
Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
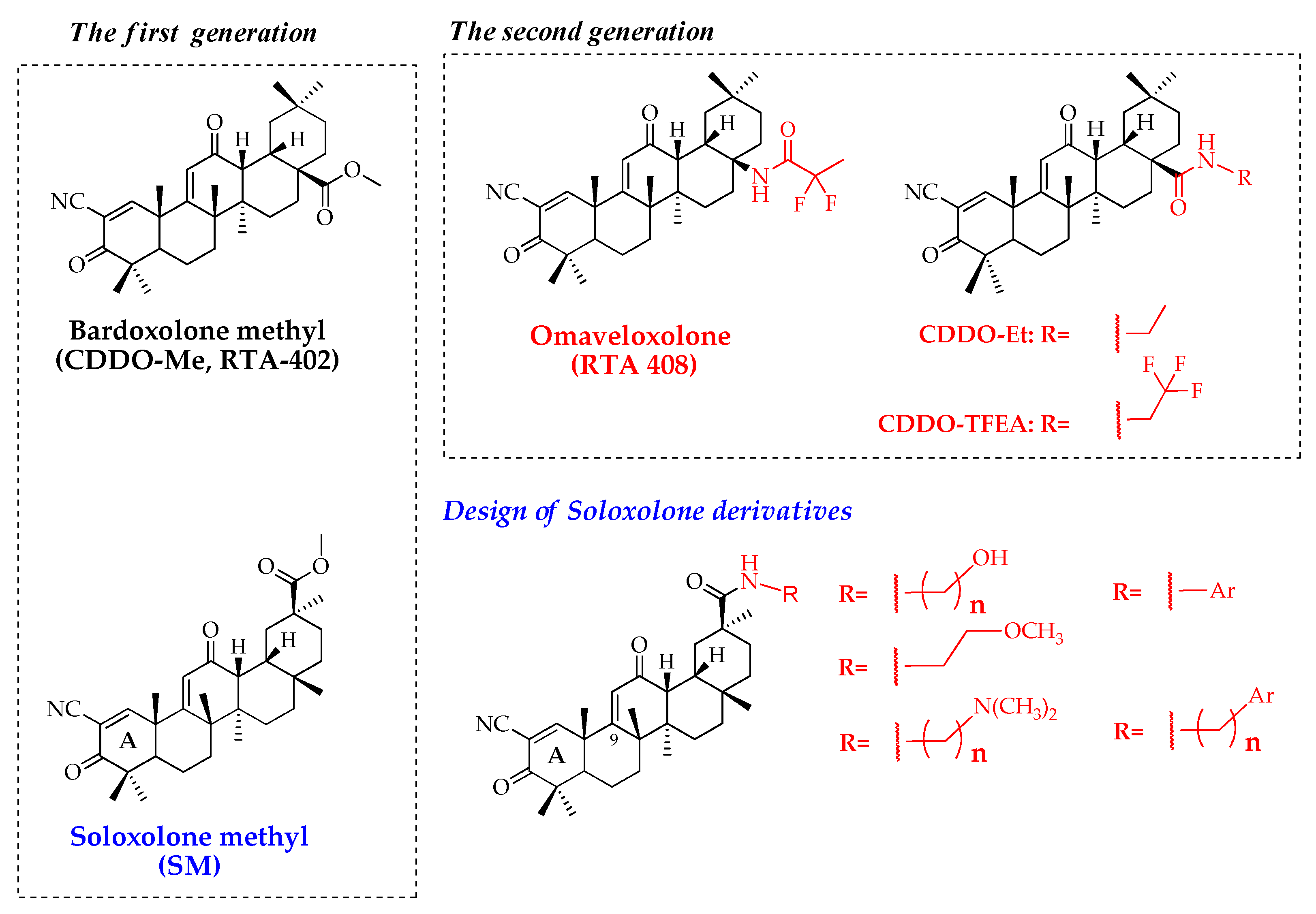
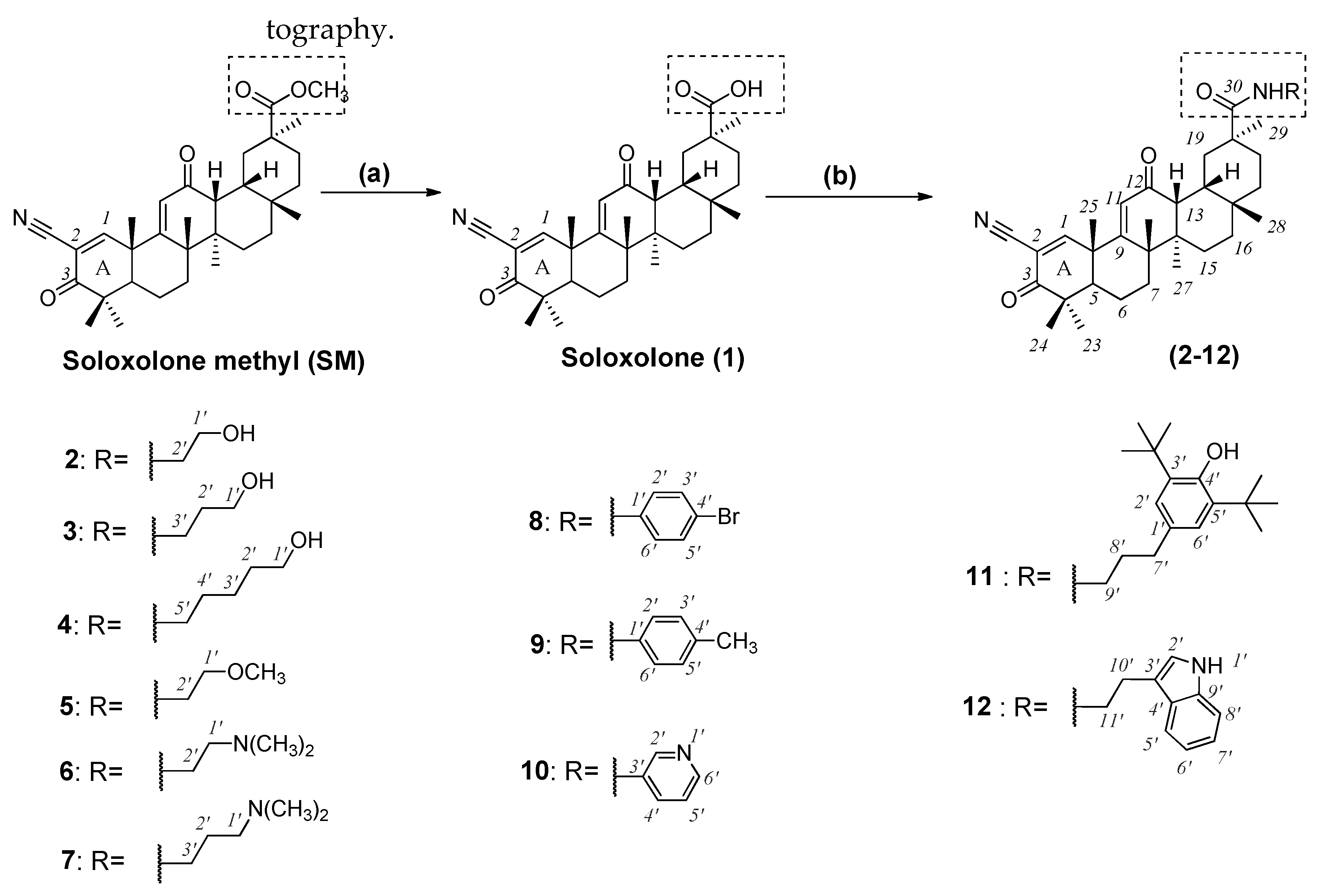
2.1. Chemistry
2.2. Biological Evaluation
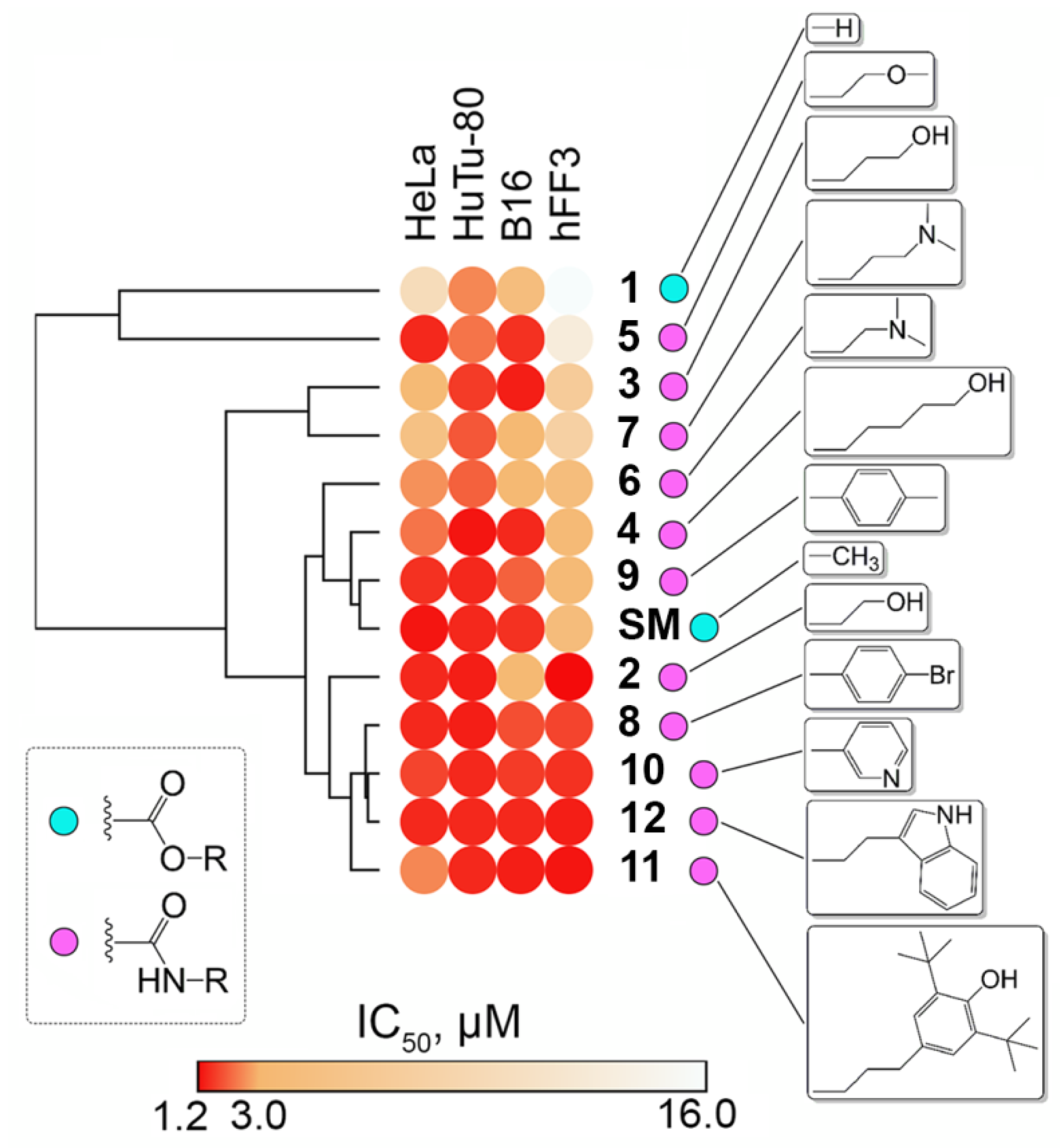
2.2.1. Cytotoxicity of Novel Compounds
- The presence of an additional N,N-dimethylamine-containing group in the side chain of soloxolone amides led to a decrease in the cytotoxicity of the derivatives compared to SM in a panel of tumor cells (average IC506 = 3.0 µM and IC507 = 4.5 µM versus IC50SM = 2.1 µM).
