
Evening Primrose Oil Improves Chemotherapeutic Effects in Human Pancreatic Ductal Adenocarcinoma Cell Lines—A Preclinical Study


 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
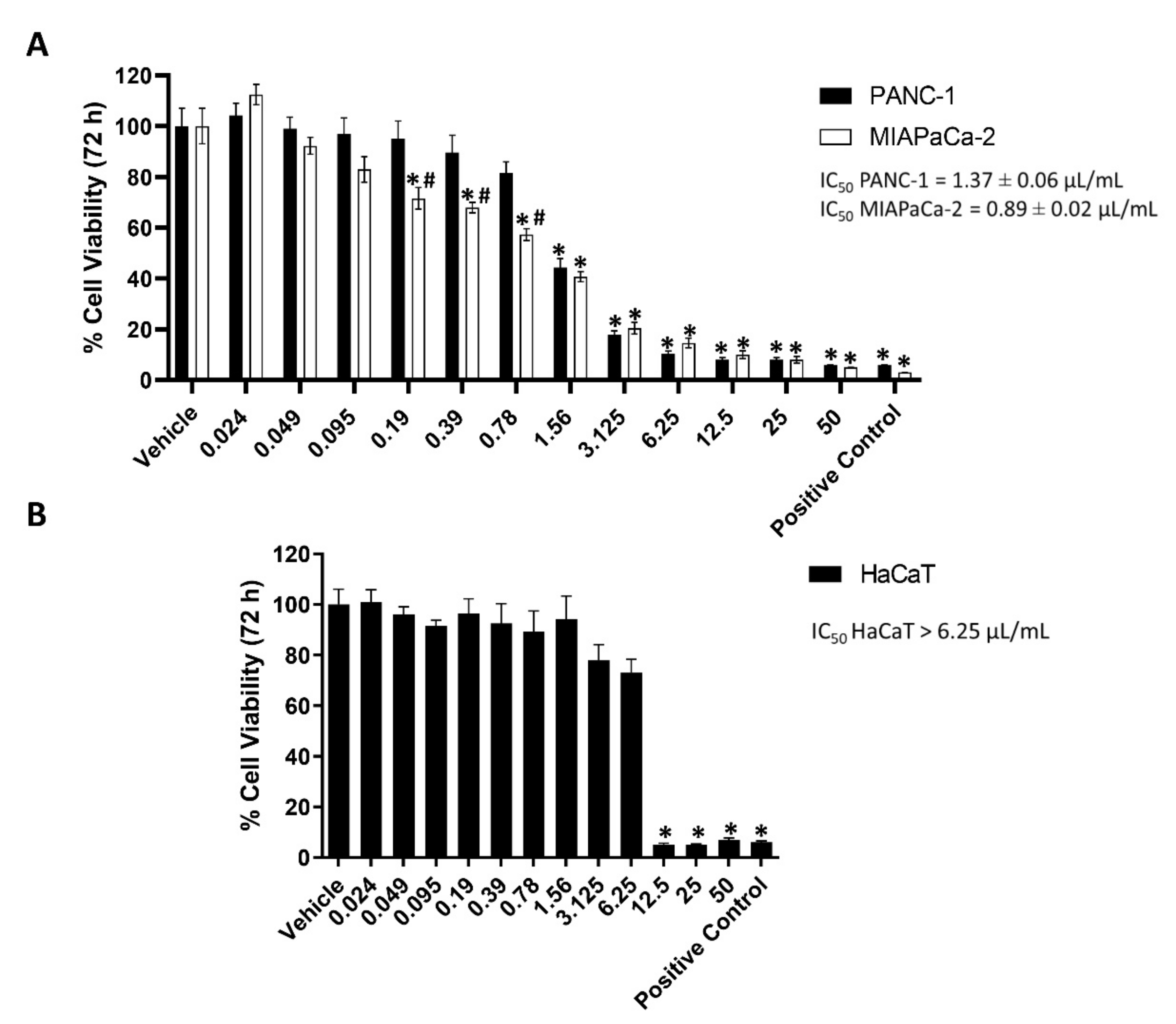
2.1. EPO Reduces PANC-1 and MIAPaCa-2 Cell Viability
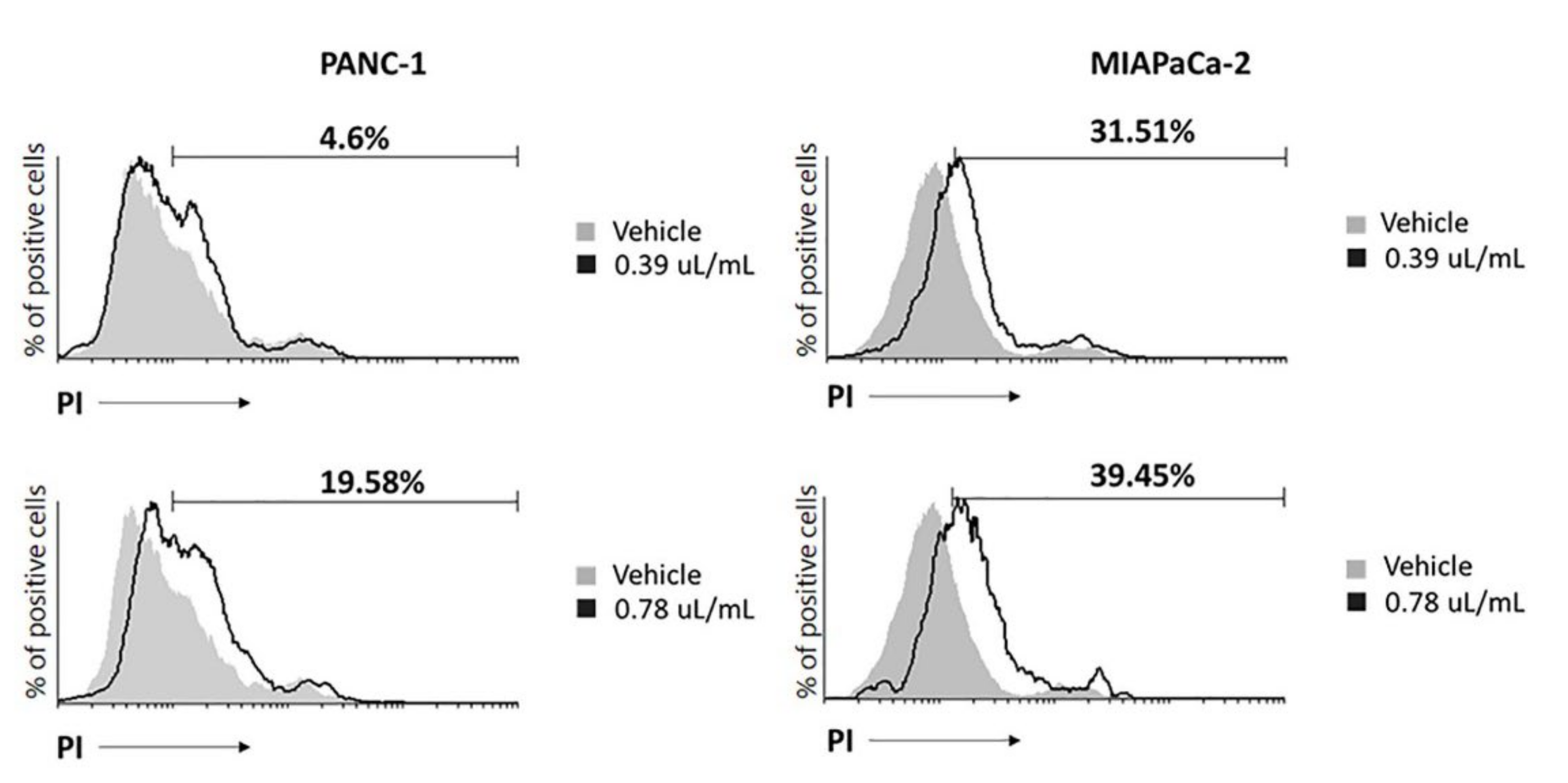
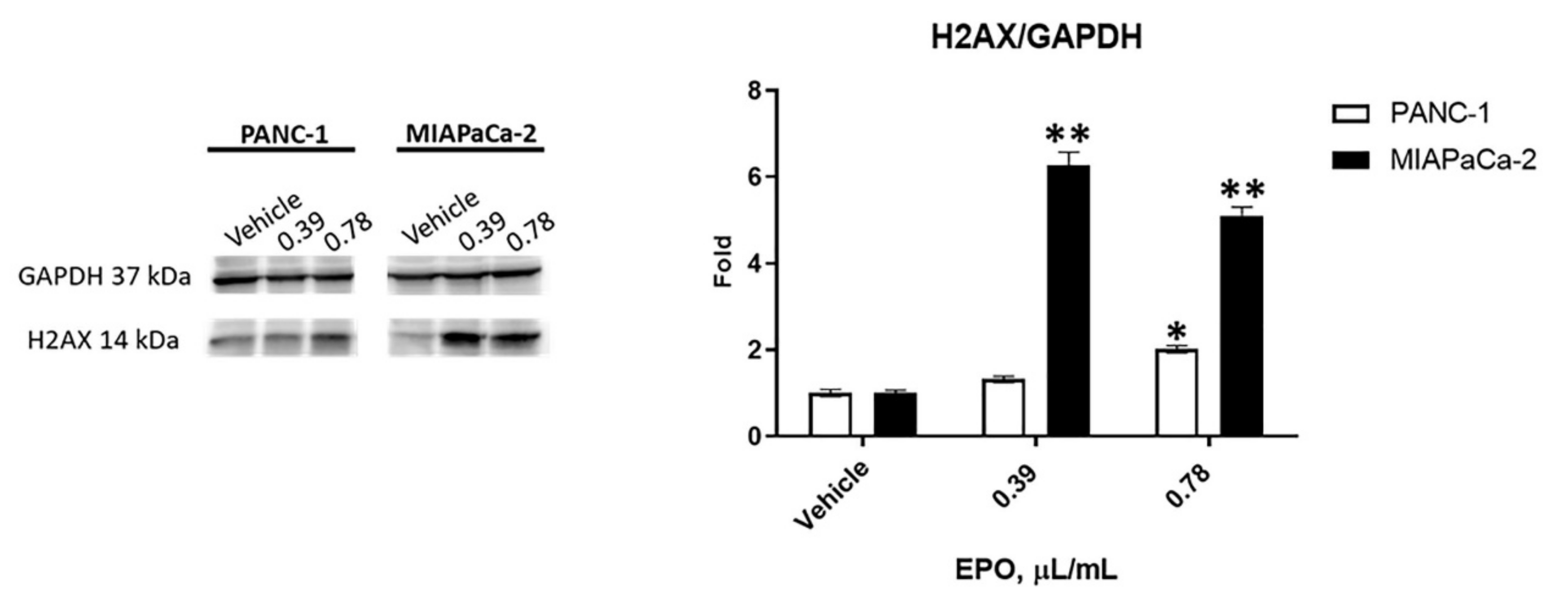
2.2. EPO Induces Cell Death in PANC-1 and MIAPaCa-2 Cell Lines
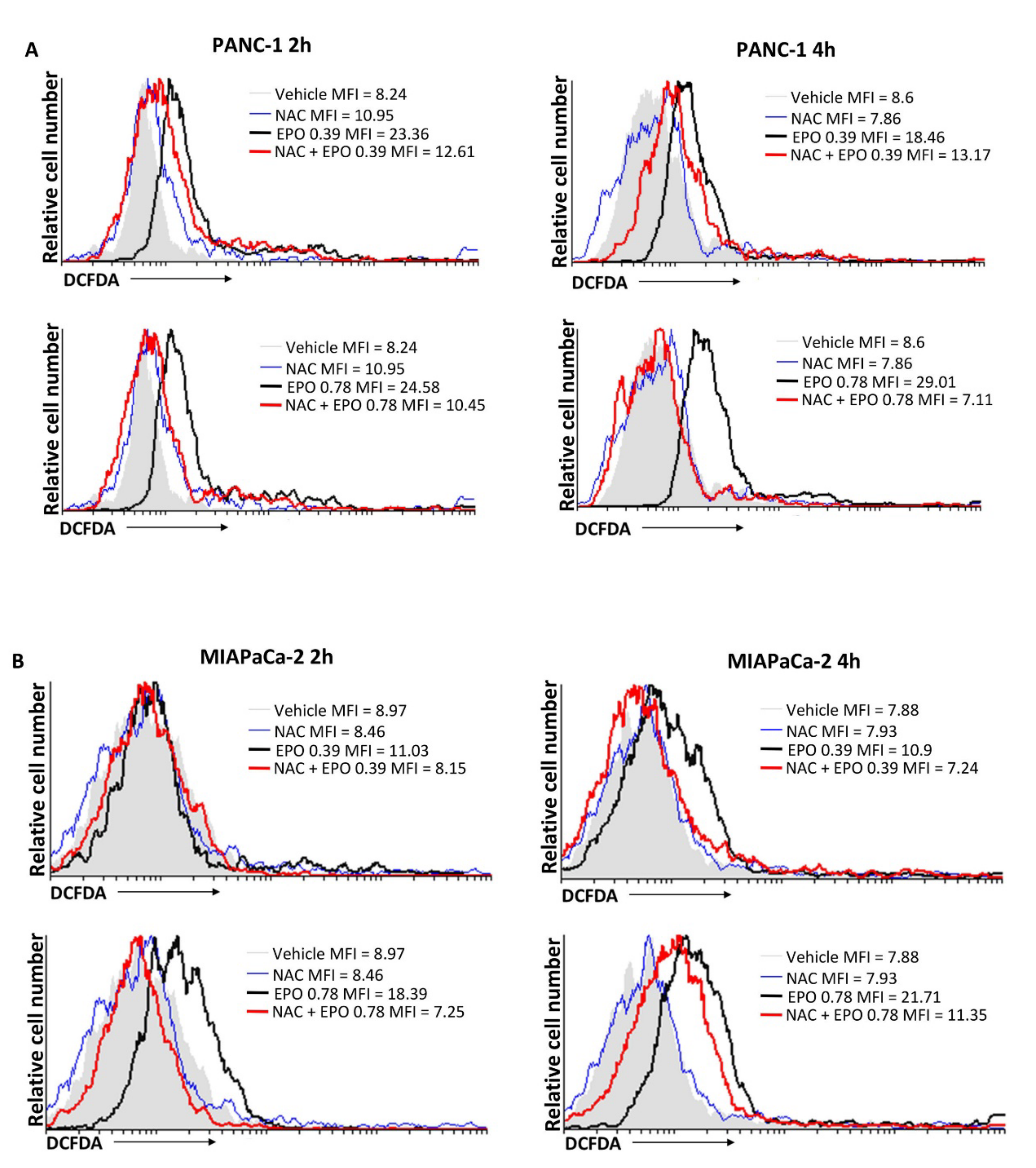
2.3. EPO Induces ROS Production in PANC-1 and MIAPaCa-2 Cell Lines
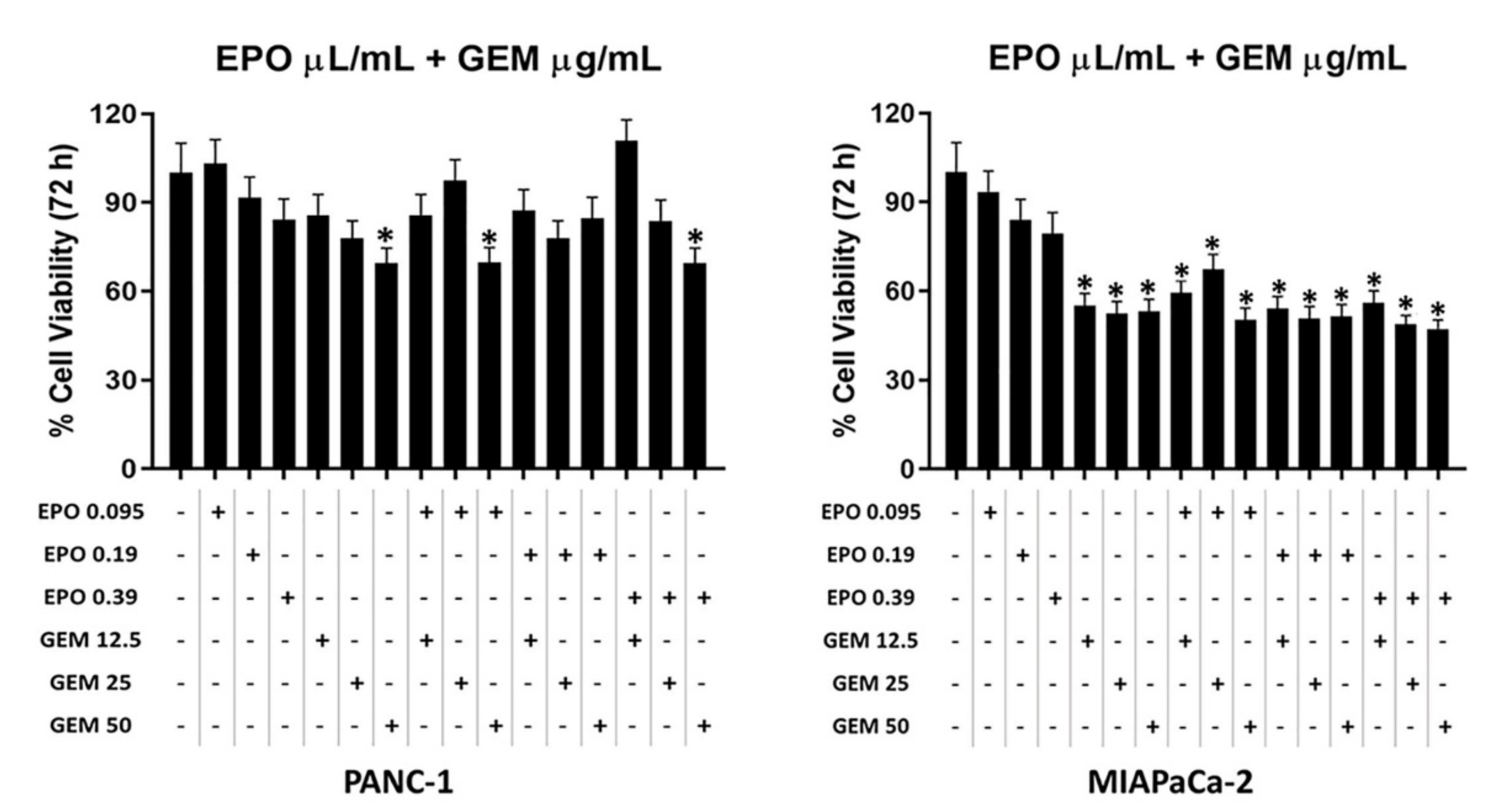
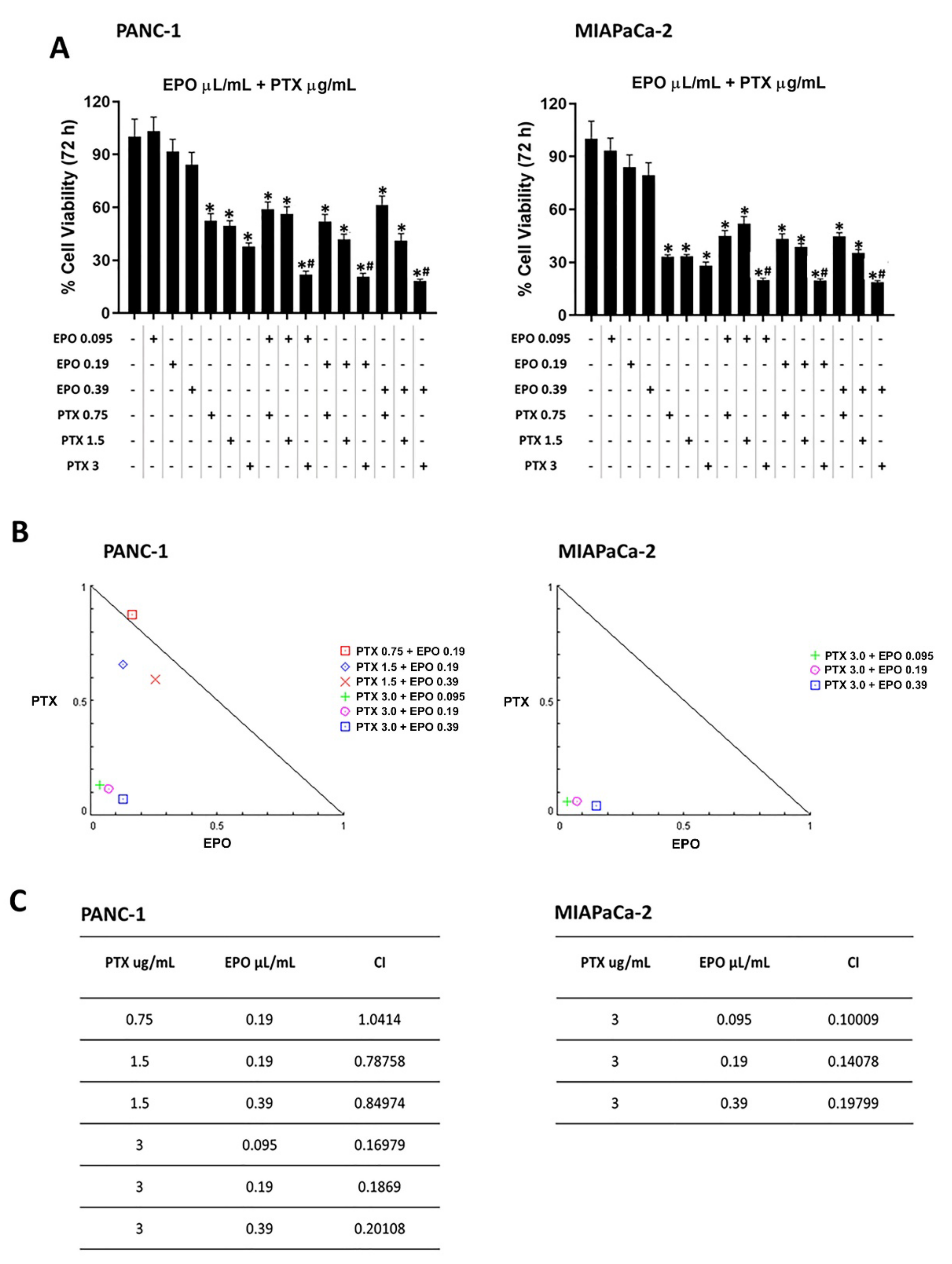
2.4. EPO Potentiates Paclitaxel Efficacy in PANC-1 and MIAPaCa-2 Cell Lines
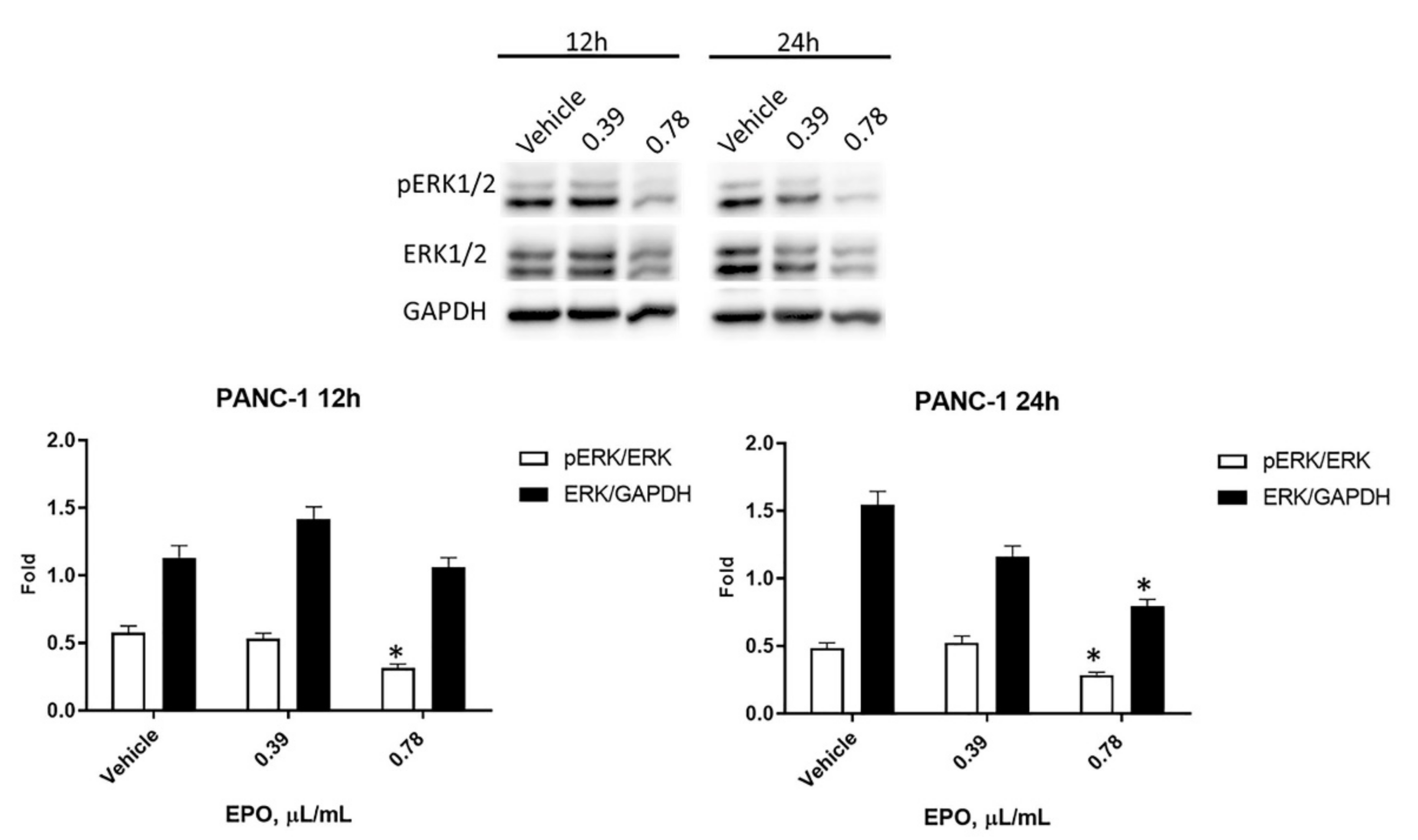
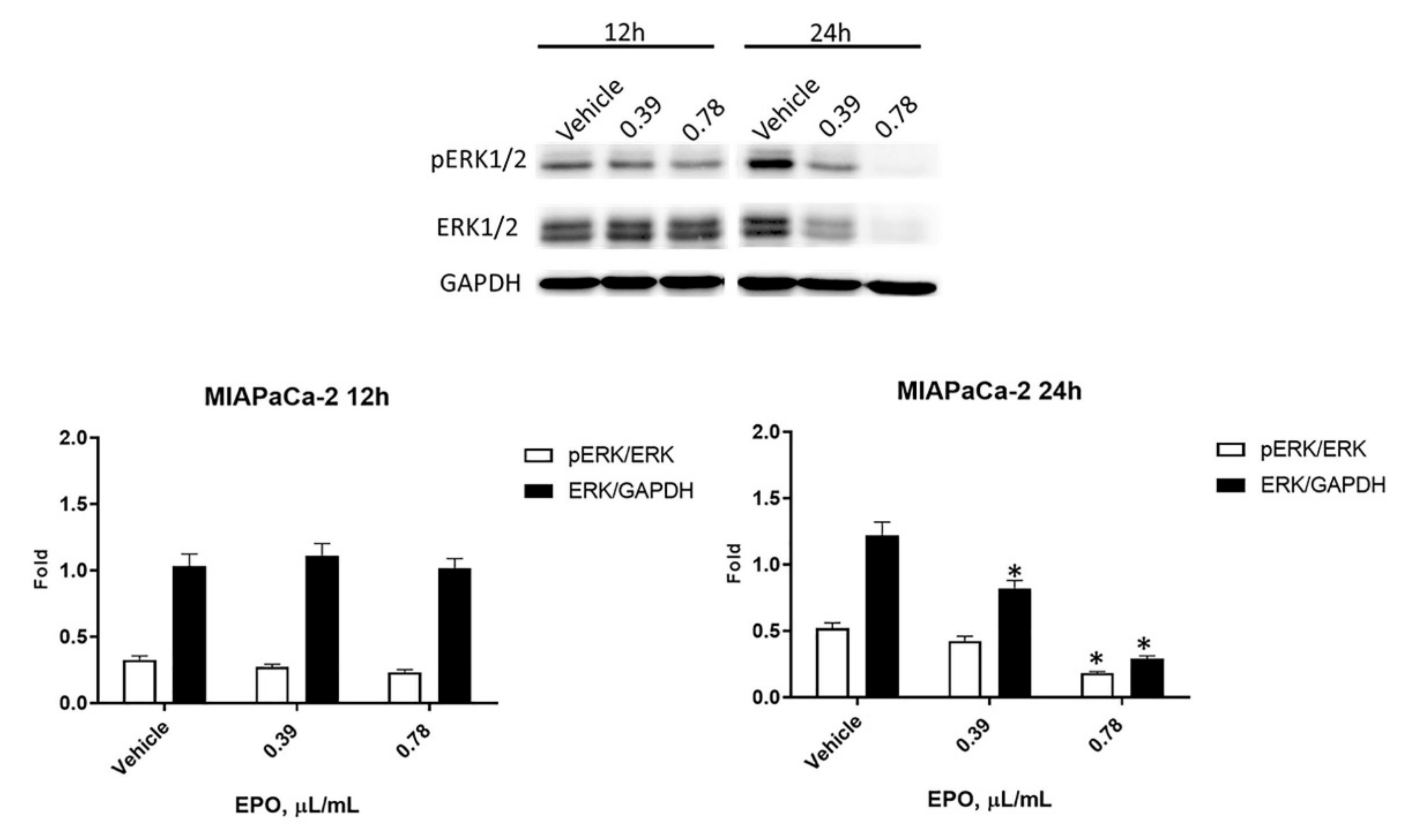
2.5. EPO Influences pERK/ERK Protein Levels and Activation
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Reagents
4.3. MTT Assay
4.4. Cell Death Assay
4.5. Western Blot Analyses
4.6. Reactive Oxygen Species (ROS) Production
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bojková, B.; Winklewski, P.J.; Wszedybyl-Winklewska, M. Dietary Fat and Cancer-Which Is Good, Which Is Bad, and the Body of Evidence. Int. J. Mol. Sci. 2020, 21, 4114. [Google Scholar] [CrossRef] [PubMed]
