
Effect of Caffeine and Flavonoids on the Binding of Tigecycline to Human Serum Albumin: A Spectroscopic Study and Molecular Docking

 ,
,  ,
,
Abstract
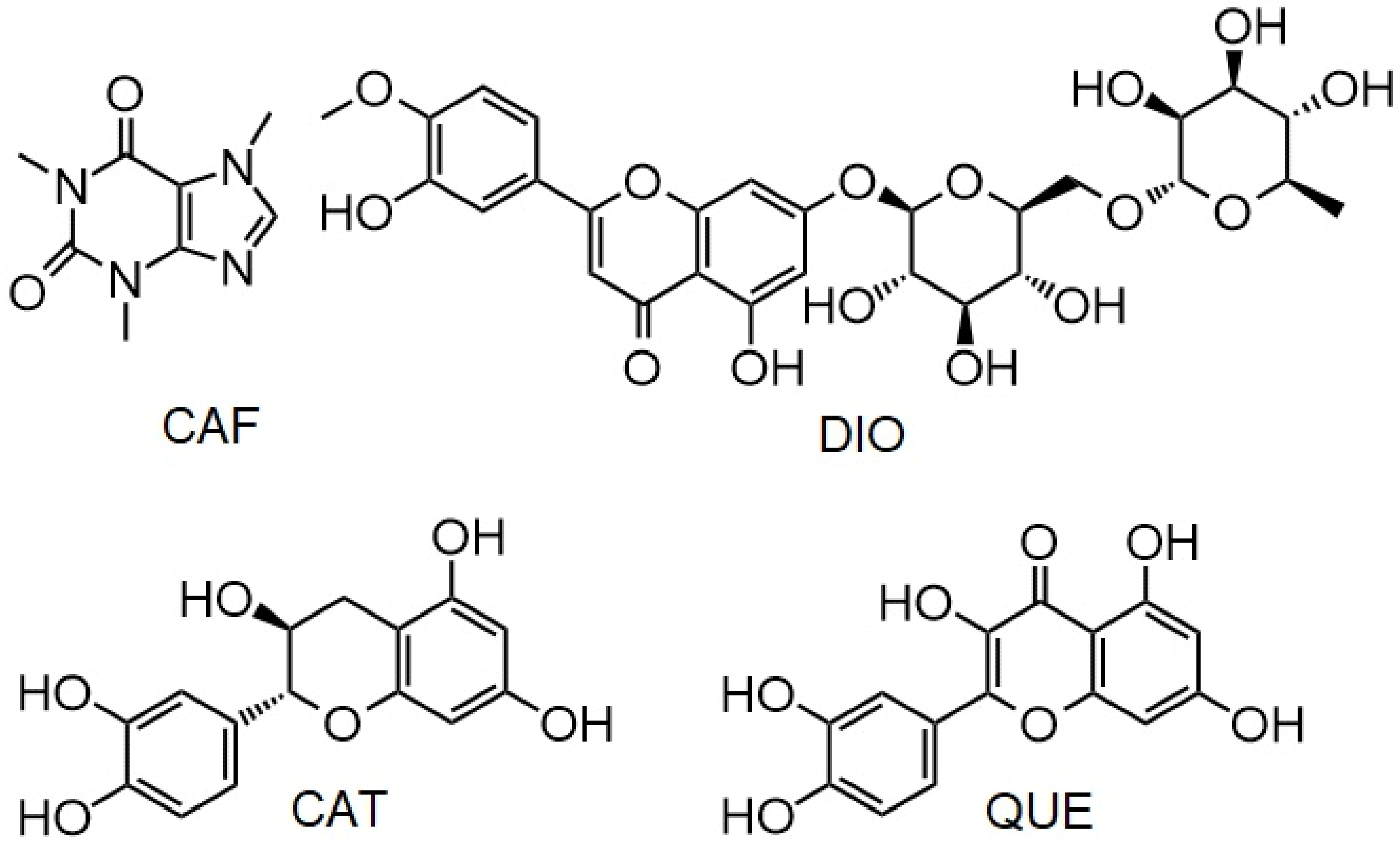
:1. Introduction
2. Results and Discussion
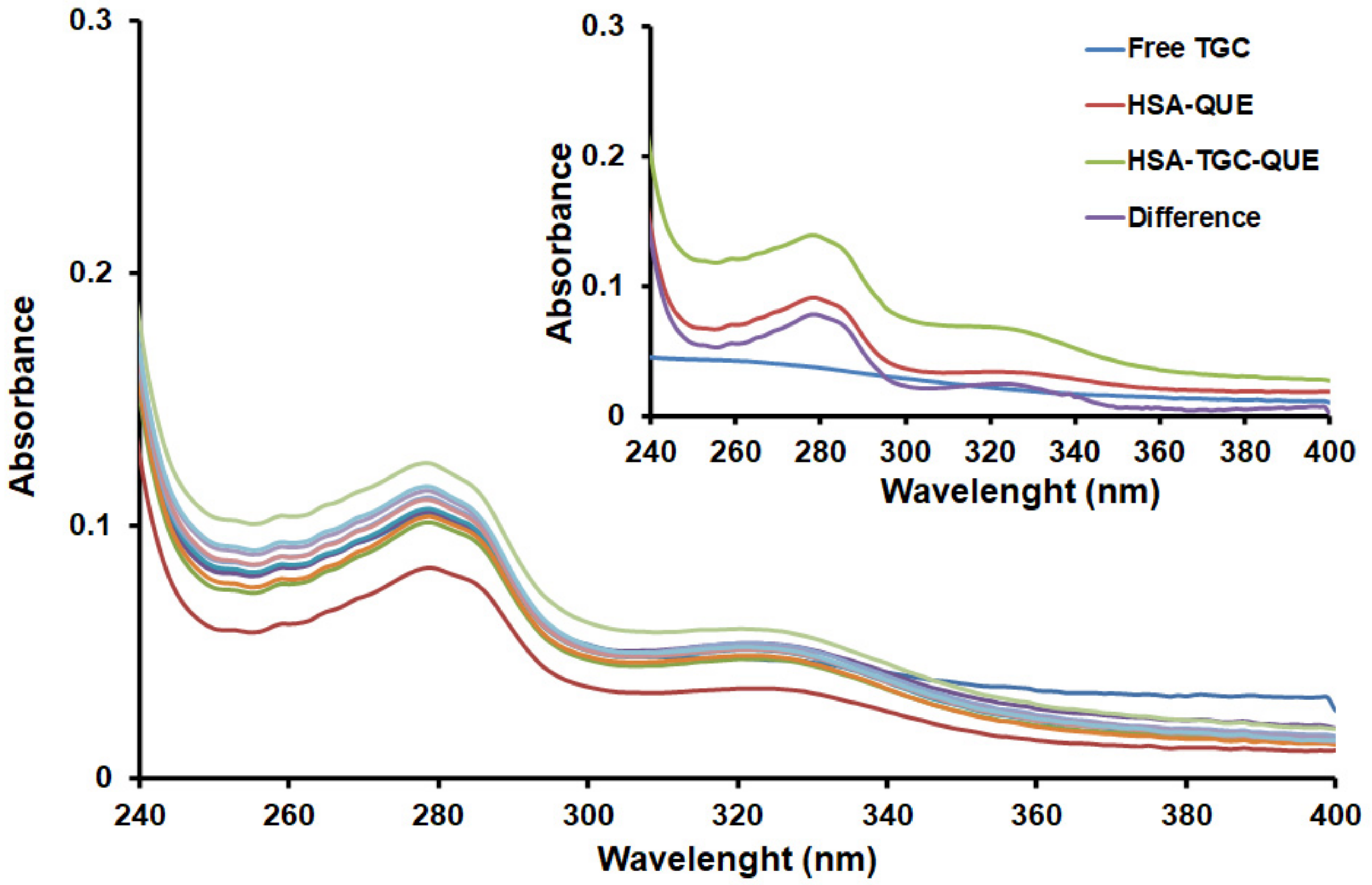
2.1. UV–Vis Absorption Spectra
2.2. Fluorescence Quenching Measurements
2.3. Quenching Mechanism
2.4. Binding Constant and Number of Binding Sites
2.5. Synchronous Fluorescence Spectra
2.6. Circular Dichroism Measurements
2.7. Fourier Transform Infrared Spectroscopy (FT-IR)
2.8. Docking Analysis
3. Materials and Methods
3.1. Materials
3.2. Solutions
3.3. Apparatus
3.3.1. Ultra-Violet Spectroscopy
3.3.2. Fluorescence Spectroscopy
3.3.3. Circular Dichroism Studies
3.3.4. Fourier Transform Infrared Spectroscopy (FT-IR)
3.3.5. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, X.M.; Carter, D.C. Atomic structure and chemistry of human serum albumin. Nature 1992, 358, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Peters, T., Jr. Serum Albumin. Adv. Protein Chem. 1985, 37, 161–245. [Google Scholar] [CrossRef]
- Curry, S.; Brick, P.; Franck, N.P. Fatty acid binding to human serum albumin: New insights from crystallographic studies. Biochim Biophys Acta 1999, 1441, 131–140. [Google Scholar] [CrossRef]
- Yamasaki, K.; Chuang, V.T.G.; Maruyama, T.; Otagiri, M. Albumin–drug interaction and its clinical implication. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 5435–5443. [Google Scholar] [CrossRef]
- Seedher, N.; Agarwal, P. Competitive binding of fluoroquinolone antibiotics and some other drugs to human serum albumin: A luminescence spectroscopic study. Luminescence 2013, 28, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Seedher, N.; Agarwal, P. Complexation of fluoroquinolone antibiotics with human serum albumin: A fluorescence quenching study. J. Lumin. 2010, 130, 1841–1848. [Google Scholar] [CrossRef]
- Kasbekar, N. Tigecycline: A new glycylcycline antimicrobial agent. Am. J. Health Syst. Pharm. 2006, 63, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, S.D.; Janković, S.M.; Matović, Z.D.; Jakovljević, I.Z.; Jelić, R.M. Interaction between tigecycline and human serum albumin in aqueous solution. Monatsh. Chem. 2015, 146, 399–409. [Google Scholar] [CrossRef]