- Hydroxyamides with C-2 and C-5 linkers (compounds 2 and 4, respectively) had similar cytotoxic activity compared to SM (average IC502 = 1.9 µM and IC504 = 2.2 µM versus IC50SM = 2.1 µM in a panel of tumor cells); the cytotoxicity of hydroxyamide with a C-3 linker was somewhat lower (average IC503 = 3.4 µM).
- The introduction of aryl-containing moieties into the side chain of soloxolone amides had virtually no effect on the toxicity of derivatives against tumor cells (average IC508−11 = 1.5–1.8 µM versus IC50SM = 1.5 µM); however, it slightly increased their cytotoxicity against fibroblasts (average IC508,10−12(hFF3) = 1.5 µM versus IC50SM (hFF3) = 4 µM);
- In the case of hFF3 fibroblasts, compounds 3 and 7 with a C-3 linker between functional groups, both in the series of hydroxyamides (compounds 2–4) and dimethylaminoamides (compounds 6–7), respectively, had a somewhat lower toxicity compared to their C-2/C-5 linker-containing counterparts (IC503 = 6.8 µM versus IC502 = 1.2 µM (C-2 linker) or IC504 = 3.6 µM (C-5 linker); IC507 = 7.7 µM versus IC506= 4.1 µM (C-2 linker)). Additionally, the replacement of hydroxy or dimethylamino groups with a methoxy moiety was desirable for soloxolone amides with a C-2 linker (IC505 = 12.8 µM versus IC502 = 1.2 µM and IC506 = 4.1 µM).
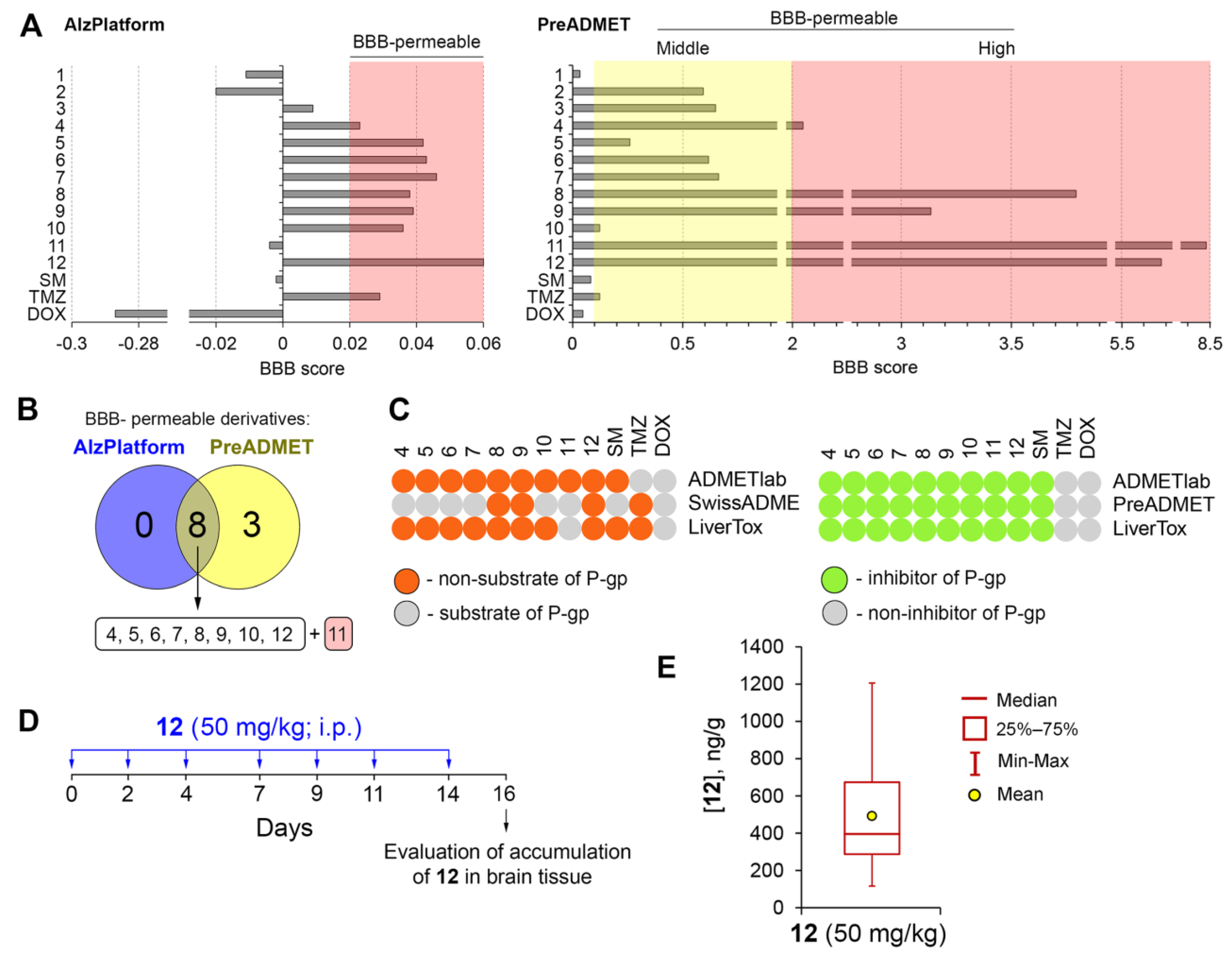
2.2.2. Analysis of the BBB Permeability of the Novel Compounds In Silico
2.2.3. Verification of BBB Permeability of Soloxolone Tryptamide (12) In Vivo
2.2.4. Cytotoxicity of Novel Soloxolone Amides (4–12) against Brain Tumor Cells
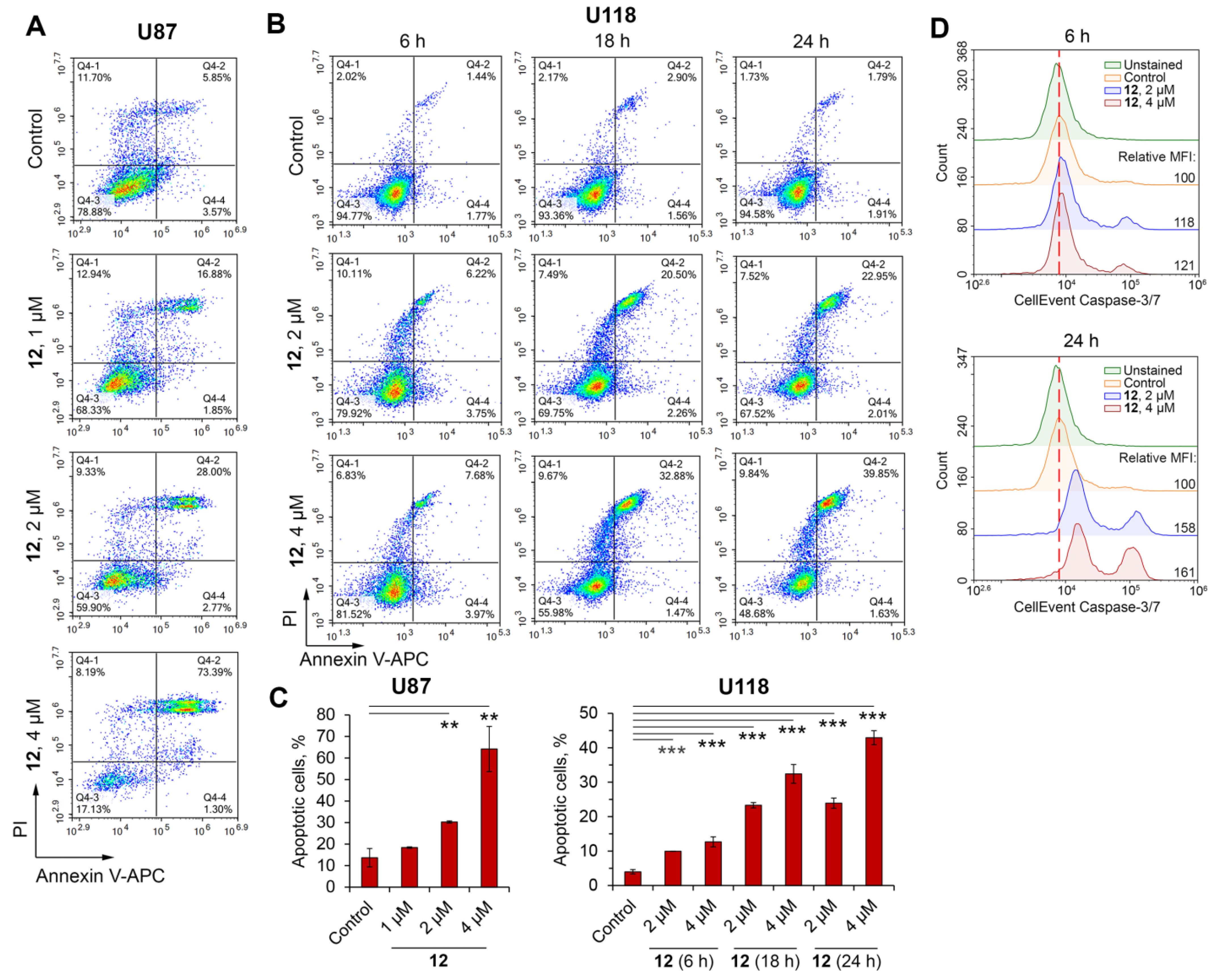
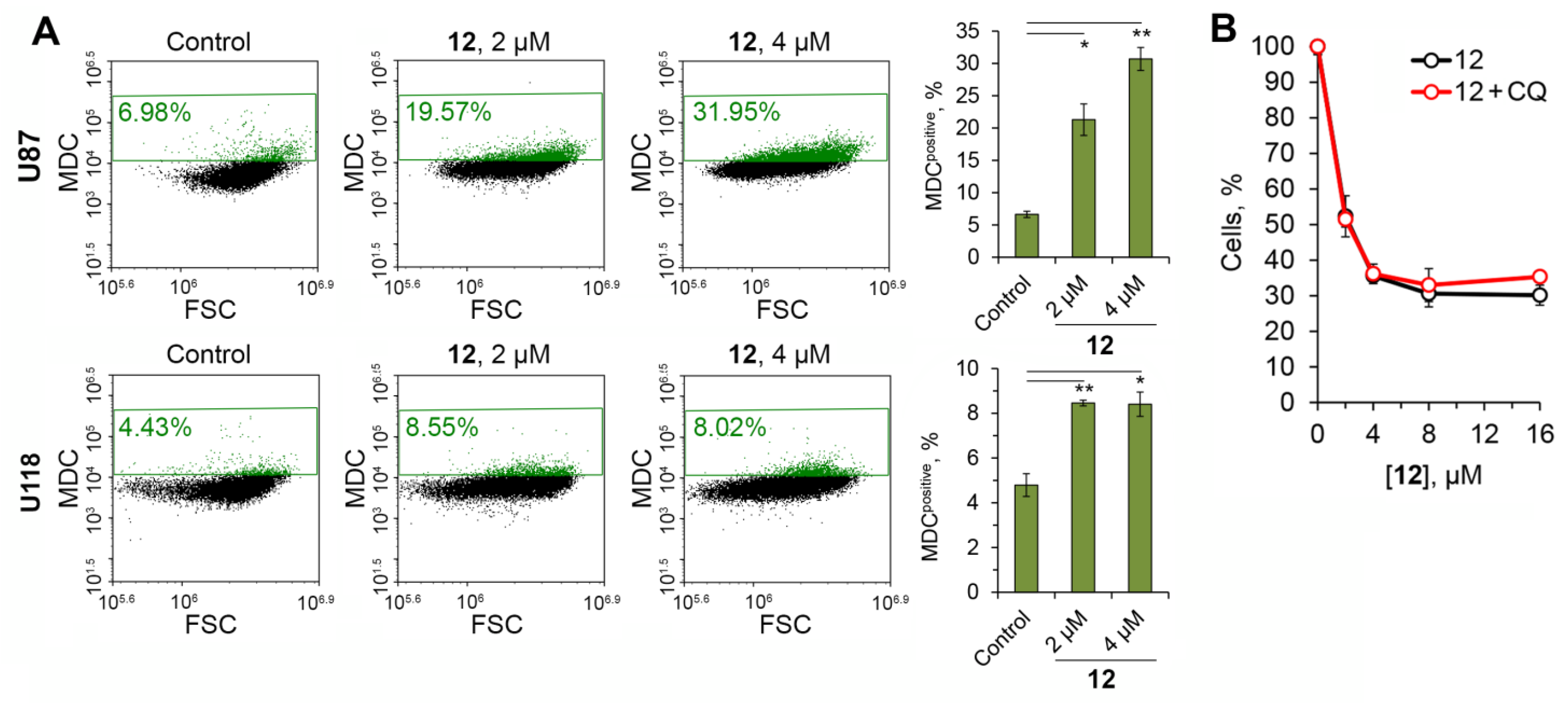
2.2.5. Soloxolone Tryptamide 12 Induced Apoptosis-Dependent and Autophagy-Independent Death of Glioblastoma Cells
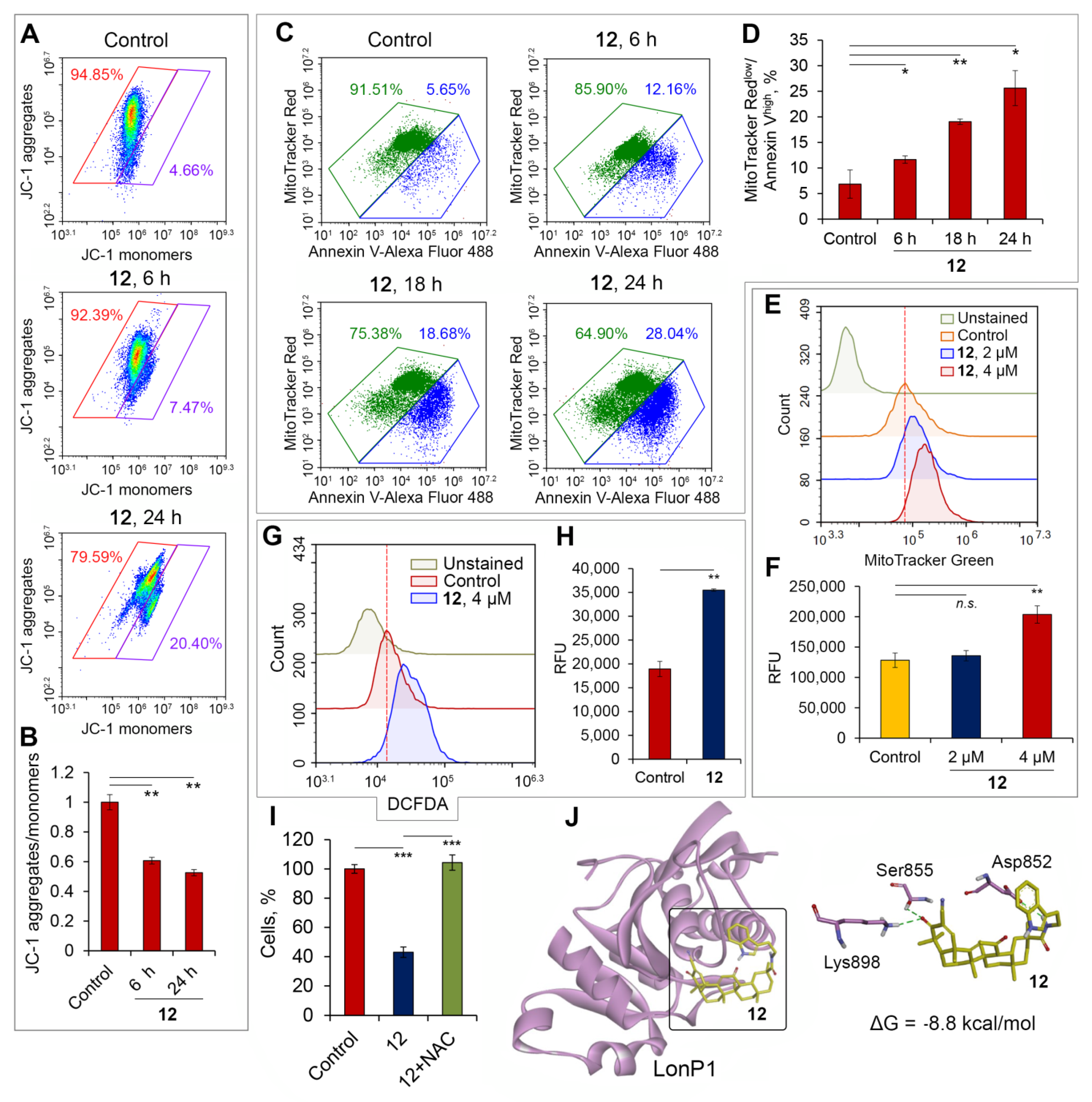
2.2.6. Soloxolone Tryptamide 12 Induced ROS-Dependent Death of Glioblastoma Cells by Disruption of Mitochondrial Homeostasis
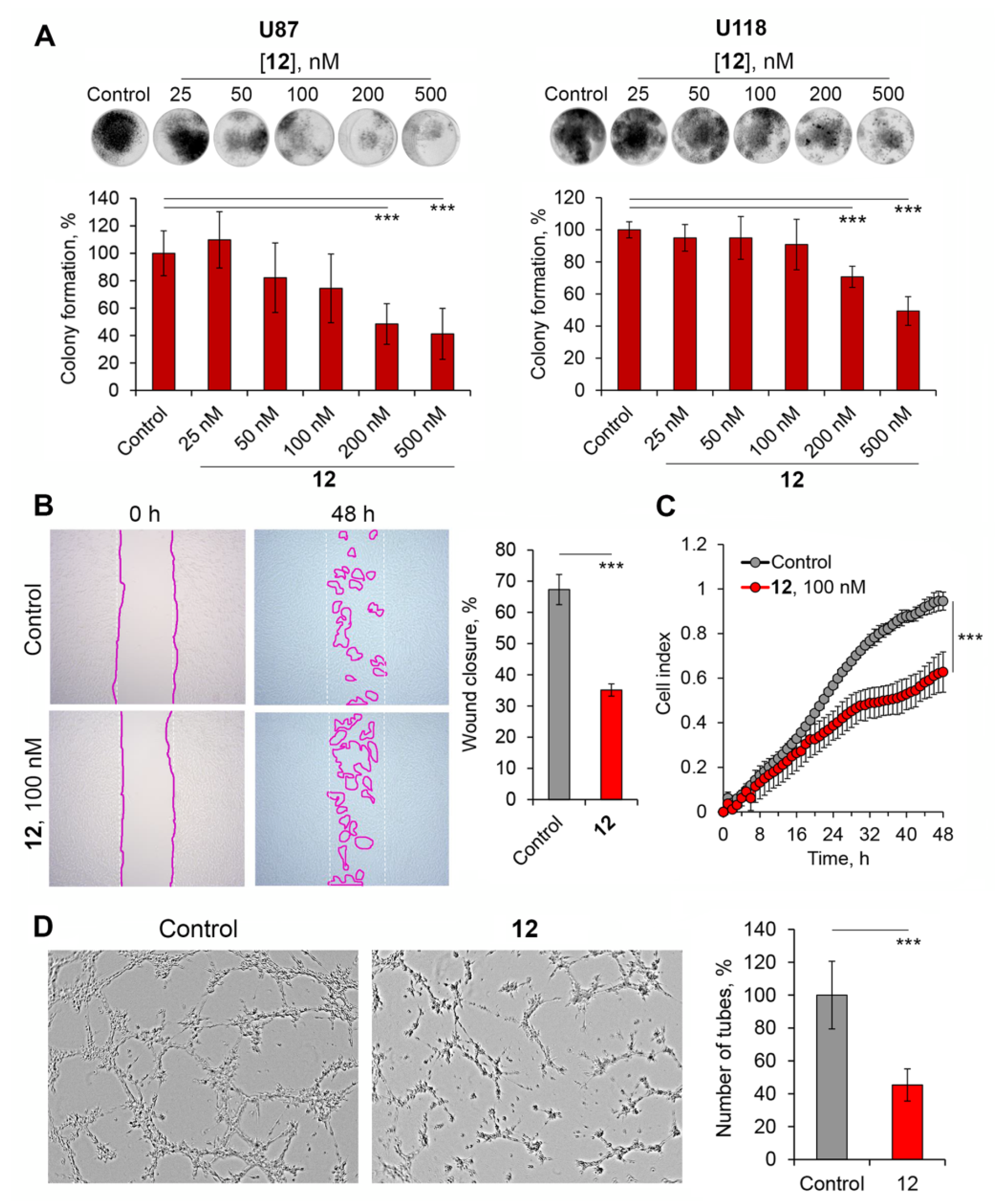
2.2.7. Soloxolone Tryptamide 12 Displayed Significant Antitumor Potential against Glioblastoma Cells at Non-Toxic Concentrations
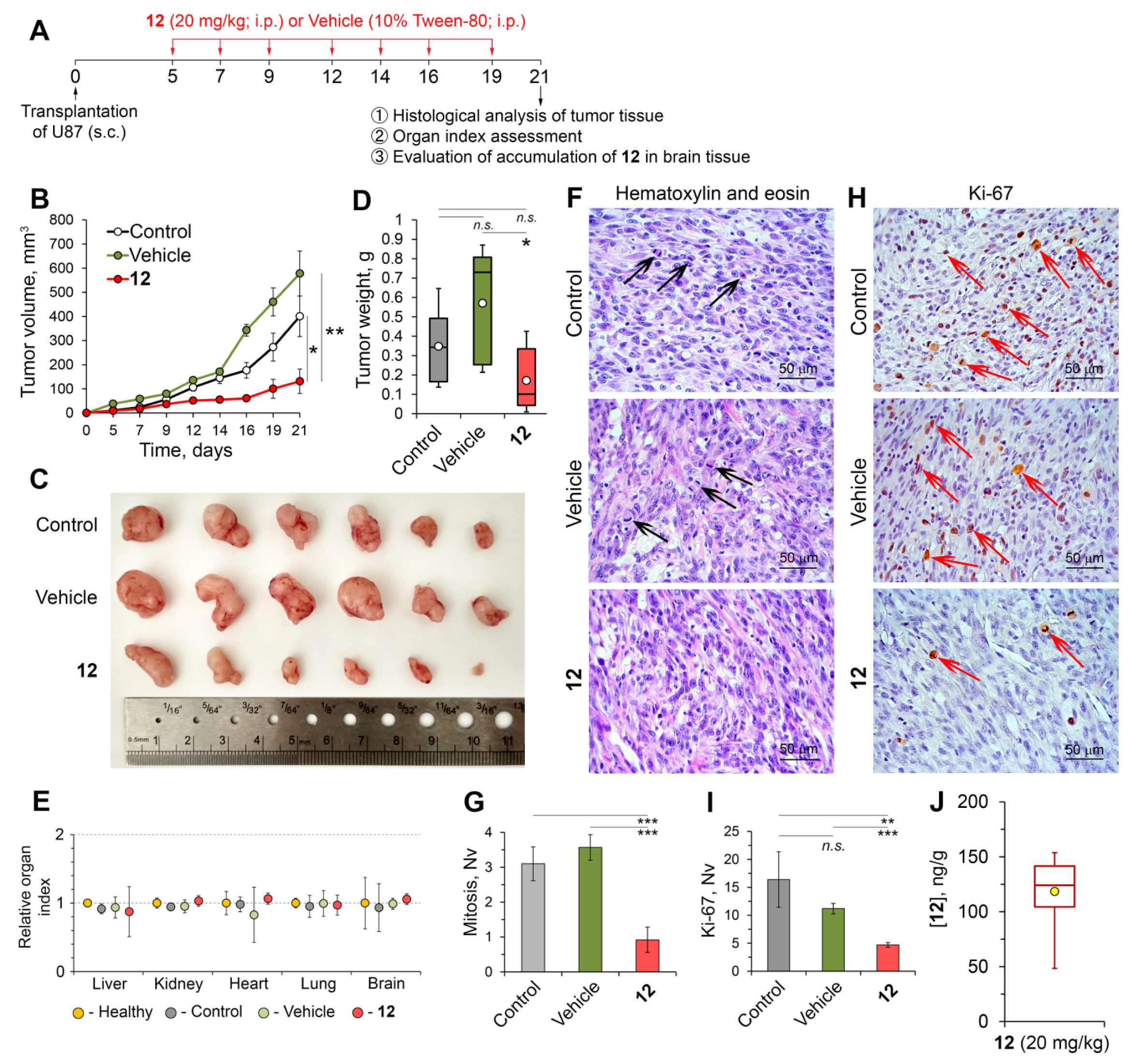
2.2.8. Compound 12 Displayed High Anti-Glioblastoma Potential In Vivo
Soloxolone Tryptamide 12 Inhibited the Growth of U87 Glioblastoma in a Murine Xenograft Model