- Lal, S.; Shekher, A.; Puneet; Narula, A.S.; Abrahamse, H.; Gupta, S.C. Cannabis and its constituents for cancer: History, biogenesis, chemistry and pharmacological activities. Pharmacol. Res. 2021, 163, 105302. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, O.; Morelli, M.B.; Annibali, D.; Aguzzi, C.; Zeppa, L.; Tuyaerts, S.; Amantini, C.; Amant, F.; Ferretti, B.; Maggi, F.; et al. The Effects of Cannabidiol and Prognostic Role of TRPV2 in Human Endometrial Cancer. Int. J. Mol. Sci. 2020, 21, 5409. [Google Scholar] [CrossRef] [PubMed]
- Luongo, M.; Marinelli, O.; Zeppa, L.; Aguzzi, C.; Morelli, M.B.; Amantini, C.; Frassineti, A.; Di Costanzo, M.; Fanelli, A.; Santoni, G.; et al. Cannabidiol and Oxygen-Ozone Combination Induce Cytotoxicity in Human Pancreatic Ductal Adenocarcinoma Cell Lines. Cancers 2020, 12, 2774. [Google Scholar] [CrossRef]
- Brunetti, A.; Marinelli, O.; Morelli, M.B.; Iannarelli, R.; Amantini, C.; Russotti, D.; Santoni, G.; Maggi, F.; Nabissi, M. Isofuranodiene synergizes with temozolomide in inducing glioma cells death. Phytomedicine 2019, 52, 51–59. [Google Scholar] [CrossRef]
