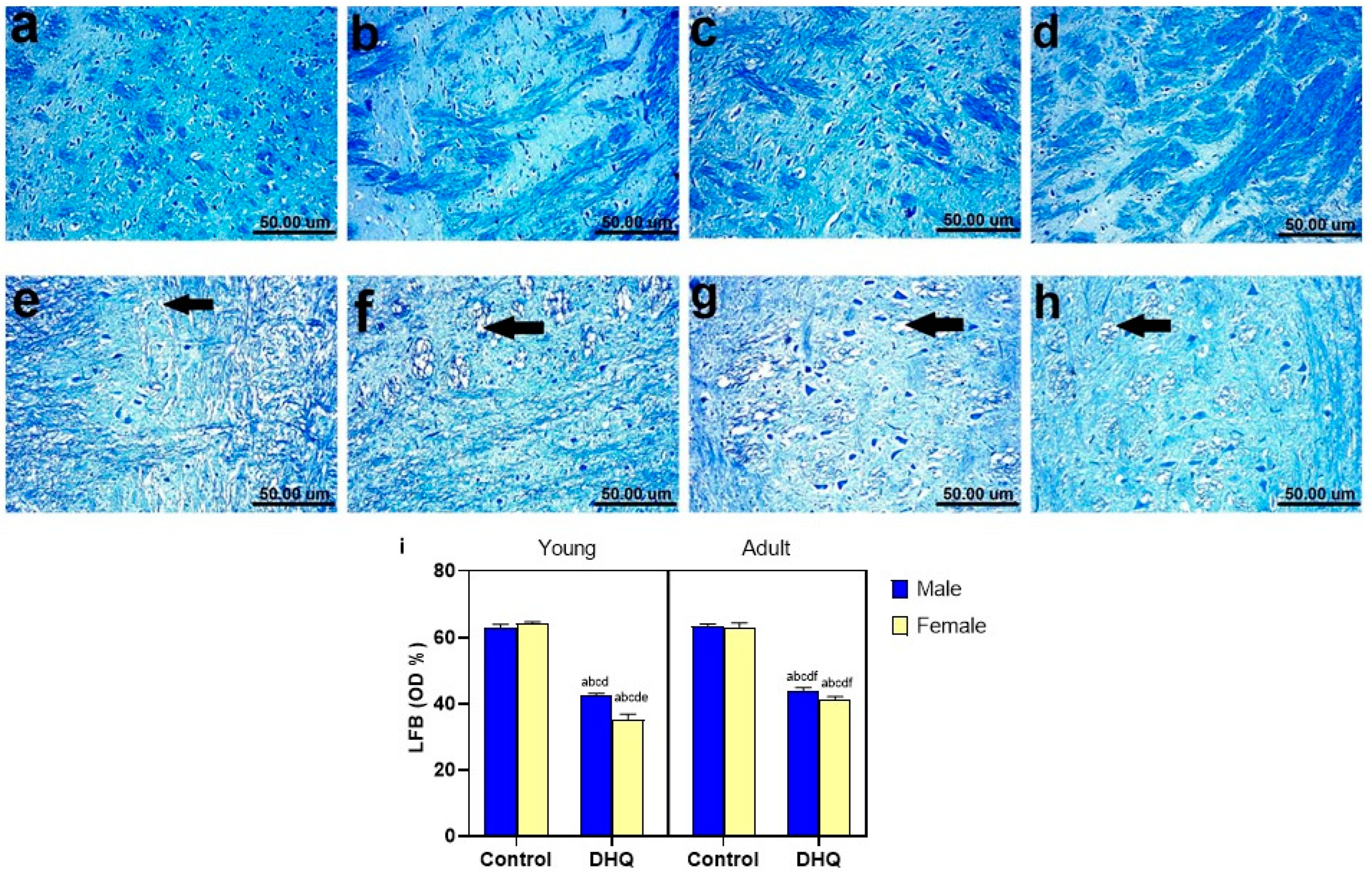
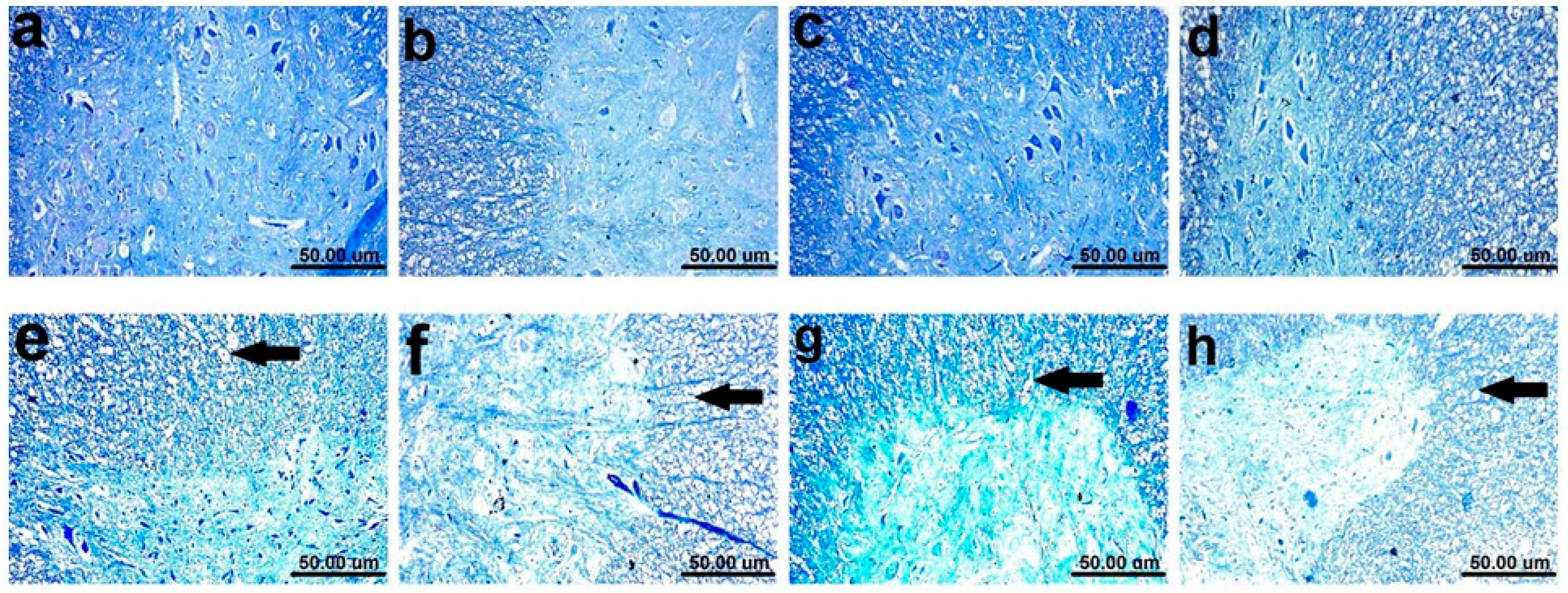
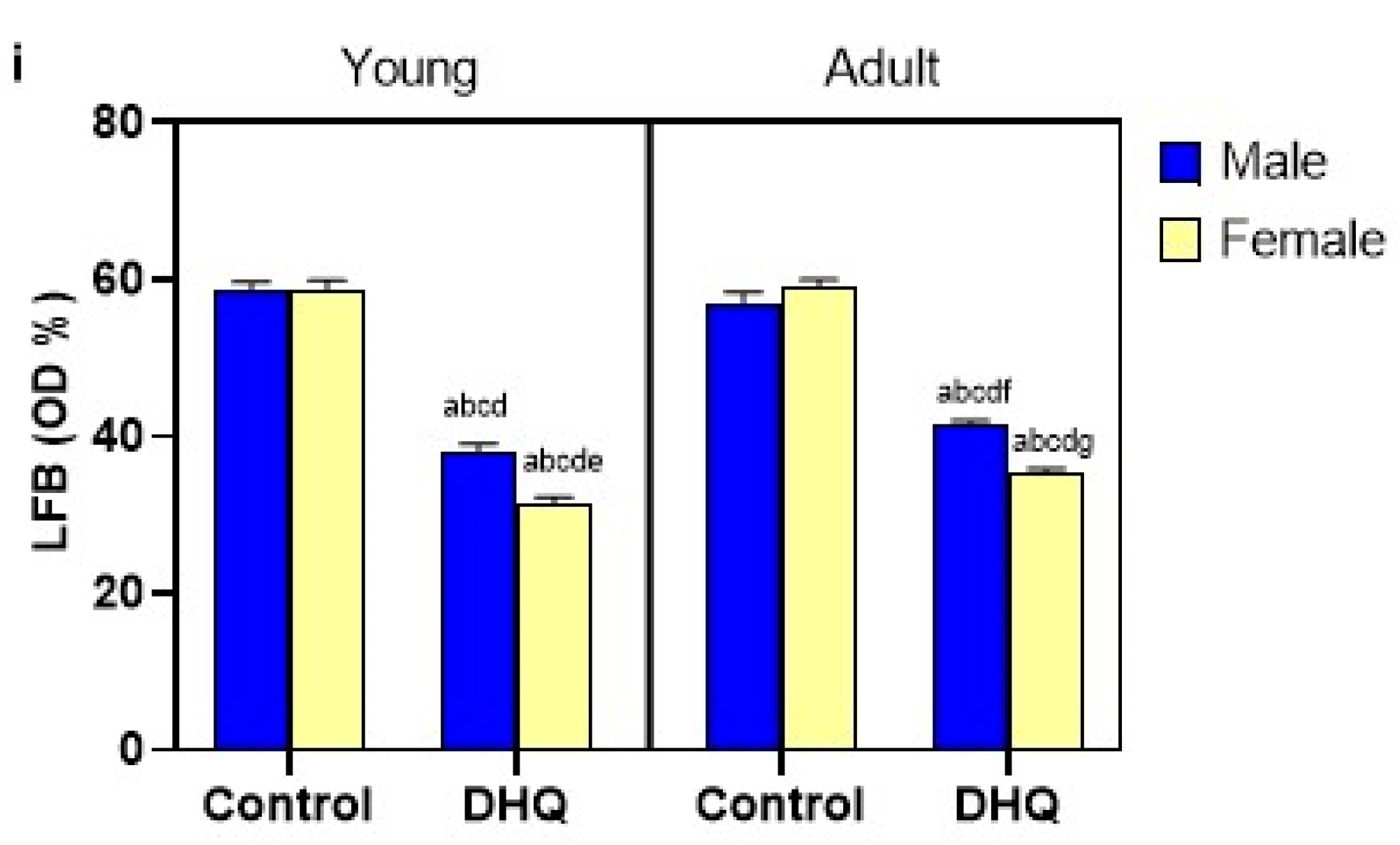
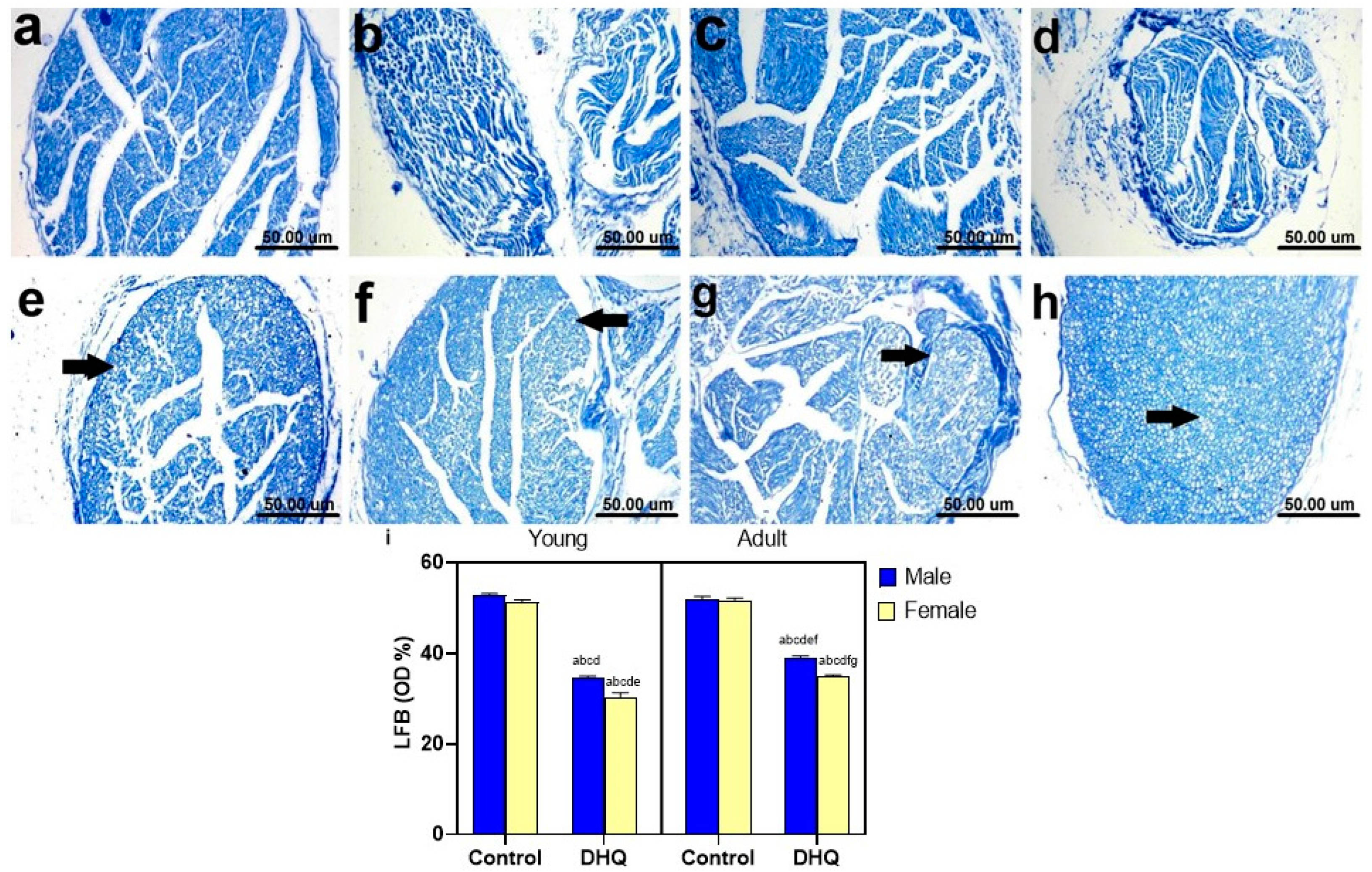
2.8. Myelin Alterations Instigated by Diiodohydroxyquinoline Administration in Cortex, Striatum, Spinal Cords, and Sciatic Nerves of Young/Adult Male/Female Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Chemicals
4.3. Experimental Design
4.4. Behavioral Assessment
4.4.1. Open Field Test
4.4.2. Rotarod Test
4.4.3. Randall–Selitto Test
4.4.4. Hot Plate Test
4.4.5. Hind Paw Cold Allodynia Test
4.5. Histological Assessment
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| OFT | Open field test |
| OFTD | Open field test distance |
| OFTMS | Open field test mean speed |
| OFTTI | Open field test time immobile |
| OFTCT | Open field test central time |
| OFTCD | Open field test central distance |
| OFTTT | Open field test thigmotaxis time |
| RRFOL | Rotarod fall off latency |