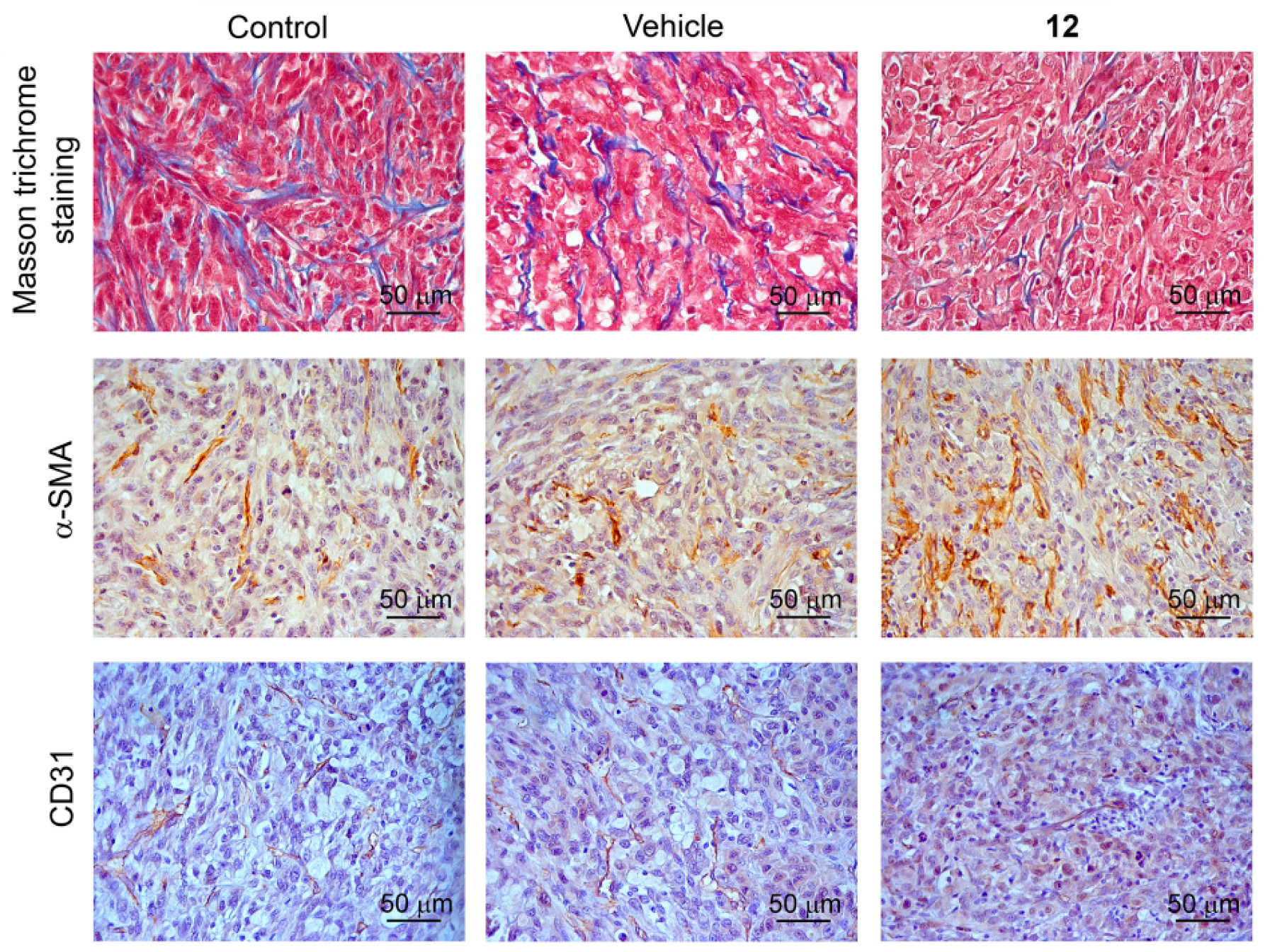
Compound 12 Improved the Tumor Microenvironment in a Subcutaneous U87 Glioblastoma Xenograft Model
3. Materials and Methods
3.1. Chemistry
3.1.1. General Experimental Procedures and Reagents
General Procedure A for Compounds (2–5, 8–12)
General Procedure B for Compounds (6–7)
3.1.2. 2-Cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-oic acid (Soloxolone) (1)
3.1.3. N-(2′-Hydroxyethyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (2)
3.1.4. N-(3′-Hydroxypropyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (3)
3.1.5. N-(5′-Hydroxypentyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (4)
3.1.6. N-(2′-Methoxyethyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (5)
3.1.7. N-(2′-(Dimethylamino)ethyl)-2-Cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (6)
3.1.8. N-(3′-(Dimethylamino)propyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (7)
3.1.9. N-(4′-Bromophenyl)-2-Cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (8)
3.1.10. N-p-Tolyl-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (9)
3.1.11. N-(Pyridin-3-yl)-2-Cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (10)
3.1.12. N-(3′-(3′’,5′’-di-tert-butyl-4′’-hydroxyphenyl)propyl)-2-Cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30- amide (11)
3.1.13. N-(2′-(1H-Indol-2-yl)-ethyl)-2-cyano-3,12-dioxo-18βH-olean-9(11),1(2)-dien-30-amide (12)
3.2. Biological Evaluations
3.2.1. Cell lines and Novel Compounds
3.2.2. Mice
3.2.3. Evaluation of Cytotoxicity of Novel Compounds by MTT Assay
3.2.4. In Silico Prediction of the Blood–Brain Barrier (BBB) Permeability of the Novel Compounds and Their P-glycoprotein (P-gp) Substrate Specificity
3.2.5. Evaluation of Brain Accumulation of 12
3.2.6. Working Solutions for Analysis of Brain Accumulation of 12
3.2.7. Brain Tissue Homogenization Protocol
3.2.8. Preparation of Calibrators and Experimental Samples for Quantification of 12 in Mouse Brain Tissues
3.2.9. HPLC–MS/MS Conditions
3.2.10. Analysis of Pro-Apoptotic Activity of 12
3.2.11. Analysis of Caspase-3/-7 Activation
3.2.12. Analysis of Autophagy-Inducing Activity of 12
3.2.13. Analysis of Mitochondrial Membrane Potential
3.2.14. Evaluation of Mitochondrial Mass
3.2.15. Analysis of ROS Production
3.2.16. Molecular Docking
3.2.17. Colony Formation Assay
3.2.18. Scratch Assay