- Vernieri, C.; Nichetti, F.; Raimondi, A.; Pusceddu, S.; Platania, M.; Berrino, F.; de Braud, F. Diet and supplements in cancer prevention and treatment: Clinical evidences and future perspectives. Crit. Rev. Oncol. Hematol. 2018, 123, 57–73. [Google Scholar] [CrossRef]
- Velentzis, L.S.; Keshtgar, M.R.; Woodside, J.V.; Leathem, A.J.; Titcomb, A.; Perkins, K.A.; Mazurowska, M.; Anderson, V.; Wardell, K.; Cantwell, M.M. Significant changes in dietary intake and supplement use after breast cancer diagnosis in a UK multicentre study. Breast Cancer Res. Treat. 2011, 128, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Timoszuk, M.; Bielawska, K.; Skrzydlewska, E. Evening Primrose (Oenothera biennis) Biological Activity Dependent on Chemical Composition. Antioxidants 2018, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-De La Paz, S.; Fernández-Arche, M.A.; Ángel-Martín, M.; García-Giménez, M.D. Phytochemical characterization of potential nutraceutical ingredients from Evening Primrose oil (Oenothera biennis L.). Phytochem. Lett. 2014, 8, 158–162. [Google Scholar] [CrossRef]
- Montserrat-De La Paz, S.; Fernández-Arche, Á.; Ángel-Martín, M.; García-Giménez, M.D. The sterols isolated from Evening Primrose oil modulate the release of proinflammatory mediators. Phytomedicine 2012, 19, 1072–1076. [Google Scholar] [CrossRef]
- Luchini, C.; Capelli, P.; Scarpa, A. Pancreatic Ductal Adenocarcinoma and Its Variants. Surg. Pathol. Clin. 2016, 9, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic Ductal Adenocarcinoma: Current and Evolving Therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef] [PubMed]
- Ansari, D.; Tingstedt, B.; Andersson, B.; Holmquist, F.; Sturesson, C.; Williamsson, C.; Sasor, A.; Borg, D.; Bauden, M.; Andersson, R. Pancreatic cancer: Yesterday, today and tomorrow. Future Oncol. 2016, 12, 1929–1946. [Google Scholar] [CrossRef] [Green Version]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, N.; Monahan, S.; Stefaniak, A.; Schwarz, M.A.; Schwarz, R.E. Inhibition of the MEK/ERK pathway augments nab-paclitaxel-based chemotherapy effects in preclinical models of pancreatic cancer. Oncotarget 2017, 9, 5274–5286. [Google Scholar] [CrossRef] [Green Version]