| CAPWL | Cold allodynia paw withdrawal latency |
| HPRL | Hotplate reaction latency |
| RSMT | Randall–Salyetto mechanical threshold |
| CYM | Control young male |
| CYF | Control young female |
| CAM | Control adult male |
| CAF | Control adult female |
| TYM | Treated young male |
| TYF | Treated young female |
| TAM | Treated adult male |
| TAF | Treated adult female |
| DHQ | Diiodohydroxyquinoline |
| 8-HQ | 8-hydroxyquinoline |
| SMON | Subacute myelo-optic neuropathy |
| PC | Principal component |
| TRPA1 | Transient receptor potential ankyrin 1 |
| H&E | Hematoxylin and Eosin |
| LFB | Luxol fast blue |
| MANOVA | Three-way analysis of variance test |
| PCA | Principal component analysis |
References
- Schnitzer, R.J. IARC monographs on the evaluation of the carcinogenic risk of chemicals to man. Environ. Res. 1978, 17, 151. [Google Scholar] [CrossRef]
- Song, Y.; Xu, H.; Chen, W.; Zhan, P.; Liu, X. 8-Hydroxyquinoline: A privileged structure with a broad-ranging pharmacological potential. Med. Chem. Comm. 2015, 6, 61–74. [Google Scholar] [CrossRef]
- Yamamoto, R.S.; Williams, G.M.; Frankel, H.H.; Weisburger, J.H. 8-Hydroxyquinoline: Chronic toxicity and inhibitory effect on the carcinogenicity of N-2-fluorenylacetamide. Toxicol. Appl. Pharmacol. 1971, 19, 687–698. [Google Scholar] [CrossRef]
- Maronpot, R.R.; Boorman, G.A. Interpretation of rodent hepatocellular proliferative alterations and hepatocellular tumors in chemical safety assessment. Toxicol. Pathol. 1982, 10, 71–78. [Google Scholar] [CrossRef]
- Bareggi, S.R.; Cornelli, U. Clioquinol: Review of its mechanisms of action and clinical uses in neurodegenerative Disorders. CNS Neurosci. Ther. 2012, 18, 41–46. [Google Scholar] [CrossRef]
- Tsubaki, T.; Honma, Y.; Hoshi, M. Neurological syndrome associated with clioquinol. Lancet 1971, 297, 696–697. [Google Scholar] [CrossRef]