3.2.19. Evaluation of Trans-Well Motility of Tumor Cells Using the xCELLigence Platform
3.2.20. Vasculogenic Mimicry
3.2.21. Tumor Transplantation and Design of Animal Experiments
3.2.22. Toxicity Assessment
3.2.23. Histology and Immunohistochemistry
3.2.24. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liby, K.T.; Sporn, M.B. Synthetic Oleanane Triterpenoids: Multifunctional Drugs with a Broad Range of Applications for Prevention and Treatment of Chronic Disease. Pharmacol. Rev. 2012, 64, 972–1003. [Google Scholar] [CrossRef] [Green Version]
- An Extended Access Program for Bardoxolone Methyl in Patients with CKD. Available online: https://clinicaltrials.gov/ct2/show/NCT03749447 (accessed on 18 March 2022).
- Extended Access Program to Assess Long-term Safety of Bardoxolone Methyl in Patients with Pulmonary Hypertension. Available online: https://clinicaltrials.gov/ct2/show/NCT03068130 (accessed on 18 March 2022).
- Chertow, G.M.; Appel, G.B.; Andreoli, S.; Bangalore, S.; Block, G.A.; Chapman, A.B.; Chin, M.P.; Gibson, K.L.; Goldsberry, A.; Iijima, K.; et al. Study Design and Baseline Characteristics of the CARDINAL Trial: A Phase 3 Study of Bardoxolone Methyl in Patients with Alport Syndrome. Am. J. Nephrol. 2021, 52, 180–189. [Google Scholar] [CrossRef]
- Salvador, J.A.R.; Leal, A.S.; Alho, D.P.S.; Gonçalves, B.M.F.; Valdeira, A.S.; Mendes, V.I.S.; Jing, Y. Highlights of Pentacyclic Triterpenoids in the Cancer Settings. Stud. Nat. Prod. Chem. 2014, 41, 33–73. [Google Scholar] [CrossRef]
- Salvador, J.A.R.; Leal, A.S.; Valdeira, A.S.; Gonçalves, B.M.F.; Alho, D.P.S.; Figueiredo, S.A.C.; Silvestre, S.M.; Mendes, V.I.S. Oleanane-, ursane-, and quinone methide friedelane-type triterpenoid derivatives: Recent advances in cancer treatment. Eur. J. Med. Chem. 2017, 142, 95–130. [Google Scholar] [CrossRef]
- Honda, T.; Honda, Y.; Favaloro, F.G.; Gribble, G.W.; Suh, N.; Place, A.E.; Rendi, M.H.; Sporn, M.B. A novel dicyanotriterpenoid, 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-onitrile, active at picomolar concentrations for inhibition of nitric oxide production. Bioorg. Med. Chem. Lett. 2002, 12, 1027–1030. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, H.; Huang, J.; Yao, Z.; Yu, J.; Zhang, W.; Zhang, L.; Wang, Z.; Zhuang, C. Discovery of bardoxolone derivatives as novel orally active necroptosis inhibitors. Eur. J. Med. Chem. 2021, 212, 113030. [Google Scholar] [CrossRef]