- Daly, J.W.; Butts-Lamb, P.; Padgett, W. Subclasses of adenosine receptors in the central nervous system: Interaction with caffeine and related methylxanthines. Cell. Mol. Neurobiol. 1983, 3, 69–80. [Google Scholar] [CrossRef]
- Gasior, M.; Jaszyna, M.; Peters, J.; Goldberg, S.R. Changes in the Ambulatory Activity and Discriminative Stimulus Effects of Psychostimulant Drugs in Rats Chronically Exposed to Caffeine: Effect of Caffeine Dose. J. Pharmacol. Exp. Ther. 2000, 295, 1101–1111. [Google Scholar] [PubMed]
- Verhoef, P.; Pasman, W.J.; van Vliet, T.; Urgert, R.; Katan, M.B. Contribution of caffeine to the homocysteine-raising effect of coffee: A randomized controlled trial in humans. Am. J. Clin. Nutr. 2002, 76, 1244–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadhav, S.B.; Singhal, R.S.; Gum, L. Laccase–gum Arabic conjugate for preparation of water-soluble oligomer of catechin with enhanced antioxidant activity. Food Chem. 2014, 150, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, C.; Artacho, R.; Giménez, R. Beneficial effects of green tea—A review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef]
- Ishii, T.; Mori, T.; Ichikawa, T.; Kaku, M.; Kusaka, K.; Uekusa, Y.; Akagawa, M.; Aihara, Y.; Furuta, T.; Wakimoto, T.; et al. Human Serum Albumin as an Antioxidant in the Oxidation of (−)-Epigallocatechin Gallate: Participation of Reversible Covalent Binding for Interaction and Stabilization. Bioorg. Med. Chem. 2010, 18, 4892–4896. [Google Scholar] [CrossRef]
- David, A.V.A.; Arulmoli, R.; Parasuraman, S. Overviews of biological importance of quercetin: A bioactive flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Lyseng-Williamson, K.A.; Perry, C.M. Micronised purified flavonoid fraction: A review of its use in chronic venous insufficiency, venous ulcers and haemorrhoids. Drugs 2003, 63, 71–100. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, N.; Vicente, V.; Martínez, C. Synergistic effect of diosmin and interferon-alpha on metastatic pulmonary melanoma. Cancer Biother. Radiopharm. 2009, 24, 347–352. [Google Scholar] [CrossRef]