- Nakae, K.; Yamamoto, S.I.; Shigematsu, I.; Kono, R. Relation between Subacute Myelo-Optic Neuropathy (S.M.O.N.) and clioquinol: Nationwide survey. Lancet 1973, 301, 171–173. [Google Scholar] [CrossRef]
- Takeda, A.; Takada, S.; Ando, M.; Itagaki, K.; Tamano, H.; Suzuki, M.; Iwaki, H.; Oku, N. Impairment of recognition memory and hippocampal long-term potentiation after acute exposure to clioquinol. Neuroscience 2010, 171, 443–450. [Google Scholar] [CrossRef]
- Mansour-Ghanaei, F.; Dehbashi, N.; Yazdanparast, K.; Shafaghi, A. Efficacy of saccharomyces boulardii with antibiotics in acute amoebiasis. World J. Gastroenterol. 2003, 9, 1832–1833. [Google Scholar] [CrossRef]
- Fisher, A.K.; Walter, F.G.; Szabo, S. Iodoquinol associated seizures and radiopacity. Clin. Toxicol. 1993, 31, 113–120. [Google Scholar] [CrossRef]
- American Academy of Pediatrics Committee on Drugs. Clioquinol (iodochlorhydroxyquin, vioform) and iodoquinol (diiodohydroxyquin): Blindness and Neuropathy. Pediatrics 1990, 86, 797–798. Available online: https://pubmed.ncbi.nlm.nih.gov/2146587/ (accessed on 13 December 2021).
- Ghaskadbi, S.; Vaidya, V.G. In vivo antimutagenic effect of ascorbic acid against mutagenicity of the common antiamebic drug diiodohydroxyquinoline. Mutat. Res. Toxicol. 1989, 222, 219–222. [Google Scholar] [CrossRef]
- Nevin, R.L. Idiosyncratic quinoline central nervous system toxicity: Historical insights into the chronic neurological sequelae of mefloquine. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. Disabling and Potentially Permanent Side Effects Lead to Suspension or Restrictions of Quinolone and Fluoroquinolone Antibiotics; EMA/175398/2019; European Medicines Agency: Amsterdam, Netherlands, 2019; Available online: https://www.ema.europa.eu/en/documents/referral/quinolone-fluoroquinolone-article-31-referral-disabling-potentially-permanent-side-effects-lead_en.pdf (accessed on 13 December 2021).