- Cao, M.; Onyango, E.O.; Williams, C.R.; Royce, D.B.; Gribble, G.W.; Sporn, M.B.; Liby, K.T. Novel synthetic pyridyl analogues of CDDO-Imidazolide are useful new tools in cancer prevention. Pharmacol. Res. 2015, 100, 135–147. [Google Scholar] [CrossRef]
- Yang, L.; Calingasan, N.Y.; Thomas, B.; Charturvedi, R.K.; Kiaei, M.; Wille, E.J.; Liby, K.T.; Williams, C.; Royce, D.; Risingson, R.; et al. Neuroprotective effects of the triterpenoid, CDDO methyl amide, a potent inducer of Nrf2-mediated transcription. PLoS ONE 2009, 4, e5757. [Google Scholar] [CrossRef] [Green Version]
- Neymotin, A.; Calingasan, N.Y.; Wille, E.; Naseri, N.; Petri, S.; Damiano, M.; Liby, K.T.; Risingsong, R.; Sporn, M.; Beal, M.F.; et al. Neuroprotective effect of Nrf2/ARE activators, CDDO ethylamide and CDDO trifluoroethylamide, in a mouse model of amyotrophic lateral sclerosis. Free Radic. Biol. Med. 2011, 51, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Reisman, S.A.; Lee, C.Y.I.; Meyer, C.J.; Proksch, J.W.; Ward, K.W. Topical application of the synthetic triterpenoid RTA 408 activates Nrf2 and induces cytoprotective genes in rat skin. Arch. Dermatol. Res. 2014, 306, 447–454. [Google Scholar] [CrossRef]
- Reisman, S.A.; Lee, C.Y.I.; Meyer, C.J.; Proksch, J.W.; Sonis, S.T.; Ward, K.W. Topical Application of the Synthetic Triterpenoid RTA 408 Protects Mice from Radiation-Induced Dermatitis. Radiat. Res. 2014, 181, 512–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probst, B.L.; Trevino, I.; McCauley, L.; Bumeister, R.; Dulubova, I.; Wigley, W.C.; Ferguson, D.A. RTA 408, a novel synthetic triterpenoid with broad anticancer and anti-inflammatory activity. PLoS ONE 2015, 10, e0122942. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.R.; Chin, M.P.; Delatycki, M.B.; Subramony, S.H.; Corti, M.; Hoyle, J.C.; Boesch, S.; Nachbauer, W.; Mariotti, C.; Mathews, K.D.; et al. Safety and Efficacy of Omaveloxolone in Friedreich Ataxia (MOXIe Study). Ann. Neurol. 2021, 89, 212–225. [Google Scholar] [CrossRef] [PubMed]
- RTA 408 Capsules in Patients with Mitochondrial Myopathy. Available online: https://clinicaltrials.gov/ct2/show/NCT02255422 (accessed on 18 March 2022).