- Khan, P.; Rahman, S.; Queen, A.; Manzoor, S.; Naz, F.; Hasan, G.M.; Luqman, S.; Kim, J.; Islam, A.; Ahmad, F.; et al. Elucidation of Dietary Polyphenolics as Potential Inhibitor of Microtubule Affinity Regulating Kinase 4: In silico and In vitro Studies. Sci. Rep. 2017, 7, 9470. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Bi, S.; Song, D.; Tian, Y.; Zhou, X.; Liu, Z.; Zhang, H. Molecular spectroscopic study on the interaction of tetracyclines with serum albumins. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2005, 61, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.N.; da Silva, I.M.; Braga, T.C.; de Fátima, Â.; Figueiredo, I.M.; Santos, J.C.C. Thimerosal changes protein conformation and increase the rate of fibrillation in physiological conditions: Spectroscopic studies using bovine serum albumin (BSA). Int. J. Biol. Macromol. 2018, 113, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Al-Lohedan, H.A. Spectroscopic and computational evaluation on the binding of safranal with human serum albumin: Role of inner filter effect in fluorescence spectral correction. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 203, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, S.D.; Nićiforović, J.M.; Živanović, S.M.; Odović, J.V.; Jelić, R.M. Spectroscopic studies on the drug-drug interaction: The influence of fluoroquinolones on the affinity of tigecycline to human serum albumin and identification of the binding site. Monatsh. Chem. 2020, 151, 999–1007. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006; ISBN 978-0-387-46312-4. [Google Scholar]
- Bobone, S.; van de Weert, M.; Stella, L. A reassessment of synchronous fluorescence in the separation of Trp and Tyr contributions in protein emission and in the determination of conformational changes. J. Mol. Struct. 2014, 1077, 68–76. [Google Scholar] [CrossRef]
- Liu, T.; Liu, M.; Guo, Q.; Liu, Y.; Zhao, Y.; Wu, Y.; Sun, B.; Wang, Q.; Liu, J.; Han, J. Investigation of binary and ternary systems of human serum albumin with oxyresveratrol/piceatannol and/or mitoxantrone by multipectroscopy, molecular docking and cytotoxicity evaluation. J. Mol. Liq. 2020, 311, 113364. [Google Scholar] [CrossRef]
- Islam, M.M.; Sonu, V.K.; Gashnga, P.M.; Moyon, N.S.; Mitra, S. Caffeine and sulfadiazine interact differently with human serum albumin: A combined fluorescence and molecular docking study. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 152, 23–33. [Google Scholar] [CrossRef]