- Chmielewska-Kassassir, M.; Sobierajska, K.; Ciszewski, W.M.; Bukowiecka-Matusiak, M.; Szczesna, D.; Burzynska-Pedziwiatr, I.; Wiczkowski, W.; Wagner, W.; Wozniak, L.A. Polyphenol Extract from Evening Primrose ( Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells. Biomolecules 2020, 10, 1574. [Google Scholar] [CrossRef]
- Whitehouse, P.A.; Cooper, A.J.; Johnson, C.D. Synergistic activity of gamma-linolenic acid and cytotoxic drugs against pancreatic adenocarcinoma cell lines. Pancreatology 2003, 3, 367–374. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Y.; Gao, D.; Yang, L.; Qian, S.Y. Dihomo-γ-linolenic acid inhibits growth of xenograft tumors in mice bearing human pancreatic cancer cells (BxPC-3) transfected with delta-5-desaturase shRNA. Redox Biol. 2019, 20, 236–246. [Google Scholar] [CrossRef]
- Khodeer, D.M.; Mehanna, E.T.; Abushouk, A.I.; Abdel-daim, M.M. Protective Effects of Evening Primrose Oil against Cyclophosphamide-Induced Biochemical, Histopathological, and Genotoxic Alterations in Mice. Pathogens 2020, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Arimura, T.; Kojima-Yuasa, A.; Suzuki, M.; Kennedy, D.O.; Matsui-Yuasa, I. Caspase-independent apoptosis induced by evening primrose extract in Ehrlich ascites tumor cells. Cancer Lett. 2003, 201, 9–16. [Google Scholar] [CrossRef]