- RTA 408 Ophthalmic Suspension for the Prevention of Corneal Endothelial Cell Loss following Cataract Surgery. Available online: https://clinicaltrials.gov/ct2/show/NCT02128113 (accessed on 19 March 2022).
- Reisman, S.A.; Gahir, S.S.; Lee, C.Y.I.; Proksch, J.W.; Sakamoto, M.; Ward, K.W. Pharmacokinetics and pharmacodynamics of the novel Nrf2 activator omaveloxolone in primates. Drug Des. Devel. Ther. 2019, 13, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.-Y.; Chou, Y.-Y.; Ciou, J.-W.; Liu, F.-Y.; Huang, S.-P.; Chien, J.-Y.; Chou, Y.-Y.; Ciou, J.-W.; Liu, F.-Y.; Huang, S.-P.; et al. The Effects of Two Nrf2 Activators, Bardoxolone Methyl and Omaveloxolone, on Retinal Ganglion Cell Survival during Ischemic Optic Neuropathy. Antioxidants 2021, 10, 1466. [Google Scholar] [CrossRef]
- Tsai, T.H.; Lin, S.H.; Wu, C.H.; Tsai, Y.C.; Yang, S.F.; Lin, C.L. Mechanisms and therapeutic implications of RTA 408, an activator of Nrf2, in subarachnoid hemorrhage–induced delayed cerebral vasospasm and secondary brain injury. PLoS ONE 2020, 15, e0240122. [Google Scholar] [CrossRef]
- Rogers, L.J.; John, T.; Park, J.; Tucker, M.; Ma, H.; Wu, Y.; Rayalam, S.; Wang, X. Growth inhibition and apoptosis of human multiple myeloma cells induced by 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid derivatives. Anticancer. Drugs 2020, 31, 806–818. [Google Scholar] [CrossRef]
- Alabran, J.L.; Cheuk, A.; Liby, K.; Sporn, M.; Khan, J.; Letterio, J.; Leskov, K.S. Human neuroblastoma cells rapidly enter cell cycle arrest and apoptosis following exposure to C-28 derivatives of the synthetic triterpenoid CDDO. Cancer Biol. Ther. 2008, 7, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Nowak, A.J.; Cohen, A.J.; Correia, E.D.; Portocarrero, C.P.; South, A.P.; Nikbakht, N. Omaveloxolone attenuates squamous cell carcinoma growth and disease severity in an Epidermolysis Bullosa mouse model. Exp. Dermatol. 2022. [Google Scholar] [CrossRef]
- Tsai, T.H.; Hsu, Y.C.; Lieu, A.S.; Huang, T.Y.; Kwan, A.L.; Lin, C.L. RTA404, an Activator of Nrf2, Activates the Checkpoint Kinases and Induces Apoptosis through Intrinsic Apoptotic Pathway in Malignant Glioma. J. Clin. Med. 2021, 10, 4805. [Google Scholar] [CrossRef]
- Huang, R.-Z.Z.; Hua, S.-X.X.; Liao, Z.-X.X.; Huang, X.-C.C.; Wang, H.-S.S. Side chain-functionalized aniline-derived ursolic acid derivatives as multidrug resistance reversers that block the nuclear factor-kappa B (NF-κB) pathway and cell proliferation. Medchemcomm 2017, 8, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, P.; Liu, J.; Yang, X.; Xu, S.; Yao, H.; Jiang, J.; Ye, W.; Wu, X.; Xu, J. Synthesis and antitumor activity of novel 3-oxo-23-hydroxybetulinic acid derivatives. Eur. J. Med. Chem. 2014, 87, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, B.; Jabin, I.; Prevost, M.; Ghostin, J.; Becker, J.-P.; Dubois, J.; Kiss, R. 18-beta-Glycyrrhetinic Acid Derivatives with Anti-Tumor Activity. WO Patent 2012/022780 A1, 23 February 2012. [Google Scholar]
- Khusnutdinova, E.; Petrova, A.; Zileeva, Z.; Kuzmina, U.; Zainullina, L.; Vakhitova, Y.; Babkov, D.; Kazakova, O. Novel a-ring chalcone derivatives of oleanolic and ursolic amides with anti-proliferative effect mediated through ros-triggered apoptosis. Int. J. Mol. Sci. 2021, 22, 9796. [Google Scholar] [CrossRef] [PubMed]
- Logashenko, E.B.; Salomatina, O.V.; Markov, A.V.; Korchagina, D.V.; Salakhutdinov, N.F.; Tolstikov, G.A.; Vlassov, V.V.; Zenkova, M.A. Synthesis and Pro-Apoptotic Activity of Novel Glycyrrhetinic Acid Derivatives. ChemBioChem 2011, 12, 784–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markov, A.V.; Sen’kova, A.V.; Zenkova, M.A.; Logashenko, E.B. Novel Glycyrrhetinic Acid Derivative Soloxolone Methyl Inhibits the Inflammatory Response and Tumor Growth in vivo. Mol. Biol. 2018, 52, 262–268. [Google Scholar] [CrossRef]
- Markov, A.V.; Kel, A.E.; Salomatina, O.V.; Salakhutdinov, N.F.; Zenkova, M.A.; Logashenko, E.B. Deep insights into the response of human cervical carcinoma cells to a new cyano enone-bearing triterpenoid soloxolone methyl: A transcriptome analysis. Oncotarget 2019, 10, 5267–5297. [Google Scholar] [CrossRef] [Green Version]
- Alper, P.; Salomatina, O.V.; Salakhutdinov, N.F.; Ulukaya, E.; Ari, F. Soloxolone methyl, as a 18βH-glycyrrhetinic acid derivate, may result in endoplasmic reticulum stress to induce apoptosis in breast cancer cells. Bioorg. Med. Chem. 2021, 30, 115963. [Google Scholar] [CrossRef]
- Markov, A.V.; Odarenko, K.V.; Sen’kova, A.V.; Salomatina, O.V.; Salakhutdinov, N.F.; Zenkova, M.A. Cyano Enone-Bearing Triterpenoid Soloxolone Methyl Inhibits Epithelial-Mesenchymal Transition of Human Lung Adenocarcinoma Cells In Vitro and Metastasis of Murine Melanoma In Vivo. Molecules 2020, 25, 5925. [Google Scholar] [CrossRef]
- Spivak, A.Y.; Gubaidullin, R.R.; Galimshina, Z.R.; Nedopekina, D.A.; Odinokov, V.N. Effective synthesis of novel C(2)-propargyl derivatives of betulinic and ursolic acids and their conjugation with β-d-glucopyranoside azides via click chemistry. Tetrahedron 2016, 72, 1249–1256. [Google Scholar] [CrossRef]
- Moreira, V.M.; Salvador, J.A.R.; Simões, S.; Destro, F.; Gavioli, R. Novel oleanolic vinyl boronates: Synthesis and antitumor activity. Eur. J. Med. Chem. 2013, 63, 46–56. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Gao, Y.; Li, L.; Guo, X.; Liu, D.; Jing, Y.; Zhao, L. Synthesis of methyl 2-cyano-3,12-dioxo-18β-olean-1,9(11)-dien-30-oate analogues to determine the active groups for inhibiting cell growth and inducing apoptosis in leukemia cells. Org. Biomol. Chem. 2014, 12, 6706–6716. [Google Scholar] [CrossRef]
- MORPHEUS. Versatile Matrix Visualization and Analysis Software. Available online: https://software.broadinstitute.org/morpheus/ (accessed on 1 April 2022).