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Clioquinol and pyrithione activate TRPA1 by increasing intracellular Zn2+. Proc. Natl. Acad. Sci. USA 2009, 106, 8374–8379. [Google Scholar] [CrossRef] [Green Version]
- Konagaya, M.; Matsumoto, A.; Takase, S.; Mizutani, T.; Sobue, G.; Konishi, T.; Hayabara, T.; Iwashita, H.; Ujihira, T.; Miyata, K.; et al. Clinical analysis of longstanding subacute myelo-optico-neuropathy: Sequelae of clioquinol at 32 years after its ban. J. Neurol. Sci. 2004, 218, 85–90. [Google Scholar] [CrossRef]
- Baumgartner, G.; Gawel, M.J.; Kaeser, H.E.; Pallis, C.A.; Rose, F.C.; Schaumburg, H.H.; Thomas, P.K.; Wadia, N.H. Neurotoxicity of halogenated hydroxyquinolines: Clinical analysis of cases reported outside Japan. J. Neurol. Neurosurg. Psychiatry 1979, 42, 1073–1083. [Google Scholar] [CrossRef] [Green Version]
- Pentland, B.; Donald, S.M. Pain in the Guillain-Barré syndrome: A clinical review. Pain 1994, 59, 159–164. [Google Scholar] [CrossRef]
- Wallace, V.C.J.; Cottrell, D.F.; Brophy, P.J.; Fleetwood-Walker, S.M. Focal Lysolecithin-Induced Demyelination of Peripheral Afferents Results in Neuropathic Pain Behavior That Is Attenuated by Cannabinoids. J. Neurosci. 2003, 23, 3221–3233. [Google Scholar] [CrossRef]
- Konagaya, M. SMON: Toxicity of clioquinol and the status quo. Brain Nerve 2015, 67, 49–62. [Google Scholar] [CrossRef]
- Yassin, M.S.; Ekblom, J.; Xilinas, M.; Gottfries, C.G.; Oreland, L. Changes in uptake of vitamin B12 and trace metals in brains of mice treated with clioquinol. J. Neurol. Sci. 2000, 173, 40–44. [Google Scholar] [CrossRef]
- Feyissa, D.D.; Aher, Y.D.; Engidawork, E.; Höger, H.; Lubec, G.; Korz, V. Individual differences in male rats in a behavioral test battery: A multivariate statistical approach. Front. Behav. Neurosci. 2017, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frick, K.M.; Baxter, M.G.; Markowska, A.L.; Olton, D.S.; Price, D.L. Age-related spatial reference and working memory deficits assessed in the water maze. Neurobiol. Aging 1995, 16, 149–160. [Google Scholar] [CrossRef]
- Carreira, M.B.; Cossio, R.; Britton, G.B. Individual and sex differences in high and low responder phenotypes. Behav. Processes 2017, 136, 20–27. [Google Scholar] [CrossRef]
- Catuara-Solarz, S.; Espinosa-Carrasco, J.; Erb, I.; Langohr, K.; Gonzalez, J.R.; Notredame, C.; Dierssen, M. Combined Treatment with Environmental Enrichment and (−)-Epigallocatechin-3-Gallate Ameliorates Learning Deficits and Hippocampal Alterations in a Mouse Model of Down Syndrome. eNeuro 2016, 3, ENEURO.0103-16.2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundblad, M.; Vaudano, E.; Cenci, M.A. Cellular and behavioural effects of the adenosine A2a receptor antagonist KW-6002 in a rat model of L-DOPA-induced dyskinesia. J. Neurochem. 2003, 84, 1398–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahoon, L. The curious case of clioquinol. Nat. Med. 2009, 15, 356–359. [Google Scholar] [CrossRef]
- Ohta, Y.; Hishikawa, N.; Sato, K.; Takemoto, M.; Yamashita, T.; Doutare, S.; Abe, K. Improving anxiety in subacute myelo-optico-neuropathy (SMON) after an automated telephone call service. Intern. Med. 2019, 58, 1081–1085. [Google Scholar] [CrossRef] [Green Version]
- Monville, C.; Torres, E.M.; Dunnett, S.B. Comparison of incremental and accelerating protocols of the rotarod test for the assessment of motor deficits in the 6-OHDA model. J. Neurosci. Methods. 2006, 158, 219–223. [Google Scholar] [CrossRef]