- Barreca, D.; Laganà, G.; Bruno, G.; Magazù, S.; Bellocco, E. Diosmin binding to human serum albumin and its preventive action against degradation due to oxidative injuries. Biochimie 2013, 95, 2042–2049. [Google Scholar] [CrossRef]
- Li, X.; Wang, S. Study on the interaction of (+)-catechin with human serum albumin using isothermal titration calorimetry and spectroscopic techniques. New J. Chem. 2015, 39, 386–395. [Google Scholar] [CrossRef]
- Mohseni-Shahri, F.S.; Housaindokht, M.R.; Bozorgmehr, M.R.; Moosavi-Movahedi, A.A. The influence of the flavonoid quercetin on the interaction of propranolol with Human serum albumin: Experimental and theoretical approaches. J. Lumin. 2014, 154, 229–240. [Google Scholar] [CrossRef]
- Berić, J.D.; Stojanović, S.D.; Mrkalić, E.M.; Matović, Z.D.; Milovanović, D.R.; Sovrlić, M.M.; Jelić, R.M. Interaction of haloperidol with human serum albumin and effect of metal ions on the binding. Monatsh. Chem. 2018, 149, 2359–2368. [Google Scholar] [CrossRef]
- Shi, J.H.; Lou, Y.Y.; Zhou, K.L.; Pan, D.Q. Elucidation of intermolecular interaction of bovine serum albumin with Fenhexamid: A biophysical prospect. J. Photochem. Photobiol. B: Biol. 2018, 180, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Shahlaei, M.; Rahimi, B.; Ashrafi-Kooshk, M.R.; Sadrjavadi, K.; Khodarahmi, R. Probing of possible olanzapine binding site on human serum albumin: Combination of spectroscopic methods and molecular dynamics simulation. J. Lumin. 2015, 158, 91–98. [Google Scholar] [CrossRef]
- Liu, B.M.; Zhang, J.; Hao, A.J.; Xu, L.; Wang, D.; Ji, H.; Sun, S.J.; Chen, B.Q.; Liu, B. The increased binding affinity of curcumin with human serum albumin in the presence of rutin and baicalin: A potential for drug delivery system. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 155, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Chen, L.; Yang, F.; Liu, C.; Bai, Y. Green, yellow and red emitting CdTe QDs decreased the affinities of apigenin and luteolin for human serum albumin in vitro. J. Hazard. Mater. 2010, 182, 696–703. [Google Scholar] [CrossRef]
- Brunton, L.L.; Hilal-Dandan, R.; Knollmann, B.C. Goodman & Gilman’s the Pharmacological Basis of Therapeutics; McGraw-Hill Education: New York, NY, USA, 2018; ISBN 1259584739. [Google Scholar]