- Auberger, J.; Vogt, S.; Hopfinger, G.; Clausen, J.; Greil, R. Topical evening primrose oil for reduction of bortezomib-induced skin reactions. Ann. Hematol. 2013, 92, 995–996. [Google Scholar] [CrossRef] [PubMed]
- Gorlach, S.; Wagner, W.; Podsȩdek, A.; Sosnowska, D.; Dastych, J.; Koziolkiewicz, M. Polyphenols from evening primrose (Oenothera paradoxa) defatted seeds induce apoptosis in human colon cancer Caco-2 cells. J. Agric. Food Chem. 2011, 59, 6985–6997. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-de la Paz, S.; Fernández-Arche, M.A.; Bermúdez, B.; García-Giménez, M.D. The sterols isolated from evening primrose oil inhibit human colon adenocarcinoma cell proliferation and induce cell cycle arrest through upregulation of LXR. J. Funct. Foods 2015, 12, 64–69. [Google Scholar] [CrossRef] [Green Version]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. ROS-Mediated Cancer Cell Killing through Dietary Phytochemicals. Oxid. Med. Cell. Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Lu, C.C.; Yang, J.S.; Huang, A.C.; Hsia, T.C.; Chou, S.T.; Kuo, C.L.; Lu, H.F.; Lee, T.H.; Wood, W.G.; Chung, J.G. Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Mol. Nutr. Food Res. 2010, 54, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Samatar, A.A.; Poulikakos, P.I. Targeting RAS-ERK signalling in cancer: Promises and challenges. Nat. Rev. Drug Discov. 2014, 13, 928–942. [Google Scholar] [CrossRef]
- Lanfredini, S.; Thapa, A.; O’Neill, E. RAS in pancreatic cancer. Biochem. Soc. Trans. 2019, 47, 961–972. [Google Scholar] [CrossRef]
- Serini, S.; Calviello, G. Modulation of Ras/ERK and Phosphoinositide Signaling by Long-Chain n-3 PUFA in Breast Cancer and Their Potential Complementary Role in Combination with Targeted Drugs. Nutrients 2017, 9, 185. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Methyl Esters (FAME) | USP Comp % |
|---|---|
| Methyl palmitate | 6.00 |
| Methyl stearate | 1.91 |
| Methyl oleate | 6.93 |
| Methyl linoleate | 74.18 |
| Methyl-y-linolenate | 9.98 |
| Methyl-o-linolenate | 0.22 |
| Methyl arachidate | 0.49 |
| Methyl eicosenoate | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeppa, L.; Aguzzi, C.; Versari, G.; Luongo, M.; Morelli, M.B.; Maggi, F.; Amantini, C.; Santoni, G.; Marinelli, O.; Nabissi, M. Evening Primrose Oil Improves Chemotherapeutic Effects in Human Pancreatic Ductal Adenocarcinoma Cell Lines—A Preclinical Study. Pharmaceuticals 2022, 15, 466. https://doi.org/10.3390/ph15040466