- Bhattacherjee, A.; Winter, M.K.; Eggimann, L.S.; Mu, Y.; Gunewardena, S.; Liao, Z.; Christianson, J.A.; Smith, P.G. Motor, somatosensory, viscerosensory and metabolic impairments in a heterozygous female rat model of rett syndrome. Int. J. Mol. Sci. 2018, 19, 97. [Google Scholar] [CrossRef] [Green Version]
- Walsh, R.N.; Cummins, R.A. The open-field test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Belzung, C.; Griebel, G. Measuring normal and pathological anxiety-like behaviour in mice: A review. Behav. Brain Res. 2001, 125, 141–149. [Google Scholar] [CrossRef]
- Nagaraju, K.; Raben, N.; Loeffler, L.; Parker, T.; Rochon, P.J.; Lee, E.; Danning, C.; Wada, R.; Thompson, C.; Bahtiyar, G.; et al. Conditional up-regulation of MHC class I in skeletal muscle leads to self-sustaining autoimmune myositis and myositis-specific autoantibodies. Proc. Natl. Acad. Sci. USA 2000, 97, 9209–9214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raben, N.; Nagaraju, K.; Lee, E.; Plotz, P. Modulation of disease severity in mice with targeted disruption of the acid α-glucosidase gene. Neuromuscul. Disord. 2000, 10, 283–291. [Google Scholar] [CrossRef]
- Franco-Pons, N.; Torrente, M.; Colomina, M.T.; Vilella, E. Behavioral deficits in the cuprizone-induced murine model of demyelination/remyelination. Toxicol. Lett. 2007, 169, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Tateishi, J. Subacute myelo-optico-neuropathy: Clioquinol intoxication in humans and animals. Neuropathology 2000, 20, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Generoso, J.S.; Giridharan, V.V.; Lee, J.; Macedo, D.; Barichello, T. The role of the microbiota-gut-brain axis in neuropsychiatric disorders. Braz. J. Psychiatry 2021, 43, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Mandell, G.L.; Douglas, R.; Bennett, J. Principles and Practice of Infectious Diseases, 3rd ed.; Churchill Livingstone: New York, NY, USA, 1990. [Google Scholar]
- Yodoxin (Glenwood). PDR Physicians’ Desk Reference, 43rd ed.; Medical Economics Company: Oradell, NJ, USA, 1989. [Google Scholar]
- Nair, A.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic. Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, W.W.; Ismail, H.M.; Khattab, M.M.; Abdelkader, N.F. Cognitive enhancing effect of diapocynin in D-galactose-ovariectomy-induced Alzheimer’s-like disease in rats: Role of ERK, GSK-3β, and JNK signaling. Toxicol. Appl. Pharmacol. 2020, 398, 115028. [Google Scholar] [CrossRef]
- Abdelkader, N.F.; Saad, M.A.; Abdelsalam, R.M. Neuroprotective effect of nebivolol against cisplatin-associated depressive-like behavior in rats. J. Neurochem. 2017, 141, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Sayed, N.H.; Fathy, N.; Kortam, M.A.; Rabie, M.A.; Mohamed, A.F.; Kamel, A.S. Vildagliptin attenuates Huntington’s disease through activation of GLP-1 receptor/PI3K/Akt/BDNF pathway in 3-nitropropionic acid rat model. Neurotherapeutics 2020, 17, 252–268. [Google Scholar] [CrossRef]
- Wang, I.-C.; Chung, C.-Y.; Liao, F.; Chen, C.-C.; Lee, C.-H. Peripheral sensory neuron injury contributes to neuropathic pain in experimental autoimmune encephalomyelitis. Sci. Rep. 2017, 7, 42304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aswar, M.; Kute, P.; Mahajan, S.; Mahajan, U.; Nerurkar, G.; Aswar, U. Protective effect of hesperetin in rat model of partial sciatic nerve ligation induced painful neuropathic pain: An evidence of anti-inflammatory and anti-oxidative activity. Pharmacol. Biochem. Behav. 2014, 124, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Almohaimeed, H.; Amin, H.; Aziz, G.; Saleh, H. Arabic gum aracia improves diabetic peripheral neuropathy in rats: Ultrastructural histopathalogical study. J. Interdiscip. Histopathol. 2019, 7, 1–9. [Google Scholar] [CrossRef]