- Seedher, N.; Agarwal, P. Effect of metal ions on some pharmacologically relevant interactions involving fluoroquinolone antibiotics. Drug Metabol. Drug Interact. 2010, 25, 17–24. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, F.; Chen, L.; Meehan, E.J.; Huang, M. A new drug binding subsite on human serum albumin and drug-drug interaction studied by X-ray crystallography. J. Struct. Biol. 2008, 162, 40–49. [Google Scholar] [CrossRef]
- Maji, A.; Beg, M.; Mandal, A.K.; Das, S.; Jha, P.K.; Kumar, A.; Sarwar, S.; Hossain, M.; Chakrabarti, P. Spectroscopic interaction study of human serum albumin and human hemoglobin with Mersilea quadrifolia leaves extract mediated silver nanoparticles having antibacterial and anticancer activity. J. Mol. Struct. 2017, 1141, 584–592. [Google Scholar] [CrossRef]
- Zhang, G.W.; Que, Q.M.; Pan, J.H.; Guo, J.B. Study of the interaction between icariin and human serum albumin by fluorescence spectroscopy. J. Mol. Struct. 2008, 881, 132–138. [Google Scholar] [CrossRef]
- Shi, J.H.; Zhou, K.L.; Lou, Y.Y.; Pan, D.Q. Multi-spectroscopic and molecular modeling approaches to elucidate the binding interaction between bovine serum albumin and darunavir, a HIV protease inhibitor. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 188, 362–371. [Google Scholar] [CrossRef]
- Matei, I.; Hillebrand, M. Interaction of kaempferol with human serum albumin: A fluorescence and circular dichroism study. J. Pharm. Biomed. Anal. 2010, 51, 768–773. [Google Scholar] [CrossRef]
- Byler, D.M.; Susi, H. Forward, changes in the amide I and amide II bands of HSA on its interaction with the ligand indicate structural changes in the HSA. Biopolymers 1986, 25, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.; Papia, P.C. Ivermectin and Doxycycline Combination as a Promising Drug Candidate Against SARS-CoV-2 Infection: A Computational Study. Physics 2020, arXiv:2012.00653. [Google Scholar]
- Gangopadhyay, A.; Chakraborty, H.J.; Datta, A. Employing virtual screening and molecular dynamics simulations for identifying hits against the active cholera toxin. Toxicon 2019, 170, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Dassault Systèmes BIOVIA. Discovery Studio, v.17.2.0; BIOVIA: San Diego, CA, USA, 2016.