- Liu, H.; Wang, L.; Lv, M.; Pei, R.; Li, P.; Pei, Z.; Wang, Y.; Su, W.; Xie, X.Q. AlzPlatform: An Alzheimer’s disease domain-specific chemogenomics knowledgebase for polypharmacology and target identification research. J. Chem. Inf. Model. 2014, 54, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Laskar, Y.B.; Mazumder, P.B.; Talukdar, A. Das Hibiscus sabdariffa anthocyanins are potential modulators of estrogen receptor alpha activity with favourable toxicology: A computational analysis using molecular docking, ADME/Tox prediction, 2D/3D QSAR and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2021, 2, 1–23. [Google Scholar] [CrossRef]
- Sporn, M.; Liby, K.; Gribble, G.; Honda, T.; Letterio, J. Novel Amide Derivatives of CDDO and Methods of Use Thereof. WO Patent 2008/136838 A1, 13 November 2008. [Google Scholar]
- Pathan, N.; Shende, P. Tailoring of P-glycoprotein for effective transportation of actives across blood-brain-barrier. J. Control. Release 2021, 335, 398–407. [Google Scholar] [CrossRef]
- Dong, J.; Wang, N.N.; Yao, Z.J.; Zhang, L.; Cheng, Y.; Ouyang, D.; Lu, A.P.; Cao, D.S. Admetlab: A platform for systematic ADMET evaluation based on a comprehensively collected ADMET database. J. Cheminform. 2018, 10, 29. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Montanari, F.; Knasmüller, B.; Kohlbacher, S.; Hillisch, C.; Baierová, C.; Grandits, M.; Ecker, G.F. Vienna LiverTox Workspace—A Set of Machine Learning Models for Prediction of Interactions Profiles of Small Molecules with Transporters Relevant for Regulatory Agencies. Front. Chem. 2020, 7, 899. [Google Scholar] [CrossRef]
- Mirzaei, S.; Gholami, M.H.; Hashemi, F.; Zabolian, A.; Farahani, M.V.; Hushmandi, K.; Zarrabi, A.; Goldman, A.; Ashrafizadeh, M.; Orive, G. Advances in understanding the role of P-gp in doxorubicin resistance: Molecular pathways, therapeutic strategies, and prospects. Drug Discov. Today 2022, 27, 436–455. [Google Scholar] [CrossRef]
- Nguyen, T.T.L.; Duong, V.A.; Maeng, H.J. Pharmaceutical Formulations with P-Glycoprotein Inhibitory Effect as Promising Approaches for Enhancing Oral Drug Absorption and Bioavailability. Pharmaceutics 2021, 13, 1103. [Google Scholar] [CrossRef]
- Titova, E.; Shagieva, G.; Ivanova, O.; Domnina, L.; Domninskaya, M.; Strelkova, O.; Khromova, N.; Kopnin, P.; Chernyak, B.; Skulachev, V.; et al. Mitochondria-targeted antioxidant SkQ1 suppresses fibrosarcoma and rhabdomyosarcoma tumour cell growth. Cell Cycle 2018, 17, 1797–1811. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ni, J.; Gao, Y.; Zhang, J.; Liu, X.; Chen, Y.; Chen, Z.; Wu, Y. Integrated proteomics and metabolomics reveals the comprehensive characterization of antitumor mechanism underlying Shikonin on colon cancer patient-derived xenograft model. Sci. Rep. 2020, 10, 14092. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Sun, H.; Yan, H.; Zhang, C.; Zhang, S.; Liu, X.; Hua, N.; Ma, X.; Zheng, J. ZC88, a novel 4-amino piperidine analog, inhibits the growth of neuroblastoma cells through blocking hERG potassium channel. Cancer Biol. Ther. 2013, 14, 450–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US Food and Drug Administration. Guidance for Industry: Bioanalytical Method Validation; US Food and Drug Administration: Silver Spring, MD, USA, 2018.
- European Medicines Agency. Guideline on Bioanalytical Method Validation; European Medicines Agency: Amsterdam, The Netherlands, 2015.
- Zanders, E.D.; Svensson, F.; Bailey, D.S. Therapy for glioblastoma: Is it working? Drug Discov. Today 2019, 24, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, S.M.; Bhardwaj, G.; Goswami, S.; Tonk, R.K.; Goyal, R.K.; Abu-Izneid, T.; Pottoo, F.H. Management of Glioblastoma Multiforme by Phytochemicals: Applications of Nanoparticle-Based Targeted Drug Delivery System. Curr. Drug Targets 2020, 22, 429–442. [Google Scholar] [CrossRef]
- Formolo, C.A.; Williams, R.; Gordish-Dressman, H.; MacDonald, T.J.; Lee, N.H.; Hathout, Y. Secretome Signature of Invasive Glioblastoma Multiforme. J. Proteome Res. 2011, 10, 3149–3159. [Google Scholar] [CrossRef] [Green Version]
- Sabari, J.; Lax, D.; Connors, D.; Brotman, I.; Mindrebo, E.; Butler, C.; Entersz, I.; Jia, D.; Foty, R.A. Fibronectin Matrix Assembly Suppresses Dispersal of Glioblastoma Cells. PLoS ONE 2011, 6, e24810. [Google Scholar] [CrossRef]
- Markov, A.V.; Zenkova, M.A.; Logashenko, E.B. Modulation of Tumour-Related Signaling Pathways by Natural Pentacyclic Triterpenoids and their Semisynthetic Derivatives. Curr. Med. Chem. 2017, 24, 1277–1320. [Google Scholar] [CrossRef]
- Murugan, S.; Amaravadi, R.K. Methods for Studying Autophagy Within the Tumor Microenvironment. Adv. Exp. Med. Biol. 2016, 899, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, J.; Li, K.; Deng, L.; Wang, H. Combination of an Autophagy Inducer and an Autophagy Inhibitor: A Smarter Strategy Emerging in Cancer Therapy. Front. Pharmacol. 2020, 11, 408. [Google Scholar] [CrossRef] [Green Version]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2019, 21, 85–100. [Google Scholar] [CrossRef]
- Yang, N.; Tang, Q.; Qin, W.; Li, Z.; Wang, D.; Zhang, W.; Cao, X.; Lu, Y.; Ge, X.; Sun, H.; et al. Treatment of obesity-related inflammation with a novel synthetic pentacyclic oleanane triterpenoids via modulation of macrophage polarization. EBioMedicine 2019, 45, 473–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibellini, L.; Pinti, M.; Bartolomeo, R.; De Biasi, S.; Cormio, A.; Musicco, C.; Carnevale, G.; Pecorini, S.; Nasi, M.; De Pol, A.; et al. Inhibition of Lon protease by triterpenoids alters mitochondria and is associated to cell death in human cancer cells. Oncotarget 2015, 6, 25466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, E.; Perl, A. Measurement of Mitochondrial Mass by Flow Cytometry during Oxidative Stress. React. Oxyg. Species 2017, 4, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnie, G.; Sotgia, F.; Lisanti, M.P. High mitochondrial mass identifies a sub-population of stem-like cancer cells that are chemo-resistant. Oncotarget 2015, 6, 30472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. ROS-mediated cancer cell killing through dietary phytochemicals. Oxid. Med. Cell. Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Zhang, B.-b.; Wang, D.-g.; Guo, F.-f.; Xuan, C. Mitochondrial membrane potential and reactive oxygen species in cancer stem cells. Fam. Cancer 2015, 14, 19–23. [Google Scholar] [CrossRef]
- Bernstein, S.H.; Venkatesh, S.; Li, M.; Lee, J.; Lu, B.; Hilchey, S.P.; Morse, K.M.; Metcalfe, H.M.; Skalska, J.; Andreeff, M.; et al. The mitochondrial ATP-dependent Lon protease: A novel target in lymphoma death mediated by the synthetic triterpenoid CDDO and its derivatives. Blood 2012, 119, 3321–3329. [Google Scholar] [CrossRef] [Green Version]
- Pinti, M.; Gibellini, L.; Nasi, M.; De Biasi, S.; Bortolotti, C.A.; Iannone, A.; Cossarizza, A. Emerging role of Lon protease as a master regulator of mitochondrial functions. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; Losi, L.; De Biasi, S.; Nasi, M.; Tartaro, D.L.; Pecorini, S.; Patergnani, S.; Pinton, P.; De Gaetano, A.; Carnevale, G.; et al. LonP1 differently modulates mitochondrial function and bioenergetics of primary versus metastatic colon cancer cells. Front. Oncol. 2018, 8, 254. [Google Scholar] [CrossRef]
- Bota, D.A.; Ngo, J.K.; Davies, K.J.A. Downregulation of the human Lon protease impairs mitochondrial structure and function and causes cell death. Free Radic. Biol. Med. 2005, 38, 665–677. [Google Scholar] [CrossRef] [Green Version]
- Kalvala, A.K.; Yerra, V.G.; Kumar, A. LONP1 induction by SRT1720 attenuates mitochondrial dysfunction against high glucose induced neurotoxicity in PC12 cells. Toxicol. In Vitr. 2020, 62, 104695. [Google Scholar] [CrossRef] [PubMed]