- Moustafa, P.E.; Abdelkader, N.F.; El Awdan, S.A.; El-Shabrawy, O.A.; Zaki, H.F. Extracellular matrix remodeling and modulation of inflammation and oxidative stress by sulforaphane in experimental diabetic peripheral neuropathy. Inflammation 2018, 41, 1460–1476. [Google Scholar] [CrossRef]
- Metwally, M.M.M.; Ebraheim, L.L.M.; Galal, A.A.A. Potential therapeutic role of melatonin on STZ-induced diabetic central neuropathy: A biochemical, histopathological, immunohistochemical and ultrastructural study. Acta Histochem. 2018, 120, 828–836. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OFTD | OFTMS | OFTTI | OFTCT | OFTCD | OFTTT | RRFOL | CAPWL | HPRL | ||
|---|---|---|---|---|---|---|---|---|---|---|
| OFTMS | R p | 0.5927 <0.0001 | ||||||||
| OFTTI | r p | −0.5052 <0.0001 | −0.4364 0.0001 | |||||||
| OFTCT | r p | 0.0374 0.7550 | 0.0111 0.9263 | −0.1215 0.3092 | ||||||
| OFTCD | r p | 0.3439 0.0030 | 0.1432 0.2300 | −0.3376 0.0037 | 0.5176 <0.0001 | |||||
| OFTTT | r p | −0.1478 0.2151 | −0.0662 0.5803 | 0.0859 0.4731 | −0.7676 <0.0001 | −0.4508 <0.0001 | ||||
| RRFOL | r p | 0.8741 <0.0001 | 0.4706 <0.0001 | −0.5371 <0.0001 | 0.1480 0.2147 | 0.4340 0.0001 | −0.2458 0.0374 | |||
| CAPWL | r p | 0.3865 0.0007 | 0.1917 0.1067 | −0.3196 0.0062 | 0.2710 0.0213 | 0.2633 0.0254 | -0.3138 0.0072 | 0.5724 <0.0001 | ||
| HPRL | r p | 0.3547 0.0022 | 0.1485 0.2132 | −0.2179 0.0659 | 0.2537 0.0315 | 0.2198 0.0635 | −0.2766 0.0186 | 0.5722 <0.0001 | 0.8728 <0.0001 | |
| RSMT | r p | 0.2752 0.0193 | 0.1131 0.3441 | −0.2083 0.0791 | 0.2674 0.0231 | 0.2327 0.0491 | −0.2255 0.0567 | 0.5419 <0.0001 | 0.8355 <0.0001 | 0.8662 <0.0001 |
| Variables | Component | Contribution | ||
|---|---|---|---|---|
| 1 (29.1%) | 2 (27.4%) | 3 (22.0%) | ||
| OFTD | 0.232 | 0.878 | 0.059 | 0.828 |
| OFTMS | −0.013 | 0.790 | −0.021 | 0.625 |
| OFTTI | −0.107 | −0.728 | −0.128 | 0.558 |
| OFTCT | 0.150 | −0.053 | 0.913 | 0.859 |
| OFTCD | 0.079 | 0.361 | 0.700 | 0.626 |
| OFTTT | −0.163 | −0.021 | −0.874 | 0.791 |
| RRFOL | 0.484 | −0.771 | 0.155 | 0.853 |
| CAPWL | 0.901 | 0.217 | 0.173 | 0.888 |
| HPRL | 0.938 | 0.154 | 0.131 | 0.920 |
| RSMT | 0.932 | 0.105 | 0.127 | 0.896 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamel, A.S.; Mohamed, A.F.; Rabie, M.A.; Elsherbiny, M.E.; Ahmed, K.A.; Khattab, M.M.; Abdelkader, N.F. Experimental Evidence for Diiodohydroxyquinoline-Induced Neurotoxicity: Characterization of Age and Gender as Predisposing Factors. Pharmaceuticals 2022, 15, 251. https://doi.org/10.3390/ph15020251
Kamel AS, Mohamed AF, Rabie MA, Elsherbiny ME, Ahmed KA, Khattab MM, Abdelkader NF. Experimental Evidence for Diiodohydroxyquinoline-Induced Neurotoxicity: Characterization of Age and Gender as Predisposing Factors. Pharmaceuticals. 2022; 15(2):251. https://doi.org/10.3390/ph15020251
Chicago/Turabian StyleKamel, Ahmed S., Ahmed F. Mohamed, Mostafa A. Rabie, Marwa E. Elsherbiny, Kawkab A. Ahmed, Mahmoud M. Khattab, and Noha F. Abdelkader. 2022. "Experimental Evidence for Diiodohydroxyquinoline-Induced Neurotoxicity: Characterization of Age and Gender as Predisposing Factors" Pharmaceuticals 15, no. 2: 251. https://doi.org/10.3390/ph15020251
APA StyleKamel, A. S., Mohamed, A. F., Rabie, M. A., Elsherbiny, M. E., Ahmed, K. A., Khattab, M. M., & Abdelkader, N. F. (2022). Experimental Evidence for Diiodohydroxyquinoline-Induced Neurotoxicity: Characterization of Age and Gender as Predisposing Factors. Pharmaceuticals, 15(2), 251. https://doi.org/10.3390/ph15020251