- The PyMOL Molecular Graphics System; Version 2.0; Schrödinger, LLC: New York, NY, USA, 2017; Available online: https://pymol.org/2/ (accessed on 21 December 2020).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System [a] | KSV × 10−4 [b] | kq × 10−12 [c] | R2 [d] | Ka × 10−5 [b] | n | R2 |
|---|---|---|---|---|---|---|
| HSA-TGC | 5.00 ± 0.03 | 5.00 | 0.996 | 0.18 ± 0.03 | 0.9 | 0.991 |
| HSA-TGC-CAF | 1.87 ± 0.04 | 1.87 | 0.977 | 0.573 ± 0.06 | 1.09 | 0.985 |
| HSA-TGC-CAT | 2.57 ± 0.06 | 2.57 | 0.989 | 1.71 ± 0.07 | 1.17 | 0.983 |
| HSA-TGC-QUE | 3.96 ± 0.05 | 3.96 | 0.987 | 22.30 ± 0.02 | 1.35 | 0.996 |
| HSA-TGC-DIO | 2.47 ± 0.05 | 2.47 | 0.986 | 1.98 ± 0.08 | 1.17 | 0.982 |
| System [b] | Autodock | Ki[c] | Ref. | |
|---|---|---|---|---|
| Site I | Site II | |||
| HSA-TGC | −24.36 | −14.60 | 5.44 × 10−5 | [8] |
| HSA-TGC-CAF | −24.89 | −15.35 | 4.33 × 10−5 | This work |
| HSA-TGC-CAT | −25.60 | −16.65 | 3.27 × 10−5 | |
| HSA-TGC-QUE | −30.75 | −18.78 | 4.13 × 10−6 | |
| HSA-TGC-DIO | −26.40 | −17.36 | 2.37 × 10−5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sovrlić, M.; Mrkalić, E.; Jelić, R.; Ćendić Serafinović, M.; Stojanović, S.; Prodanović, N.; Tomović, J. Effect of Caffeine and Flavonoids on the Binding of Tigecycline to Human Serum Albumin: A Spectroscopic Study and Molecular Docking. Pharmaceuticals 2022, 15, 266. https://doi.org/10.3390/ph15030266
Sovrlić M, Mrkalić E, Jelić R, Ćendić Serafinović M, Stojanović S, Prodanović N, Tomović J. Effect of Caffeine and Flavonoids on the Binding of Tigecycline to Human Serum Albumin: A Spectroscopic Study and Molecular Docking. Pharmaceuticals. 2022; 15(3):266. https://doi.org/10.3390/ph15030266
Chicago/Turabian StyleSovrlić, Miroslav, Emina Mrkalić, Ratomir Jelić, Marina Ćendić Serafinović, Stefan Stojanović, Nevena Prodanović, and Jovica Tomović. 2022. "Effect of Caffeine and Flavonoids on the Binding of Tigecycline to Human Serum Albumin: A Spectroscopic Study and Molecular Docking" Pharmaceuticals 15, no. 3: 266. https://doi.org/10.3390/ph15030266
APA StyleSovrlić, M., Mrkalić, E., Jelić, R., Ćendić Serafinović, M., Stojanović, S., Prodanović, N., & Tomović, J. (2022). Effect of Caffeine and Flavonoids on the Binding of Tigecycline to Human Serum Albumin: A Spectroscopic Study and Molecular Docking. Pharmaceuticals, 15(3), 266. https://doi.org/10.3390/ph15030266