- Gibellini, L.; Pinti, M.; Boraldi, F.; Giorgio, V.; Bernardi, P.; Bartolomeo, R.; Nasi, M.; De Biasi, S.; Missiroli, S.; Carnevale, G.; et al. Silencing of mitochondrial Lon protease deeply impairs mitochondrial proteome and function in colon cancer cells. FASEB J. 2014, 28, 5122–5135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Nafría, J.; Ondrovičová, G.; Blagova, E.; Levdikov, V.M.; Bauer, J.A.; Suzuki, C.K.; Kutejová, E.; Wilkinson, A.J.; Wilson, K.S. Structure of the catalytic domain of the human mitochondrial Lon protease: Proposed relation of oligomer formation and activity. Protein Sci. 2010, 19, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wlodawer, A.; Sekula, B.; Gustchina, A.; Rotanova, T.V. Structure and the Mode of Activity of Lon Proteases from Diverse Organisms. J. Mol. Biol. 2022, 434, 167504. [Google Scholar] [CrossRef]
- Brix, N.; Samaga, D.; Belka, C.; Zitzelsberger, H.; Lauber, K. Analysis of clonogenic growth in vitro. Nat. Protoc. 2021, 16, 4963–4991. [Google Scholar] [CrossRef]
- Welch, D.R.; Hurst, D.R. Defining the Hallmarks of Metastasis. Cancer Res. 2019, 79, 3011–3027. [Google Scholar] [CrossRef]
- Cai, H.; Liu, W.; Liu, X.; Li, Z.; Feng, T.; Xue, Y.; Liu, Y. Advances and Prospects of Vasculogenic Mimicry in Glioma: A Potential New Therapeutic Target? Onco. Targets. Ther. 2020, 13, 4473. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, X.; Zhao, P.; Zhao, H.; Gao, W.; Wang, L. Celastrol Suppresses Glioma Vasculogenic Mimicry Formation and Angiogenesis by Blocking the PI3K/Akt/mTOR Signaling Pathway. Front. Pharmacol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Mitra, D.; Ray, S.; Biswas, N.; Banerjee, S.; Majumder, B.; Mustafi, S.M.; Murmu, N. Reversing effect of Lupeol on vasculogenic mimicry in murine melanoma progression. Microvasc. Res. 2019, 121, 52–62. [Google Scholar] [CrossRef]
- Viennois, E.; Merlin, D.; Gewirtz, A.T.; Chassaing, B. Dietary emulsifier-induced low-grade inflammation promotes colon carcinogenesis. Cancer Res. 2017, 77, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Yassin, M.A.; Leknes, K.N.; Sun, Y.; Lie, S.A.; Finne-Wistrand, A.; Mustafa, K. Surfactant tuning of hydrophilicity of porous degradable copolymer scaffolds promotes cellular proliferation and enhances bone formation. J. Biomed. Mater. Res. Part A 2016, 104, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Caddeo, C.; Manca, M.L.; Peris, J.E.; Usach, I.; Diez-Sales, O.; Matos, M.; Fernàndez-Busquets, X.; Fadda, A.M.; Manconi, M. Tocopherol-loaded transfersomes: In vitro antioxidant activity and efficacy in skin regeneration. Int. J. Pharm. 2018, 551, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.; Li, Q.; Zhang, X.; Chen, Y.; Yu, K. mTOR Promotes Tissue Factor Expression and Activity in EGFR-Mutant Cancer. Front. Oncol. 2020, 10, 1615. [Google Scholar] [CrossRef]
- Ye, Z.; Ai, X.; Zhao, L.; Fei, F.; Wang, P.; Zhou, S. Phenotypic plasticity of myeloid cells in glioblastoma development, progression, and therapeutics. Oncogene 2021, 40, 6059–6070. [Google Scholar] [CrossRef] [PubMed]
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular Matrix in the Tumor Microenvironment and Its Impact on Cancer Therapy. Front. Mol. Biosci. 2019, 6, 160. [Google Scholar] [CrossRef] [Green Version]
- Mohiuddin, E.; Wakimoto, H. Extracellular matrix in glioblastoma: Opportunities for emerging therapeutic approaches. Am. J. Cancer Res. 2021, 11, 3742. [Google Scholar]
- Berg, T.J.; Pietras, A. Radiotherapy-induced remodeling of the tumor microenvironment by stromal cells. Semin. Cancer Biol. 2022; in press. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, W.; Wang, X.; Liu, Y.; Li, X. Sequential Drug Release to Modulate Collagen Synthesis and Promote Micelle Penetration in Tumors. ACS Biomater. Sci. Eng. 2019, 5, 1343–1353. [Google Scholar] [CrossRef]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Annese, T.; Ruggieri, S.; Tamma, R.; Crivellato, E. Limitations of Anti-Angiogenic Treatment of Tumors. Transl. Oncol. 2019, 12, 981–986. [Google Scholar] [CrossRef]
- Park, J.S.; Park, I.; Koh, G.Y. Benefits and Pitfalls of Tumor Vessel Normalization. In Tumor Angiogenes: A Key Target Cancer Therapy; Springer: Cham, Switzerland, 2019; pp. 51–71. [Google Scholar] [CrossRef]
- Wechman, S.L.; Emdad, L.; Sarkar, D.; Das, S.K.; Fisher, P.B. Vascular mimicry: Triggers, molecular interactions and in vivo models. Adv. Cancer Res. 2020, 148, 27–67. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, Y.; Shi, X.; Hao, S.; Zhang, F.; Guo, Z.; Gao, Y.; Guo, H.; Liu, L. Oridonin inhibits tumor angiogenesis and induces vessel normalization in experimental colon cancer. J. Cancer 2021, 12, 3257. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Sawant, M.A.; Lavhale, M.S.; Krishnapati, L.S.; Ghaskadbi, S.; Sitasawad, S.L. AECHL-1, a novel triterpenoid, targets tumor neo-vasculature and impairs the endothelial cell cytoskeleton. Angiogenesis 2015, 18, 283–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. Software news and update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, C.; Bagga, M.; Kaur, A.; Westermarck, J.; Abankwa, D. ColonyArea: An ImageJ plugin to automatically quantify colony formation in clonogenic assays. PLoS ONE 2014, 9, e92444. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | IC50, µM | |||
|---|---|---|---|---|
| Cervix | Duodenum | Melanoma | Fibroblasts | |
| HeLa | HuTu-80 | B16 | hFF3 | |
| 1 | 9.9 ± 0.7 | 2.5 ± 0.3 | 4.2 ± 0.3 | 16.0 ± 1.9 |
| 2 | 1.5 ± 0.1 | 1.4 ± 0.1 | 3.3 ± 0.2 | 1.2 ± 0.1 |
| 3 | 3.5 ± 0.2 | 1.7 ± 0.2 | 1.4 ± 0.1 | 6.8 ± 1.3 |
| 4 | 2.3 ± 0.4 | 1.3 ± 0.1 | 1.5 ± 0.1 | 3.6 ± 0.3 |
| 5 | 1.5 ± 0.2 | 2.3 ± 0.2 | 1.6 ± 0.1 | 12.9 ± 0.8 |
| 6 | 2.6 ± 0.5 | 2.1 ± 0.5 | 3.3 ± 0.5 | 4.1 ± 0.3 |
| 7 | 4.9 ± 0.8 | 2.0 ± 0.7 | 3.3 ± 0.8 | 7.7 ± 1.1 |
| 8 | 1.5 ± 0.1 | 1.4 ± 0.1 | 1.9 ± 0.1 | 1.8 ± 0.1 |
| 9 | 1.6 ± 0.1 | 1.5 ± 0.1 | 2.1 ± 0.3 | 3.6 ± 0.3 |
| 10 | 1.8 ± 0.1 | 1.5 ± 0.2 | 1.7 ± 0.1 | 1.6 ± 0.1 |
| 11 | 2.5 ± 0.1 | 1.5 ± 0.1 | 1.4 ± 0.1 | 1.3 ± 0.2 |
| 12 | 1.5 ± 0.1 | 1.5 ± 0.1 | 1.5 ± 0.2 | 1.4 ± 0.2 |
| SM | 1.3 ± 0.1 | 1.5 ± 0.1 | 1.6 ± 0.1 | 4.0 ± 0.4 |
| ID | IC50, µM | |||
|---|---|---|---|---|
| Glioblastoma | Neuroblastoma | |||
| U87 | U118 | KELLY | Neuro2a | |
| 4 | 2.0 ± 0.1 | 3.1 ± 0.5 | 1.5 ± 0.1 | 1.9 ± 0.2 |
| 5 | 5.5 ± 1.1 | 7.0 ± 0.7 | 1.6 ± 0.1 | 2.7 ± 0.3 |
| 6 | 4.6 ± 0.4 | 3.8 ± 0.5 | 1.6 ± 0.1 | 2.8 ± 0.4 |
| 7 | 5.3 ± 0.7 | 5.2 ± 1.3 | 1.8 ± 0.3 | 3.7 ± 0.6 |
| 8 | 1.7 ± 0.1 | 2.0 ± 0.1 | 1.4 ± 0.1 | 1.6 ± 0.1 |
| 9 | 2.0 ± 0.2 | 1.9 ± 0.2 | 1.4 ± 0.1 | 1.6 ± 0.1 |
| 10 | 2.5 ± 0.2 | 2.1 ± 0.2 | 1.5 ± 0.1 | 1.4 ± 0.1 |
| 11 | 1.5 ± 0.1 | 1.9 ± 0.1 | 1.8 ± 0.2 | 1.4 ± 0.1 |
| 12 | 1.9 ± 0.1 | 1.9 ± 0.2 | 1.8 ± 0.1 | 1.6 ± 0.1 |
| SM | 1.8 ± 0.1 | 1.9 ± 0.1 | 1.6 ± 0.1 | 1.4 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markov, A.V.; Ilyina, A.A.; Salomatina, O.V.; Sen’kova, A.V.; Okhina, A.A.; Rogachev, A.D.; Salakhutdinov, N.F.; Zenkova, M.A. Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo. Pharmaceuticals 2022, 15, 603. https://doi.org/10.3390/ph15050603
Markov AV, Ilyina AA, Salomatina OV, Sen’kova AV, Okhina AA, Rogachev AD, Salakhutdinov NF, Zenkova MA. Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo. Pharmaceuticals. 2022; 15(5):603. https://doi.org/10.3390/ph15050603
Chicago/Turabian StyleMarkov, Andrey V., Anna A. Ilyina, Oksana V. Salomatina, Aleksandra V. Sen’kova, Alina A. Okhina, Artem D. Rogachev, Nariman F. Salakhutdinov, and Marina A. Zenkova. 2022. "Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo" Pharmaceuticals 15, no. 5: 603. https://doi.org/10.3390/ph15050603
APA StyleMarkov, A. V., Ilyina, A. A., Salomatina, O. V., Sen’kova, A. V., Okhina, A. A., Rogachev, A. D., Salakhutdinov, N. F., & Zenkova, M. A. (2022). Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo. Pharmaceuticals, 15(5), 603. https://doi.org/10.3390/ph15050603