Zeppa L, Aguzzi C, Versari G, Luongo M, Morelli MB, Maggi F, Amantini C, Santoni G, Marinelli O, Nabissi M. Evening Primrose Oil Improves Chemotherapeutic Effects in Human Pancreatic Ductal Adenocarcinoma Cell Lines—A Preclinical Study. Pharmaceuticals. 2022; 15(4):466. https://doi.org/10.3390/ph15040466
Chicago/Turabian StyleZeppa, Laura, Cristina Aguzzi, Giorgia Versari, Margherita Luongo, Maria Beatrice Morelli, Federica Maggi, Consuelo Amantini, Giorgio Santoni, Oliviero Marinelli, and Massimo Nabissi. 2022. "Evening Primrose Oil Improves Chemotherapeutic Effects in Human Pancreatic Ductal Adenocarcinoma Cell Lines—A Preclinical Study" Pharmaceuticals 15, no. 4: 466. https://doi.org/10.3390/ph15040466
APA StyleZeppa, L., Aguzzi, C., Versari, G., Luongo, M., Morelli, M. B., Maggi, F., Amantini, C., Santoni, G., Marinelli, O., & Nabissi, M. (2022). Evening Primrose Oil Improves Chemotherapeutic Effects in Human Pancreatic Ductal Adenocarcinoma Cell Lines—A Preclinical Study. Pharmaceuticals, 15(4), 466. https://doi.org/10.3390/ph15040466